Подсемейство Sedoideae (Crassulaceae) флоры Сибири и российского Дальнего Востока (систематика, биоморфология, филогения)
С. Б. Гончарова
Глава 7. Биоморфология представителей подсемейства Sedoideae
7.1. Понятие «жизненная форма», подходы и предпосылки классификации
Термин «жизненная форма» (Lebensform) был предложен E. Warming в 1884 году, по его определению, это – «форма, в которой вегетативное тело растения находится в гармонии с внешней средой в течение всей его жизни от семени до отмирания» (цит. по: Серебрякова, 1980).
«Жизненная форма» (ЖФ) – многозначная категория, включающая в себя понятие, класификационную единицу и конкретный организм. Как понятие «жизненная форма» – фундаментальная категория в биоморфологии. Как классификационная единица – наименьшая, неделимая конвергентная таксономическая единица эколого-морфологической (биоморфологической) иерархической классификации. В отличие от «вида» «жизненная форма» отражает фенотипическое сходство, а не генетическое родство (Хохряков, 1981).
Каждый организм характеризуется уникальной, свойственной только ему ЖФ (биоморфой), это – «совокупность некоторого числа признаков, определяющих внешний облик (габитус) растения и отражающих его приспособления к условиям местообитания» (Серебрякова, 1980, с. 76), однако если организм всегда принадлежит к единственно возможному виду, то вид может характеризоваться несколькими жизненными формами в зависимости от условий произрастания. Вариации жизненных форм в пределах вида могут послужить источником как дивергентной эволюции видов, так и конвергентной эволюции жизненных форм (Bocher, цит. по: Серебрякова, 1980).
Существуют два основных подхода к определению понятия «жизненная форма»: 1) классификационный, принимающий, что растение характеризуется биоморфой, присущей взрослому генеративному растению (Серебряков, 1962, 1964; Серебрякова, 1980; Шорина, 2000), и 2) индивидуалистический, принимающий, что растение в течение жизни проходит различные этапы, являющиеся отдельными биоморфами, названными «фазами морфогенеза» или «онтобиоморфами» (Серебрякова, 1980; Хохряков, 1981; Мазуренко, 1986; Шорина, 2000 б).
Мы, вслед за И.Г. Серебряковым (1964, с. 146), жизненной формой считаем «габитус определенной группы растений (включая подземные и надземные органы), возникающий в онтогенезе в результате роста и развития в определенных условиях среды. Габитус исторически возникает в данных почвенно-климатических условиях как выражение приспособленности растений к этим условиям».
Габитус определяется «конструкцией» организма (Хохряков, 1994), сочетанием закономерно повторяющихся единиц (метамеров или модулей разного порядка). Структурные единицы, формирующие жизненные формы – побеги и побеговые системы. Наиболее важные из них – одноосный (моноподиальный) побег [универсальный модуль]; комплекс сохраняющегося одноосного побега и комплекс побеговой оси одного видимого порядка [основные модули] (Шафранова, 1980, 2001; Шафранова, Гатцук, 1994; Гатцук, 1994; Савиных, 2000).
Для корневищных трав основной модуль – это монокарпический побег, парциальный куст или система парциальных кустов, для полудревесных – комплекс побеговой оси одного видимого порядка.
Мы, вслед за В.А. Недолужко (1997), считаем, что при классифицировании жизненных форм необходимо использовать преимущественно признаки вегетативной сферы растения, а именно: тип универсального и основного модулей, особенности их сочетания (архитектурные модели), характер нарастания и ветвления системы побегов (модели побегообразования), особенности подземных органов, отражающие приспособления растений к современным или историческим условиям среды обитания.
Из всего разнообразия систем жизненных форм, существующих в современной биоморфологии, нами использованы классификации И.Г. Серебрякова (1964) и С. Raunkiaer (1934, 1937). Их сочетание, по нашему мнению, позволяет наиболее полно охарактеризовать растение. Система И.Г. Серебрякова отражает интегральную реакцию организма на среду обитания, она характеризуется не только физиономическими признаками, но и рядом биологических свойств: длительностью жизни универсального, основного модулей и растения в целом, ритмом развития, способом питания, способом вегетативного размножения и т.д. Эта система иерархична и преследует цель показать общее направление эволюции жизненных форм.
Главные признаки, положенные в основу классификации К. Raunkiaer (1934), – положение почек возобновления относительно поверхности почвы и способ их защиты от воздействия неблагоприятных условий среды. Однако, как заметил Ж. Манжено (цит. по: Серебрякова, 1972), эти признаки настолько существенны, что с ними оказался прочно коррелятивно связанным целый комплекс других очень важных признаков, в том числе и физиономических.
Наибольшие затруднения при классифицировании биоморф растений сезонного климата вызывает разделение на «древесные» и «травянистые», так как, несмотря на длительную историю существования науки о жизненных формах, биоморфологи не пришли к единому мнению о понятии «травянистого растения». В связи с этим за основу мы взяли определение Л.Е. Гатцук (1976): травянистые многолетники: 1) растения с удлиненными и полурозеточными побегами, надземная ортотропная часть осей которых живет только один вегетационный период; 2) растения, у которых могут сохраняться многолетние ползучие или плагиотропные части, плотно прижатые к земле или расположенные под поверхностью и укореняющиеся; 3) растения с розеточными побегами, оси которых тесно прижаты к субстрату. Бесспорно, что к «травянистым» должны быть отнесены гемикриптофиты, криптофиты и терофиты. Однако определить степень «плотности» прижатия к субстрату не всегда бывает просто, особенно в условиях скальных или каменистых местообитаний, и в этом случае различить «полудревесные» и «травянистые» биоморфы как по физиономическим, так и по анатомическим признакам не всегда легко. Критерий, предложенный А.П. Хохряковым (1976), – существование у трав цикличности в нарастании, отмирании и втягивающей деятельности осевых (побеговых и корневых метамеров) – также далеко не всегда применим к растениям, обитающим на каменистых грунтах.
Процессы возникновения и преобразования, а также взаимосвязи различных биоморф могут изучаться как в масштабах ценозов и флор, так и в узких таксономических группах. В первом случае спектр биоморф в пределах ценоза или флоры отражает реакцию различных организмов на влияние сходных условий окружающей среды и позволяет выявить общие тенденции приспособления растений к ним. Во втором – отражает возможные преобразования групп близких организмов под влиянием различающихся условий.
Признаки вегетативной сферы, жизненных форм и форм роста всегда имели большое значение в систематике семейства. Так, S Schönland (1891) в обработке толстянковых для первого издания «Die Natürlichen Pflanzenfamilien» детально описал развитие хамефитных Sedum. Т. Nakai (1938), как отмечено выше, изучая японские виды Rhodiola, впервые указал на аксилярную природу генеративных побегов и описал чешуевидные корневищные листья. G. Hegi (1921-1923) в обработке семейства Crassulaceae флоры Центральной Европы разделил Sedum s. l. на три группы, соответственно способу перезимовки: хамефиты, гемикриптофиты и терофиты. Данным автором описан ход онтогенеза многолетних очитков. Он указал, что некоторые виды формируют плотные куртины, или «маты», а также выделил два типа побегов: 1) стерильные плагиотропные и укореняющиеся, многолетние и 2) цветоносные ортотропные, однолетние. Н. Ohba (1977, 1978) в обработке Sedoideae Восточной Азии в качестве одного из важнейших признаков, характеризующих выделенный им род Hylotelephium, привел описание однолетних цветоносных побегов, развивающихся из перезимовавших пазушных почек на прошлогодних участках побегов или из придаточных почек на корневищах. В классификации европейских, северо-африканских и западно-азиатских Sedoideae V. Grulich (1984) указал, что большинство представителей подсемейства хамефиты, однако роды Hylotelephium и Aizopsis включают в себя главным образом гемикриптофиты с эпи-гипогеогенными корневищами, а криптофитные жизненные формы характерны для примитивных групп, с плоскими листьями (Rhodiola, Hylotelephium, Aizopsis).
Специальные работы, посвященные биоморфологии толстянковых, немногочисленны, они охватывают, как правило, небольшие таксономические или географические группы. Так, G. Knaben (1966), изучая малолетние очитки Скандинавии, выделил одно- и двулетние типы. Автор высказал предположение, что двулетники являются переходными между гемикриптофитами и терофитами, а также проанализировал ход онтогенеза выделенных им типов. R.T. Clausen (1977) подробно изучен морфогенез мексиканских двулетних видов. Одна из наиболее полных классификаций жизненных форм Sedum s.l. (включая Rhodiola, Hylotelephium, Aizopsis, Orostachys, Pseudosedum и др.), построенная на основе анализа биоморфологических признаков 70 видов рода, представлена W. Böttcher и E. Jäger (1984). Авторами рассмотрено распространение биоморф в соответствии с климатическими условиями, выявлено 10 типов четко различающихся жизненных форм, коррелирующих с ритмами климата:
- 1) вечнозеленые кустарники и карликовые деревья с выраженно суккулентными «круглыми» (вальковатыми) или плоскими листьями, Pachyphyllum-тип;
- 2) вечнозеленые, образующие дерновинки хамефиты с ползучими безрозеточными побегами и «круглыми» листьями, Rupestre-тип;
- 3) вечнозеленые с розеточными побегами, Atlanticum-тип;
- 4) вечнозеленые или почти вечнозеленые, образующие дерновинки плосколистные хамефиты, Spurium-тип;
- 5) летнезеленые симподиально возобновляющиеся травы, Telephium-тип;
- 6) летнезеленые карликовые кустарники, Populifolium-тип;
- 7) летнезеленые, моноподиально возобновляющиеся травы, Rhodiola-тип;
- 8) зимнезеленые «круглолистные» хамефиты, Amplexicaule-тип;
- 9) плосколистные терофиты, Drymarioides-тип;
- 10) «круглолистные» (вальковатолистные) терофиты (малолетние), Annuum-тип.
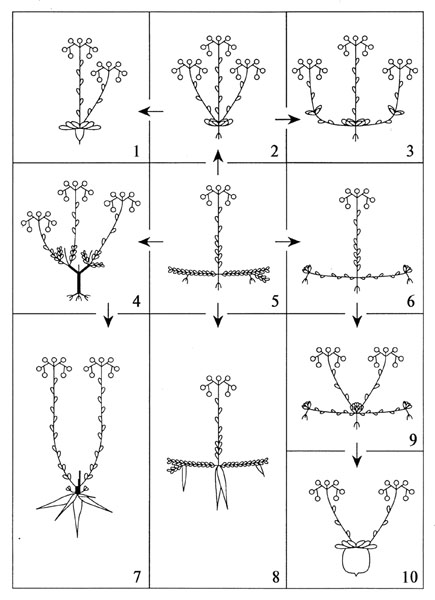
Рис. 35. Жизненные формы и формы роста евро-азиатских Sedoideae (Hart, 1994).
1-3 – однолетники и гапаксантные формы; 5 – малоспециализированные многолетние формы; 6, 9-10 – розеточные, столонные и моноподиальные формы; 4, 7 – кустарниковые, древесные и корневищные формы; 7,8 – клубнекорневые формы.
|
Еще более подробная и обстоятельная работа была выполнена H. ‘t Hart (1994). На основании изучения особенностей морфологии, анатомии и особенностей роста и развития 74 европейских Sedoideae названным автором были выделены 10 комбинаций жизненных форм и форм роста (рис. 28). H. ‘t Hart отмечает, что представленные жизненные формы в целом соответствуют таковым, предложенным W. Böttcher и E. Jäger. Основное отличие классификации H. ‘t Hart в использовании анатомических признаков.
Все известные нам классификации жизненных форм отдельных родов и семейства Crassulaceae в целом, разработанные российскими авторами (Безделева, 1993; Гончарова, 1996б, 1997, 2000а; Бялт, 1999а, 2004б; Адонина, 2002), основаны на системе И.Г. Серебрякова (1962, 1964). Классификации отражают различные аспекты формирования и функционирования биоморф (структурно-морфологические, фитоценотические и др.) и, в целом, соответствуют друг другу.
Обзор биоморф дальневосточных Sedum s.l. (включая Hylotelephium и Aizopsis) для целей систематики дан Т.А. Безделевой (1993). На основании анализа строения подземных и надземных органов 14 видов автором сделан вывод о большом значении признаков биоморф для дифференциации видов рода и выделены 5 типов жизненных форм: 1) стержнекорневые травянистые поликарпики, с моноциклическими удлиненными монокарпическими побегами; 2) корневищно-стержнекорневые травянистые поликарпики, с удлиненными моноциклическими монокарпическими побегами; 3) корневищно-стержнекорневые травянистые поликарпики, с полурозеточными моноциклическими монокарпическими побегами; 4) короткокорневищные травянистые поликарпики, с удлиненными моноциклическими монокарпическими побегами и мочкой в разной степени утолщенных придаточных корней; 5) вегетативно размножающиеся короткокорневищные травянистые поликарпики, с удлиненными монокарпическими побегами.
Обзор литературных источников, посвященных биоморфам толстянковых, показал, что отсутствует единообразие в классифицировании жизненных форм, это значительно затрудняет сравнение результатов и поиск закономерностей биоморфологической эволюции семейства. Но достичь его довольно трудно, так как значительное разнообразие подходов к классификации приводит к разнообразию названий биоморф. |
7.2. Классификация жизненных форм Sedoideae Сибири и российского Дальнего Востока
Суммируя данные, приведенные в гл. 4, можно сказать, что изученные виды имеют существенные различия в продолжительности жизненных циклов, периодичности плодоношения, способах возобновления. Монокарпические побеги изученных Sedoideae и их системы характеризуются различными сочетаниями следующих признаков:
- 1) побеги могут быть удлиненными или полурозеточными;
- 2) побег, как правило, моноциклический, редко ди- или полициклический;
- 3) надземные части побега отмирают полностью или частично;
- 4) корневища эпигеогенные (обычно наземные) либо гипогеогенные (подземные);
- 5) возобновление системы побегов моноподиальное, симподиальное или смешанное;
- 6) ветвление системы побегов мезосимподиальное или базисимподиальное;
- 7) подземные органы представлены корневищами, каудексами, запасающими корнями, кистевой или редко стержневой корневой системой;
- 8) почки возобновления расположены на многолетних частях побегов (резидах), они могут быть погружены или полупогружены в почву, располагаться выше или на уровне субстрата.
Основываясь на собственных исследованиях (см. также гл. 4), анализе литературных данных и используя обозначенные выше подходы, мы предлагаем следующую классификацию биоморф Sedoideae флоры Сибири и российского Дальнего Востока.
Таблица 7
Классификация жизненных форм Sedoideae флоры Сибири и Российского Дальнего Востока
отдел Древесные растения
тип Кустарники и кустарнички
класс Кустарнички с ассимилирующими побегами суккулентного типа (хамефиты)
подкласс Суккулентнолистовые кустарнички Hylotelephium populifolium
отдел Полудревесные растения
тип Полукустарники и полукустарнички
класс Полукустарнички с ассимилирующими побегами суккулентного типа (хамефиты)
подкласс Суккулентнолистовые полукустарнички Aizopsis middendorfiana ssp. middendorfianа, A. middendorfianа ssp. arcuatа, A. middendorfianа ssp. sichotensis, A. kurilensis
отдел Наземные травянистые растения
тип Травянистые поликарпики
класс Травянистые поликарпики с ассимилирующими побегами суккулентного типа
подкласс Суккулентнолистовые травянистые облигатные поликарпики
группа Полурозеточные хамефиты
подгруппа Образующие дерновинки (латки) Orostachys aggregata, O. furusei, O. paradoxa
группа Длиннопобеговые хамефиты Hylotelephium cyaneum, H. pluricaule, H. ewersii, Aizopsis hybrida, Sedum acre
группа Длиннопобеговые гемикриптофиты
подгруппа Каудексообразующие Aizopsis selskiana, Rhodiola borealis, R. rosea ssp. rosea, R. pinnatifida, R. pinnatifida ssp. subpinnata, R. rosea ssp. sachalinensis, R. rosea ssp. krivochizhinii
подгруппа Подушковидные гемикриптофиты Rhodiola coccinea, R. quadrifida
подгруппа Корневищно-стержнекорневые гемикриптофиты
вариант Моноподиально корневищно-стержнекорневые Rhodiola algida, R. angusta, R. integrifolia, R. ishidae, R. stephanii
группа Длиннопобеговые криптофиты
подгруппа Короткокорневищные длиннопобеговые криптофиты
вариант Симподиально корневищные Aizopsis aizoon ssp. aizoon, A. aizoon ssp. baicalensis A. kamtschatica, A. litoralis, A. × pilosa, Hylotelephium erythrostictum, H. pallescens, H. ussuriense, H. verticillatum, H. viviparum, Pseudosedum lievenii, S. albertii
вариант Корневищно-клубнекорневые криптофиты Hylotelephium pseudospectabile, H. telephium ssp. stepposum, H. triphyllum ssp. triphyllum, H. triphyllum ssp. sukaczevii
подкласс Суккулентнолистовые травянистые факультативные поликарпики (олигокарпики)
группа Полурозеточные гемикриптофиты Orostachys fimbriata, O. gorovoii, O. japonica, O. malacophylla, O. spinosa, O. thyrsiflora
тип Травянистые монокарпики
класс Травянистые монокарпики с ассимилирующими побегами суккулентного типа
группа Полурозеточные гемикриптофиты-терофиты Orostachys maximowiczii
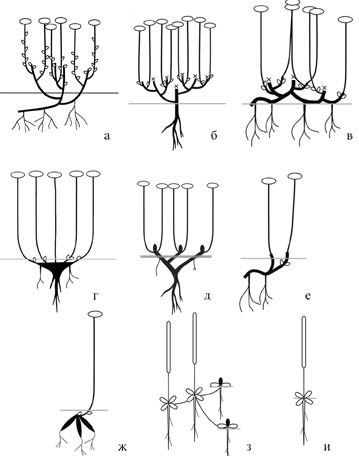
Рис. 36. Жизненные формы Sedoideae Сибири и российского Дальнего Востока.
а – кустарнички; б – полукустарнички; в – длиннопобеговые суккулентнолистовые хамефиты; г – каудексообразующие гемикриптофиты; д – моноподиально корневищно-стержнекорневые гемикриптофиты; е – симподиально корневищные криптофиты; ж – корневищно-клубнекорневые криптофиты; з – образующие дерновинки полурозеточные хамефиты; и – полурозеточные гемикриптофиты.
|
Как видно из табл. 7 (см. также рис. 36), большинство Sedoideae изучаемого региона – суккулентнолистовые поликарпические травы. Подобные биоморфы обычны и для семейства в целом, около 70 % видов Crassulaceae относятся к отделу «Наземные травы» (Бялт, 2004б). Наиболее обычны корневищные криптофиты, менее представлены – гемикриптофиты. Для видов этих групп характерны отсутствие системы главного корня и наличие четко выраженного ризома, состоящего из значительного числа годичных приростов.
Как отмечено выше, корневища представителей этих групп различаются типом нарастания, в соответствии с ним мы разделяем биоморфы с симподиально (рис. 36, в, е) и моноподиально (рис. 36, д) нарастающими корневищами. Хотя многолетние органы представителей обеих групп близки морфологически, но они имеют существенные анатомические особенности (см. также гл. 4) и, вероятно, различное происхождение.
И.Г. Серебряков относит к отделу «наземных травянистых растений» растения, которые «совершенно лишены надземных одревесневших многолетних осей; все надземные стебли травянистые и покровные ткани их первичные, без перидермы» (1964, с. 170). Однако существует «серьезный конфликт» между биоморфологическими и анатомическими критериями, так как «одревеснение» и перидерма часто присутствуют и в однолетних, типично «травянистых» побегах (например, у Aizopsis aizoon и Hylotelephium triphyllum, см. также гл. 4).
Широко принимается, что многолетние травянистые растения сезонного климата – гемикриптофиты и криптофиты, однако изучение разнообразия жизненных форм в различных таксонах показало и существование здесь травянистых хамефитов (Гончарова, 1997; Савиных, 2000). Хамефиты, по определению С. Raunkiaer (1937, с. 34), – «plants whose buds or shoot-apices … are situated on shoot or portion of shoots which either lie on surface of the earth or are situated quite near to it, so that … snow can protect them, or … the buds be partially protected by the withered remains of the plants on the surface of the earth», то есть растения, почки или апексы побегов которых либо лежат на поверхности почвы, либо расположены близко к ней и могут бытьзакрыты снегом или опадом. Необходимо отметить, что в зависимости от условий произрастания (на мелкоземистых рыхлых почвах), такие почки могут частично погружаться в субстрат, демонстрируя этим возможность перехода к гемикриптофитам. |
Способность образовывать дерновинки (куртины, латки) часто упоминается при описаниях биоморф толстянковых (Knaben, 1966; Горбачевская и др., 2000). При этом формируется особый тип жизненных форм «травянистых хамефитов», характеризующийся наличием одно-, редко двулетних вегетативно-генеративных побегов и малолетних наземных побегов, своеобразных «наземных корневищ», слагающихся из некоторого числа годичных приростов (рис. 36, в). Подобно типичному «подземному» корневищу «наземное» имеет хорошо выраженную метамерную структуру, оно нарастает у верхушки и отмирает у основания, но, в отличие от «подземного», его способность к ветвлению и вегетативному размножению хорошо развита, а анатомические особенности [сильная паренхиматизация стелы, отсутствие типичных годичных колец, вторичное утолщение, преимущественно, за счет образования мощной коровой паренхимы и корки, пучковое строение стелы у O. paradoxa (см. рис. 21, гл. 4)] указывают на «травянистость» такого корневища.
Как видно из табл. 7, Hylotelephium pseudospectabile, H. telephium ssp. stepposum, H. triphyllum ssp. triphyllum, H. triphyllum ssp. sukaczevii и классифицированы нами как корневищно-корнеклубневые криптофиты, они характеризуются наличием специализированных запасающих корней, развивающихся на очень коротких корневищах. Эти биоморфы очень редко встречаются среди очитковых, не более 10 из них имеют подобные жизненные формы, но как отмечено выше, все представители Hylotelephium sect. Hylotelephium имеют в большей или меньшей степени утолщенные корни.
Единственный вид – H. populifolium отнесен нами к классу кустарничков с ассимилирующими побегами суккулентного типа. Это, вероятно, реликт третичной флоры (Пяк, 2003), сохранивший жизненную форму, характерную для представителей толстянковых более теплых климатов.
Для «полудревесных» биоморф характерно то, что удлиненные ортотропные надземные оси на значительной части их длины остаются травянистыми (или слабо одревесневают) и отмирают ежегодно, сохраняются и одревесневают только базальные части надземных осей (Серебряков, 1964). «Полудревесные» растения также довольно редки среди толстянковых, произрастающих в условиях умеренного климата, полукустарнички обычны среди субтропических и тропических Crassulaceae, но сезонные и бессезонные полукустарнички существенно отличаются как морфологически, так и анатомически (Böttcher, Jäger, 1984; Hart, 1994).
Довольно большую группу составляют каудексообразующие и моноподиально корневищно-стержнекорневые гемикриптофиты, представленные главным образом в роде Rhodiola. Биоморфу моноподиально корневищно-стержнекорневых растений можно также считать вариантом каудексообразующей жизненной формы, так как главный корень также функционирует значительную часть жизни особи. Главное отличие этих биоморф заключается в длине веточек каудекса (каудикул), у первых каудикулы короткие, у вторых – длинные (иногда несколько десятков). Корневищно-стержнекорневые биоморфы очитковых, вероятно, можно считать переходными между каудексообразующими и корневищными биоморфами, так как в начале жизненного цикла (часто многие десятки лет) они развиваются как каудексообразующие, а в конце (также весьма длительный период времени) – как корневищные (см. также гл. 7.3).
Два вида (Rhodiola coccinea и R. quadrifida) отнесены нами к подгруппе подушковидных растений. Это – высокоспециализированные биоморфы, адаптированные к жизни в экстремальных экологических условиях (Волков, 2003). И.В. Волков (2003) классифицирует биоморфы, характерные названным видам, как – розеточные подушки и отмечает, что в отличие от других типов подушек, розеточные не сохраняются как единое целое, так как отдельные побеги отмирают по отцветании, они заменяются укореняющимися побегами возобновления. Прошлогодние отмершие побеги Rhodiola coccinea и R. quadrifida не опадают (как это типично для большинства других очитковых), а многие годы сохраняются на растении. Между этими побегами скапливаются листовой опад и мелкозем, обеспечивая защиту почкам возобновления и возможность более активного образования придаточных корней. Характерной особенностью подушковидных растений является также развитие мощного стержневого корня, выполняющего «якорную» функцию. С. Raunkiaer (1934) относил подушковидные растения к хамефитам, однако, хотя их почки возобновления располагаются значительно выше уровня почвы, но они погружены в субстрат, скапливающийся внутри подушки, а следовательно, являются гемикриптофитами.
Как уже было отмечено нами выше, некоторые представители рода Orostachys могут вести себя либо как монокарпики, либо как поликарпики. И возможно, что большая или меньшая степень поликарпичности является реакцией на условия обитания. В соответствии с этим мы классифицируем их как «травянистые факультативные поликарпики с ассимилирующими побегами (дициклическими монокарпическими) суккулентного типа». Кроме того, у некоторых представителей варьирует и продолжительность жизни монокарпического побега, они могут быть либо моно-, либо дициклическими (O. maximowiczii).
Жизненные формы большинства изученных представителей Sedoideae из естественных местообитаний весьма стабильны (термины по: Жмылев, 2000), однако существует и ряд лабильных видов. Так, Aizopsis litoralis в условиях скальных и щебнистых местообитаний является короткокорневищным растением, а на прибрежных песках, при погребении и укоренении плагиотропной части побега, у него образуется разветвленное корневище, длина годичного прироста которого может достигать 12-15 см, то есть возникает длиннокорневищная биоморфа. Еще более разнообразны варианты жизненных форм A. kamtschatica, в зависимости от условий обитания он может формировать полукустарничковые, короткокорневищные или переходные между двумя типами жизненные формы.
Значительные вариации биоморф наблюдаются в условиях культуры на улучшенном агрофоне. Например, у Aizopsis selskiana при благоприятных условиях перезимовки подобно хамефитам образуются почки возобновления гораздо выше уровня почвы. Наблюдается общая тенденция своеобразной «хамефитизации» гемикриптофитов и даже криптофитов.
Рассматривая разнообразие жизненных форм внутри таксономических групп различного ранга, необходимо отметить их биоморфологическую полиморфность. Можно сказать только то, что изученные представители родов Hylotelephium, Aizopsis, Pseudosedum и Rhodiola характеризуются длиннопобеговыми биоморфами, а рода Orostachys – полурозеточными, однако, как показали недавние молекулярно-филогенетические исследования (Mayzumi, Ohba, 2004), Orostachys – парафилитическая группа, и вполне вероятно, что единообразие жизненных форм в группе является результатом параллельного развития. Стоит также отметить, что все Rhodiola обладают преимущественно моноподиальным нарастанием, которое, вероятно, следует считать синапоморфным признаком рода.
Современное распространение основных типов биоморф в евроазиатской части ареала подсемейства, свидетельствует о их климатической обусловленности (Böttcher, Jäger, 1984). Наиболее широко распространены корневищные гемикриптофиты, что обусловлено, вероятно, их максимальной приспособленностью к условиям сезонного климата. Интересно, что широколистные летнезеленые многолетники сконцентрированы на восточной оконечности континента, тогда как вальковатолистные терофиты произрастают главным образом на западной его стороне. Около 40 % всех представителей Sedoideae флоры Европы – вальковатолистные терофиты (Böttcher, Jäger, 1984; Hart, 1991).
Понятие жизненной формы неразрывно связано с понятием вегетативной подвижности, включающим в себя интенсивность разрастания, захвата и удержания территории и естественное вегетативное размножение (Смирнова, 1987) [см. также гл. 7.4]. Среди изученных видов Sedoideae мы выделяем следующие группы (терминология и определения по: Смирнова, 1987).
1. Вегетативно неподвижные – каудексообразующие гемикриптофиты и полурозеточные, не образующие дерновинки гемикриптофиты. Разрастание незначительное, только за счет увеличения размеров каудекса или вообще отсутствует у полурозеточных. Полегающие побеги способны укореняться и образовывать дочерние каудексы, за счет этого возможно некоторое продвижение в пространстве, но значительной роли в распространении вида оно не имеет. Вероятна и сенильная партикуляция, она не сопровождается омоложением потомства и не играет существенной роли в самоподдержании популяции. К этой группе нами отнесены: Aizopsis selskiana, Orostachys gorovoii, O. japonicа, O. maximowiczii, О. malacophylla, O. spinosa, Rhodiola rosea ssp. rosea, R. quadrifida, R. borealis, R. pinnatifida, R. pinnatifida ssp. subpinnata, R. rosea ssp. sachalinensis, R. rosea ssp. krivochizhinii, S. albertii.
2. Вегетативно малоподвижные – корневищные гемикриптофиты, короткокорневищные и корнеклубневые криптофиты, а также кустарнички и полукустарнички с прямостоячими побегами. Интенсивное разрастание отсутствует, имеют место сенильная и нормальная партикуляции. По мнению Т.И. Серебряковой (1986), последний тип размножения полезен виду, поскольку обеспечивает некоторое омоложение потомства, и индивид долго может удерживать площадь обитания, но такое размножение не приводит к захвату новых площадей. К данной группе нами отнесены: Aizopsis aizoon ssp. aizoon, A. aizoon ssp. baicalensis, A. litoralis, Hylotelephium erythrostictum, H. pallescens, H. pseudospectabile, H. telephium ssp. stepposum, H. triphyllum ssp. triphyllum, H. triphyllum ssp. sukaczevii, H. ussuriense, Pseudosedum lievenii, Rhodiola algida, R. angusta, R. integrifolia, R. ishidae, R. stephanii.
3. Вегетативно подвижные – эта группа объединяет полукустарнички с приподнимающимися побегами, полурозеточные образующие дерновинки хамефиты, травянистые хамефиты и короткокорневищные криптофиты со специализированным вегетативным размножением:
3а. Виды, подвижность которых обеспечивается интенсивным нарастанием и ветвлением скелетных осей (плагиотропных и укореняющихся). Это, как правило, растения скальных местообитаний. Скальные породы имеют тенденцию к разрушению, и партикуляция дает возможность растению расселяться на движущемся субстрате. Образование «наземных» и эпигеогенных разветвленных корневищ позволяет использовать минимальный поверхностный слой почвы (Aizopsis hybrida, Hylotelephium cyaneum, H. ewersii, H. pluricaule, Orostachys aggregata, O. furusei, O. paradoxa, Sedum acre), а подземных (эпи-, гипогеогенных) корневищ, любые доступные щели между камнями (Aizopsis middendorfiana ssp. middendorfianа, A. kamtschatica, A. kurilensis).
3б. Виды, подвижность которых обеспечивается специализированными вегетативными пропагулами, размножающиеся глубоко омоложенными вегетативными диаспорами, образующимися на различных стадиях онтогенеза (сарментация, гомофазная вивипария). Вегетативные диаспоры часто заменяют семена. Длительность жизни клона становится неопределенно долгой, а интенсивность рассеивания вегетативных диаспор обеспечивает максимальную вегетативную подвижность. К этой группе нами отнесены Hylotelephium verticillatum и H. viviparum, Orostachys spinosa.
Необходимо отметить, что некоторые представители рода Orostachys в зависимости от реализующейся жизненной формы проявляют различную вегетативную подвижность и могут быть отнесены либо к вегетативно неподвижным (при отсутствии вегетативного размножения), либо к вегетативно подвижным (при его наличии).
Анализируя современное распространение видов, характеризующихся различными биоморфами, можно констатировать, что в высокие широты дальше других продвинулись гемикриптофиты, что, вероятно, объясняется лучшей защищенностью их почек возобновления по сравнению с хамефитами и меньшей зависимостью начальных фаз сезонного развития от климатических условий по сравнению с криптофитами (см. гл. 7.5).
К югу, по мере приближения к аридным районам, спектр биоморф сужается, начинают преобладать терофиты, которые, как было сказано выше, образуют южную границу ареала рода, проникая в аридные области, что, по-видимому, свидетельствует о наилучшей адаптированности однолетних жизненных форм к дефициту воды.
7.3. Онтогенез Sedoideae Сибири и российского Дальнего Востока
Жизненные формы являются результатом онтогенеза растения – упорядоченной, генетически обусловленной последовательности развития одной или, в случае вегетативного размножения, ряда поколений особей от диаспоры до естественной смерти, осуществляющейся в сложных связях со средой, по наследственно выработавшейся программе (Серебрякова, 1971; Скрипчинский, 1977; Смирнова, 1987).
Несмотря на значительное разнообразие биоморф Sedoideae, их онтогенез отличается удивительным единообразием и может быть сведен к трем типам: онтогенез симподиально возобновляющихся длиннопобеговых биоморф – «Aizoon» тип с двумя подтипами («Aizoon» и «Sichotense»); моноподиально возобновляющихся длиннопобеговых – «Rhodiola» тип с двумя подтипами («Rosea» и «Angusta»); симподиально возобновляющихся полурозеточных биоморф – «Orostachys» тип с двумя подтипами («Paradoxa» и «Maximowiczii»). Различия выделенных типов заключаются главным образом в специфике фаз морфогенеза, через которые проходит растение в течение жизни.
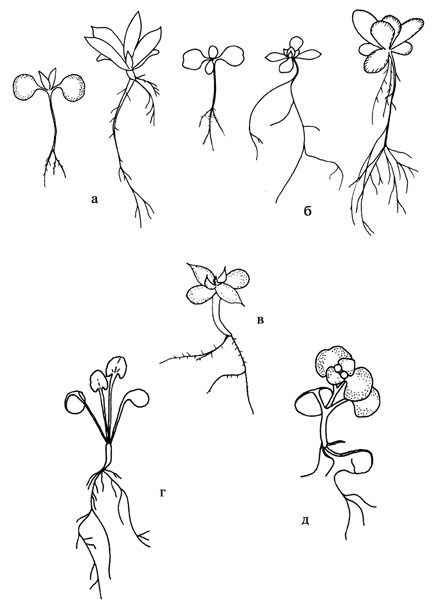
Рис. 37. Проростки и ювенильные растения представителей Sedoideae.а – Orostachys maximowiczii; б – O. malacophylla; в – O. spinosa; д – Rhodiola rosea; е – Aizopsis litoralis
|
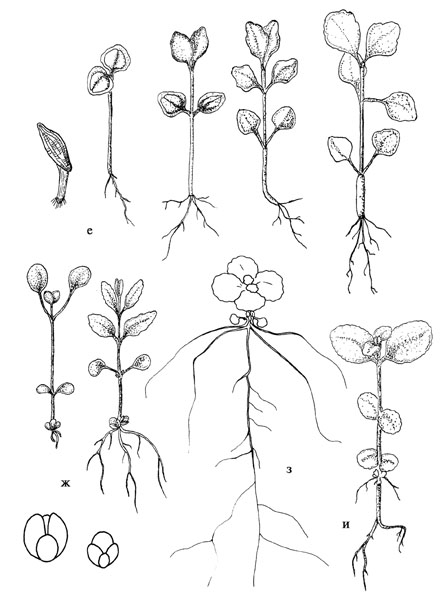
Продолжение рис. 37. е – Aizopsis aizoon; ж – Hylotelephium viviparum; з, и – Hylotelephium pallescens, з – весенний посев, и – подзимний посев. |
В случае раннего прорастания, после образования второй-третьей пары листьев растение переходит в ювенильное состояние (j). Семядоли отмирают, на гипокотиле начинают образовываться придаточные корни, а в пазухах семядолей и первой пары листьев – боковые почки. К концу первого вегетационного периода образуются 8–9 листьев, причем первые пары перекрестно-супротивны, а последующие листья имеют листорасположение, типичное для взрослого растения, то есть очередное с высокими значениями формул филлотаксиса. Наряду с продолжающимся ростом и ветвлением системы главного корня появляются гипокотильные и стеблевые придаточные корни. К концу вегетационного периода гипокотиль и базальная часть втягиваются в почву, образуя зачаток короткого корневища.
При позднем прорастании в первый вегетационный сезон почечка не трогается в рост, первичный побег не развивается, а в пазухах семядолей образуются боковые почки, на гипокотиле – придаточные корни. В таком состоянии растение перезимовывает. К началу следующего вегетационного периода семядоли отмирают. В ювенильное состояние (j) особь переходит в следующий вегетационный период, образуя моноподиально нарастающий одноосный (первичный) побег.
При вступлении в генеративный период онтогенез «Aizoon» – типа разделяется на два подтипа: «Sichotense» и собственно «Aizoon».
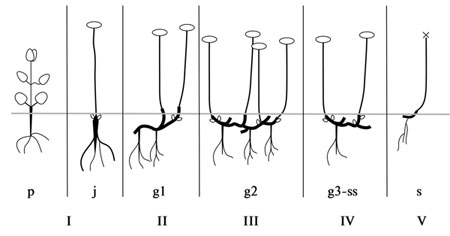
Рис. 38. Онтогенез «Aizoon» подтипа.
I – первичный побег (p-j), II – первичный симподий (g1), III – главный симподий (куст) (g2), IV – партикула (парциальный куст) (g3-ss), V – сенильная партикула (s).
|
«Aizoon» подтип. У корневищных и корнеклубневых геофитов (рис. 38) после отмирания надземной части первичного побега, в следующие (второй-третий) вегетационные периоды особь вступает либо в имматурное (im), либо в генеративное состояние (g1). В рост трогаются почки возобновления, располагающиеся на сохраняющейся прошлогодней (в будущем многолетней) части первичного побега, погруженной в почву. Система главного корня постепенно отмирает, образование корней идет на многолетних частях прошлогодних побегов, наиболее активно – в основаниях монокарпических побегов. Тип корневой системы меняется со стержнекорневой на стержне-кистекорневую, а затем и на кистекорневую (см. также гл. 4).
Во взрослом генеративном состоянии (g2) растение состоит из 2-50 надземных побегов и корневища (эпи-, гипогеогенного), в большей или меньшей степени разветвленного. Процессы новообразования и отмирания уравновешиваются. Среди видов данного типа наблюдается тенденция к уменьшению числа побегов у особи и сокращению длительности жизни многолетних частей корневища. Например, особи Hylotelephium pallescens, Н. viviparum редко имеют более 5 побегов, а корневище обновляется полностью за 2-3 года. |
В последующие (пятый, шестой и т.д.) вегетационные периоды, когда растения достигают старого генеративного и субсенильного состояния (g3-ss), активизируются процессы отмирания. Разрушение начинается в наиболее старых частях корневища и распространяется на более молодые. Для видов с этим типом характерны сенильная (ss) и нормальная партикуляции (g3) (разделение растения на жизнеспособные части). Последняя обеспечивает некоторую вегетативную подвижность и возможность неопределенно долго удерживать территорию. Вегетативное размножение не играет важной роли в жизни этой группы видов, за исключением Н. viviparum и Н. verticillatum, размножающихся, в основном, вегетативно.
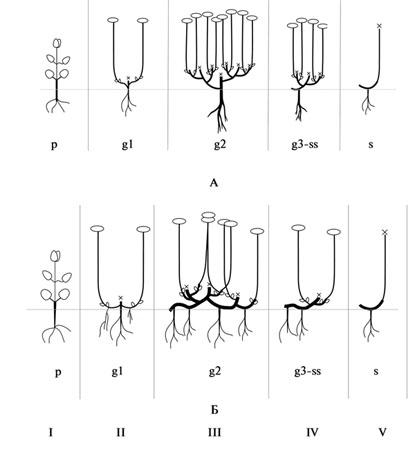
Рис. 39. Онтогенез «Sichotense» подтипа.
А – полудревесных хамефитов; Б – травянистых хамефитов.
I – первичный побег (p-j), II – первичный симподий (g1), III – главный симподий (куст) (g2), IV – партикула (парциальный куст) (g3-ss), V – сенильная партикула (s).
|
«Sichotense» подтип. У полудревесных и травянистых хамефитов (рис. 39) в базальной части первичного побега располагаются 2-3 сближенных узла, формирующих в дальнейшем «скелет» особи (скелетная зона побега). Далее следует удлиненная часть побега – ассимилирующая зона и соцветие. Симподиально ветвясь, особь образует боковые монокарпические побеги (2-го..., N-го порядков). После плодоношения вся «удлиненная» часть побега отмирает, сохраняется только скелетная, несущая почки возобновления. Многолетняя зона каждого монокарпического побега входит в скелетную основу растения, образуя первичный куст. Система главного корня продолжает расти, из базальных частей побегов (подземных и надземных) формируется смешанное корневище. В последующие годы ветвление осуществляется посредством побегов, развивающихся из пазушных почек на скелетных частях побегов.
При вступлении во взрослое генеративное состояние (g2) особь представляет собой сильно разветвленную систему резидов. Система главного корня хорошо развита (Aizopsis middendorfianа ssp. sichotensis) или полностью заменена вторично стержне-кистекорневой системой, сформирован главный симподий (или главный куст).
В последующие годы, у старых генеративных и субсенильных особей (g3-ss) процессы отмирания усиливаются, разрушение начинается в наиболее старой части куста и распространяется к периферии. Происходит старческая (пресенильная) партикуляция.
Сенильное состояние (s) слабо выражено, характеризуется утратой способности к цветению и плодоношению и постепенным отмиранием скелетной системы.
У полукустарничков с приподнимающимися побегами и травянистых хамефитов партикуляция может происходить на более ранних этапах (во взрослом генеративном состоянии), что способствует продлению жизни особи и захвату территорий. |
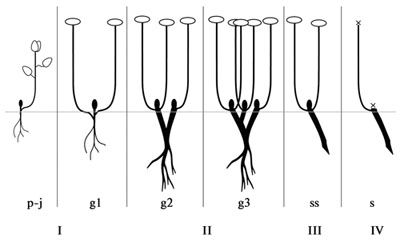
Рис. 40. Онтогенез «Angusta» подтипа.
I – первичный моноподий (p-g1), II – главный моноподий (g2-g3), III – партикула (ss), IV – сенильная партикула (s).
|
«Rhodiola» тип включает в себя онтогенезы каудексообразующих (подтип «Rosea») и моноподиально нарастающих корневищно-стержнекорневых гемикриптофитов (подтип «Angusta», рис. 40). Главное отличие этого типа в том, что верхушечная почка функционирует, как правило, в течение всей жизни растения, цветоносные побеги закладываются в пазухах листьев верхушечной почки. Как и онтогенез «Aizoon» типа, онтогенез «Rhodiola», как правило, простой (редко сложный) и может быть описан такой последовательностью морфогенетических фаз: диаспора – первичный моноподий (p-g1) – главный моноподий (g2-g3) – партикула (ss) – сенильная партикула (s).
После прорастания семян и образования первой пары настоящих листьев верхушечная почка прекращает видимый рост (р). Семядоли могут сохраняться до конца первого вегетационного периода. В пазухах листьев формируются почки, из которых на следующий год образуются 1-3 вегетативных побега. Растение вступает в ювенильный период (j), который может продолжаться несколько вегетационных сезонов. В культуре растения зацветают, как правило, на 2–3-й год, в естественных условиях ювенильный период может растягиваться на многие годы (до 30 лет, по данным В.Н. Андреевой с соавторами, 1987). |
Система главного корня у каудексообразующих биоморф (подтип «Rosea») функционирует в течение всей жизни и в совокупности с многолетними основаниями побегов образует каудекс. Каудекс разрастается радиально за счет увеличения числа резид. На нем образуются придаточные корни, и имеется большой запас спящих почек. Из почек ежегодно развиваются 3-10 и более побегов, растения вступают в генеративный период (g), который может продолжаться более сотни лет. По данным Е.Л. Нухимовского с соавторами (1987), самый крупный куст Rhodiola rosea, найденный в естественных условиях в высокогорьях Алтая, имел 249 побегов и возраст около 200 лет. Генеративные органы закладываются в почках либо в конце вегетационного периода, предшествующего цветению, либо за год–три до цветения (Андреева и др., 1987). При погружении в субстрат отмирающих обычно частей побегов может происходить образование дочерних каудексов. Нередко наблюдается травматическая партикуляция. Каудексы разламываются при механических воздействиях (движении грунтов, снеготаянии, повреждении животными), и отделившиеся части (партикулы) укореняются.
По мере старения каудекса имеет место ослабление побегообразования, нередко наблюдается сенильная партикуляция. «Каудекс отличается от корневища тем, что не отмирает в базальной части, а вместе с первичной корневой системой функционирует в течение всей его жизни» (Серебряков, Серебрякова, 1965, с. 80). У видов данной жизненной формы, в отличие от всех вышеназванных, отсутствует уравновешивание процессов нарастания и отмирания подземной части, нарастание продолжается вплоть до наступления сенильного периода.
У моноподиально нарастающих корневищно-стержнекорневых гемикриптофитов, как отмечено выше, главный корень также сохраняется значительную часть жизни, однако веточки каудекса значительно превышают его по длине, а часто главный корень полностью отмирает, растение становится корневищным. Онтогенез «Angusta» подтипа в целом сходен с описанным выше «Aizoon» подтипом. Отличия заключаются в типе нарастания корневища, верхушечная почка функционирует большую часть жизни. При неблагоприятных условиях может происходить гибель верхушечной почки, образуются «гемисимподиальные» с вынужденным перевершиниванием побеги (Нухимовский и др., 1987).
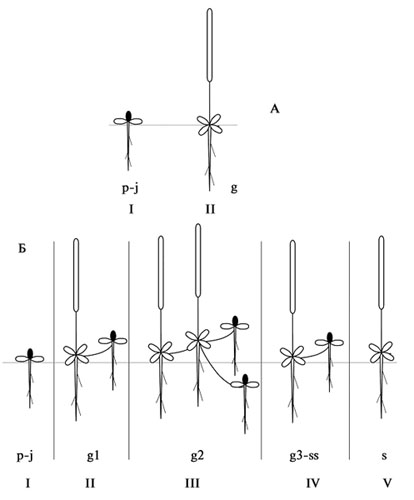
Рис. 41. Онтогенез «Orostachys» типа.
А – «Maximowiczii» подтип. I – вегетативное розеточное растение (p-j), II – генеративное (вегетативно невозобновляющееся) полурозеточное растение (g).
Б – «Paradoxa» подтип. I – вегетативное розеточное растение (p-j); II – полурозеточное, III – вегетативно возобновляющееся генеративное растение (g1-g2), IV – партикула (g3-ss), V – генеративного (вегетативно невозобновляющегося) полурозеточного растения (s).
|
«Orostachys» тип включает в себя онтогенезы симподиально возобновляющихся полурозеточных биоморф [полурозеточных хамефитов, гемикриптофитов и терофитов] (рис. 41). В зависимости от экологических условий представители рода Orostachys могут проходить различные варианты онтоморфогенеза: простой онтогенез без смены поколений («Maximowiczii» подтип) и сложный со сменой поколений («Paradoxa» подтип). В первом случае цикл развития складывается из фаз: вегетативного розеточного растения (p-j) и генеративного (вегетативно невозобновляющегося) полурозеточного растения (g). Во втором случае – из фаз: вегетативного розеточного растения (p-j); полурозеточного, вегетативно возобновляющегося генеративного растения (g1-g2), партикулы (g3-ss) и генеративного (вегетативно невозобновляющегося) полурозеточного растения (s).
Семена Orostachys прорастают ранней весной, прорастание, как и у представителей других родов и биоморф, дружное, но отдельные проростки могут появляться в течение нескольких лет. Сидячие семядольные листья могут либо сохраняться до конца первого вегетационного периода, либо отмирать. Главный корень может отмирать или сохраняться до конца жизни растения, но, как правило, он не выделяется из общей массы придаточных корней. Первые настоящие листья образуются через 4-6 недель после прораста-ния, растение вступает в фазу вегетативной розетки, в которой может находиться от одного до 3 (редко более, O. spinosa) лет, в редких случаях в текущий вегетационный период из верхушечной почки образуется колосовидное олиственное соцветие, и растение вступает в фазу генеративного (вегетативно невозобновляющегося) полурозеточного растения, отмирая после плодоношения (O. maximowiczii). В типичном случае растение вступает в генеративный период на второй-третий год и также отмирает после цветения и плодоношения. Полный цикл развития завершается в течение 1-5 лет («Maximowiczii» тип), сенильный период не выражен.
«Paradoxa» подтип (рис. 41, Б). В течение первого вегетационного периода образуется вегетативное розеточное растение (p-j), во второй год из апикальной почки формируется олиственное соцветие, растения цветут и плодоносят, из пазушных почек формируются новые вегетативные розетки. Генеративный побег отмирает после плодоношения почти до основания, оставшаяся часть побега входит в состав наземного корневища. Постепенно из многолетних частей слагается сильно разветвленное наземное корневище с большим количеством вегетативных розеточных и генеративных полурозеточных побегов (g-g2) (дерновинка или латка). Вегетативные розетки могут образовываться на столоновидных побегах, вырастающих из пазух листьев, также формируя более или менее плотную дерновинку. В последующие годы начинается разрушение корневищ, отмирание столонов и образование многочисленных партикул (g3-ss). Растения существуют в форме «скрытого» клона (термин Е.Л. Нухимовского, 1987). В таком состоянии растения могут существовать бесконечно долго. Сенильное состояние выражено слабо и заключается в ослаблении цветения и плодоношения и снижении интенсивности вегетативного возобновления и размножения. |
7.4. Репродуктивная биология Sedoideae
Репродуктивная биология – область ботаники, изучающая процессы возобновления и размножения растений. Возобновление – понятие, относящееся к популяции растений, размножение – к растению как организму (Левина, 1981; Терехин, 2000).
Размножение – совокупность процессов, приводящих к увеличению числа потомков. Основные типы размножения цветковых растений – половое и бесполое, первое включает в себя главным образом семенное, второе – вегетативное. Вегетативное размножение делится на размножение специализированными органами (вегетативная вивипария, сарментация и т.д.) и неспециализированными [партикуляция] (Шорина, 2000а).
Для большинства представителей как Crassulaceae, так и Sedoideae семенное размножение – основной способ размножения, однако большое значение в существовании видов семейства и самоподдержании популяций имеют и различные способы вегетативного размножения.
Семенное размножение Sedoideae
Опыление. Sedoideae имеют насекомоопыляемые, хазмогамные цветки, перенос пыльцы осуществляется насекомыми-опылителями в открытом цветке. Как мы уже отмечали выше (гл. 4), цветки большинства изученных представителей Sedoideae обоеполые, только представители подрода Rhodiola рода Rhodiola имеют раздельнополые цветки и являются двудомными, что определяет особенности репродуктивной биологии видов подрода. Отдельные цветки очитковых довольно мелкие, их диаметр редко превышает 5-7 мм, однако они у большинства видов яркие (желтые, белые или малиновые) и собраны в крупные (иногда достигающие 15-20 см в диаметре) соцветия, привлекающие насекомых-опылителей. Бледные зеленоватые цветки характерны для видов, размножающихся преимущественно вегетативно (Hylotelephium verticillatum, H. viviparum). Привлечению опылителей способствует и обильное выделение нектара.
Представителям Crassulaceae свойственно как перекрестное, так и самоопыление. Цветки Crassulaceae, как правило, протандрические, редко протерогинические (Spongberg, 1978; Виноградова, 1981; Бялт, 2001), то есть в большинстве случаев пыльца созревает раньше, чем рыльце готово воспринять ее, что препятствует самоопылению и в определенной степени способствует перекрестному. Причем первыми раскрываются пыльники наружного круга тычинок и спустя некоторое время – второго, что удлиняет продолжительность периода выбрасывания пыльцы и увеличивает вероятность успешного опыления. Особенности строения соцветий (см. гл. 4), разновременность зацветания отдельных цветков позволяют насекомым переносить пыльцу как в пределах одного соцветия (гейтоногамия), так и между соседними растениями (ксеногамия). С этих позиций колосовидные соцветия представителей Orostachys более приспособлены к перекрестному опылению, поскольку, по нашим наблюдениям, насекомые-опылители, двигаясь снизу вверх, сначала посещают цветки, находящиеся в женской фазе развития, а затем – в мужской, более молодые и расположенные выше по соцветию, что практически полностью предотвращает самоопыление. Однако и в данном случае сохраняется его возможность как резервного способа опыления, вступающего в действие при отсутствии перекрестного.
При двудомности растений самоопыление полностью исключено. В подсемействе Sedoideae двудомность характерна для видов, обитающих, главным образом, в условиях высокогорий или морских побережий (подрод Rhodiola рода Rhodiola). По мнению H. Ohba (1978), двудомность – более прогрессивный признак, чем двуполость или однодомность, однако, на наш взгляд, полное исключение возможности самоопыления ставит и популяцию, и вид в уязвимое положение.
Как само-, так и перекрестное опыление играют важную роль в жизни вида. Перекрестное опыление повышает уровень гетерозиготности и ведет к эволюционным преобразованиям популяций, образованию новых форм организации, а самоопыление повышает уровень гомозиготности и ведет к распаду вида на множество стойких форм (Шмальгаузен, 1983; Демьянова, 2000).
Прорастание семян. Свежесобранные семена исследованных видов очитковых, в своем большинстве, имеют высокую лабораторную всхожесть (Гончарова, Абанькина, 1999). Прорастание семян на свету незатрудненное и у большинства видов начинается на 2–3-й (при t +24оС) или 4–6-й (при t+18-20оC) день опыта. В зависимости от температурных условий проращивания максимальная всхожесть наблюдается соответственно на 2–4-й или 6–8-й день от начала прорастания. Прорастание семян может быть дружным (5–9 дней) или растянутым (20–90 дней), единичные всходы могут появляться спустя год после посева. Незатрудненное прорастание семян изученных Sedoideae обусловлено их строением: тонкими покровами, крупным дифференцированным зародышем, в значительной степени утилизирующим в процессе своего развития питательные вещества запасающих тканей (Гончарова, Абанькина, 1999).
И хотя прорастание семян в лабораторных условиях незатрудненное, в естественных условиях семенам изученных очитковых свойствен покой, обусловленный внешними факторами и обеспечивающий прорастание семян при наступлении максимально благоприятных условий, как правило, весной следующего после созревания вегетационного периода.
Семена Sedoideae светочувствительны. Это выражается в полном отсутствии прорастания (Hylotelephium pallescens, Н. ussuriense) или появлении единичных проростков (Aizopsis selskiana, А. aizoon, А litoralis, A. middendorfiana subsp. sichotensis) в темноте. Подобная светочувствительность рассматривается как неглубокий физиологический покой [по классификации М.Г. Николаевой с соавторами (1985)]. Он характеризуется обычно временным сужением диапазона как световых, так и температурных условий прорастания. Причина сужения – особое физиологическое состояние зародыша, проявляющееся в пониженной ростовой активности, сочетающееся с недостаточной газопроницаемостью тканей, непосредственно его окружающих (Муйжарая и др., 1983; Николаева и др., 1985; Kinzel, 1913).
Покой, обусловленный свойствами покровов семян, отмечаемый у семян некоторых европейских видов Sedum s.l. (Николаева и др., 1985), у изученных нами Sedoideae не выявлен (Гончарова, Абанькина, 1999). Тонкая спермодерма и маломощный эндосперм не являются препятствием для набухания и прорастания семян.
Условия созревания и хранения семян оказывают большое влияние на прорастание и сохранение ими всхожести. Так, свежие семена Aizopsis litoralis из прибрежных местообитаний, собранные нами в 1998 г., обладали невысокой (24-26 %) лабораторной всхожестью, а в некоторых случаях она составляла только 6-8 %. Вероятно, причиной этого явилось углубление физиологического покоя, индуцируемое условиями дозаривания семян, складывающимися в естественных местообитаниях вида на морском побережье (значительная инсоляция, сильный ветер) и приводящими к повышенному иссушению семян. Воздействие на такие семена гибберелином (100 мг/л, 50 мг/л, 24 часа), замачивание в KMnO4 и H2O2 (24-48 часов), а также скарификация не оказывали заметного влияния на всхожесть. Положительный эффект давала длительная холодная стратификация (1-3 месяца), в результате которой всхожесть возросла до 76%.
При хранении в лабораторных условиях (t+18о … + 20оС, влажность 30-50 %) семена Hylotelephium и Orostachys, созревающие в октябре-ноябре, сохраняют высокую (более 50 %) всхожесть в течение 35 недель, а семена представителей Aizopsis и Rhodiola, созревающие в июле-сентябре, – до 20 недель. Снижение жизнеспособности семян Sedoideae при сохранении еще высокой всхожести проявляется в удлинении периода прорастания и/или снижении дружности прорастания, а в дальнейшем и в появлении проростков с различными аномалиями. Через год хранения в лабораторных условиях семена изученных видов очитковых становятся невсхожими. Относительно быстрая потеря жизнеспособности семенами исследованных видов также обусловлена их строением.
Хранение семян Sedoideae при температуре +40С позволяет сохранить высокую всхожесть (более 50%) до двух лет, а у некоторых Hylotelephium и Orostachys – до четырех, Rhodiola – до 7-8 лет (Андреева и др., 1987).
В лабораторных условиях очитковые обладают высокой потенциальной всхожестью, но в естественных условиях редко можно найти проростки. Объясняется это, по-видимому, тем, что семена редко попадают в благоприятные для прорастания и развития условия, а часть проросших, вероятно, погибает. Кроме того, семена очитков светочувствительны и требовательны к количеству влаги. Излишнее заглубление или сильное высушивание ингибируют прорастание и дальнейшее развитие проростков.
Вегетативное размножение – увеличение числа особей одного растения в результате отделения от материнского растения специализированных или неспециализированных органов. Изученным очитковым присуще как специализированное (вивипария, сарментация), так и неспециализированное (партикуляция в широком смысле, включая парциаляцию) вегетативное размножение.
Неспециализированное вегетативное размножение
Партикуляция – любой процесс разделения вегетативного тела растения на жизнеспособные части (Васильев и др., 1988). В зависимости от стадии онтогенеза, на которой она происходит, выделяют сенильную и нормальную партикуляции (Работнов, 1969; Смирнова, 1974, 1987; Васильев и др., 1988; Шорина, 2000а).
Сенильная партикуляция происходит при старческом распаде особи, она не сопровождается омоложением потомства, новообразований не происходит, части клона имеют пониженную жизнеспособность. И хотя и происходит увеличение числа особей, как нами отмечено выше, этот тип размножения не имеет существенного значения ни в самоподдержании популяции, ни в захвате новых площадей, ни в распространении видов очитковых. Среди Sedoideae сенильная партикуляция обычна для каудексообразующих и подушковидных гемикриптофитов (Aizopsis selskiana, Rhodiola rosea ssp. rosea, R. coccinea R. quadrifida).
Нормальная партикуляция происходит на генеративной стадии онтогенеза, обычен такой тип размножения у корневищных гемикриптофитов, короткокорневищных и корневищно-корнеклубневых криптофиты, полукустарничков, травянистых хамефитов (Rhodiola angusta, R. integrifolia, Aizopsis aizoon, A. middendorfianа ssp. sichotensis, Hylotelephium erythrostictum, H. pseudospectabile, Sedum acre и др.). Как мы уже отмечали, этот тип размножения позволяет виду бесконечно долго удерживать площадь обитания, но не приводит к захвату новых площадей.
Crassulaceae вообще и Sedoideae в частности имеют поразительную способность к регенерации, в связи с этим травматическая партикуляция (разделение материнской особи на жизнеспособные неспециализированные части при воздействии человека, животных или природных факторов) – обычный способ неспециализированного вегетативного размножения толстянковых – также имеет существенное значение в жизни как особи, так и популяции.
Специализированное вегетативное размножение
Сарментация – специализированный способ вегетативного размножения, при котором происходит образование отделяющихся от материнского растения отпрысков (Барыкина, 2000; Батыгина, Васильева, 2002). Этот тип размножения очень широко распространен среди толстянковых, однако среди изученных очитковых он довольно редок и встречается главным образом у представителей рода Orostachys (см. также гл. 4). У видов этого рода часто образуются легко отделяющиеся от материнского растения дочерние розетки. При сарментации, как и гомофазной вивипарии, дочерние особи резко омоложены (онтогенетическое состояние соответствует ювенильному) и выживаемость их очень велика.
Вивипария. Объем понятия «вивипария» и классификация ее типов и до сих пор остаются дискуссионными, мы, вслед за авторами «Эмбриологии цветковых растений…» (Батыгина, Брагина, 2000), различаем гетерофазную и гомофазную вивипарию – генеративную и вегетативную соответственно. В результате живорождения генеративная или вегетативная диаспора образует без периода покоя проростки еще на материнском растении (Батыгина, Брагина, 2000).
У толстянковых спорадически бывают случаи генеративной вивипарии, когда, в результате стрессового изменения влажности или температуры семена в листовках прорастают на растении до высыпания на землю.
Инфлоральная гомофазная вивипария свойственна Hylotelephium viviparum – облигатно живородящему виду – вегетативные диаспоры в массе формируются на соцветии, однако, хотя и очень редко, наряду с вегетативными диаспорами растения образуют и единичные жизнеспособные семена. В конце вегетационного периода пропагулы опадают с материнского растения и укореняются, покровные листочки выводковой почки выполняют одновременно защитную и трофическую функции. Весной пропагула, подобно семени, прорастает, «проросток», развивающийся из нее, выглядит идентично настоящим проросткам, развивающимся из семян (рис. 37, ж).
Репродуктивная стратегия Hylotelephium verticillatum сочетает семенное размножение и каулигенную вивипарию, вегетативные диаспоры образуются в пазухах листьев и в конце вегетационного периода, как и у Hylotelephium viviparum, опадают и укореняются (приложение 16), растения выглядят сходно с ювенильными особями, развившимися из семян близкородственных видов. Необходимо отметить, что гомофазная вивипария довольно часто встречается в семействе в родах Sedum s. str., Echeveria, Bryophyllum и Kalanchoe, не связанных между собой филогенетически и, возможно, является результатом параллельной эволюции.
Значение вегетативного размножения для Sedoideae. Принято считать, что вегетативная вивипария является реакцией на стрессовые условия обитания, она наиболее характерна для растений, произрастающих в полярных, высокогорных и пустынных районах, с коротким вегетационным периодом (Тихомиров, 1963; Юсуфов, 1972; Батыгина, Брагина, 2000). Появление специализированных типов вегетативного размножения у представителей Sedoideae является, вероятно, реакцией на влияние биотических факторов, а именно – конкуренции с другими видами иных жизненных форм, обитающих в ценозе. Вегетативные диаспоры очитковых более конкурентоспособны, чем генеративные (семена), и менее требовательны к свету, влаге; способны расти на задерненных участках. Подобно семенам, вегетативные диаспоры в массе возникают на растении и, опадая, дают начало новым особям, значительно омоложенным по сравнению с материнскими (Любарский, 1961; Васильев и др., 1988). Онтогенетическое состояние вегетативных зачатков Hylotelephium viviparum соответствует состоянию проростка, а H. verticillatum – ювенильного растения, потомство их сильно омоложено по сравнению с материнскими растениями и, соответственно, более жизненно.
Необходимо отметить, что виды, обитающие в одинаковых условиях, далеко не всегда имеют одинаковые репродуктивные стратегии. Например, H. viviparum и H. pallescens, произрастая часто в сходных местообитаниях, характеризуются различными репродуктивными стратегиями, первый вид размножается преимущественно вегетативно, второй – генеративно. Вероятно, современная стратегия вида является отражением не только условий его обитания, но и истории формирования, а также проявлением принципа комплементарности членов одного сообщества.
Эволюционное значение вегетативного размножения для самоподдержания вида также вызывает массу дискуссий. Одни авторы (Джапаридзе, 1963) считают его «эволюционно обреченным», другие (Юсуфов, 1972) – «эволюционно перспективным». Мы полностью согласны с мнением Р.Е. Левиной (1981, с.11), что вегетативное размножение «сохранилось и специализировалось у всех групп растений как самый надежный механизм, страхующий вид от вымирания». Несомненным преимуществом вегетативного размножения является «сохранение благоприятного генотипа и увеличение его численности» (Юсуфов, 1972, с. 396), однако отсутствие генотипической изменчивости ограничивает его эволюционную перспективность, в том числе и для представителей Sedoideae.
7.5. Ритмы сезонного развития
Периодичность жизненных процессов растений выражается в закономерно повторяющихся морфологически различных этапах развития. Одно из проявлений периодичности – сезонные и годичные циклы, связанные с чередованием периодов роста и покоя. У растений сезонных климатов периодичность роста и развития обусловлена главным образом климатическими факторами, основными из которых являются температура, влажность и длина дня (Серебряков, 1966, Шульц, 1981, Юсуфов, 1996).
Рост и развитие организма - результат взаимодействия между эндогенными и экзогенными факторами, то есть между его внутренней организацией и внешней средой (Любименко, 1924; Шульц, 1981; Стародубцев, 1984). Скоррелированность взаимодействия наследственно закрепленных свойств организма и современных климатических условий и определяет ритм развития растений - их феноритмотип. Феноритмотип, по определению И.В. Борисовой (1972), - группа видов со сходными длительностью и сроками начала и конца вегетации, сходными ритмами смен вегетации и покоя.
Sedoideae флоры Сибири и российского Дальнего Востока - длительно вегетирующие растения, активная вегетация которых продолжается всю или большую часть безморозного периода года. Анализ хода сезонного развития изученных видов очитковых позволяет нам выделить три различных феноритмотипа: 1) осенне-зимне-летнезеленый - характерный для хамефитов, биологически вечнозеленых, несущих ассимилирующие зеленые органы в течение всего года; 2) весенне-летне-осеннезеленый - характерный для гемикриптофитов и криптофитов, вегетирующих в течение безморозного периода года; 3) весенне-летнезеленый (гемиэфемероиды) - для каудексообразующих и короткокорневищных гемикриптофитов из рода Rhodiola, их вегетация заканчивается задолго до наступления холодов.
В сезонном цикле развития изученных Sedoideae выделяются фазы: весеннего отрастания, активного роста побегов, бутонизации, цветения, плодоношения, отмирания монокарпических надземных побегов, зимнего покоя, а у хамефитов дополнительно к названным - фаза осеннего развития монокарпических побегов.
Функционирование монокарпического побега у хамефитов (Aizopsis hybrida, А. middendorfiana, Hylotelephium pluricaule, H. cyaneum, Orostachys, Sedum acre и др.) начинается осенью. В зимний период рост затормаживается, и растение вступает в фазу «относительного» покоя. До начала активного роста побег находится в фазе «розетки» - открытой ассимилирующей почки возобновления. Такие розетки несут как нормально развитые, так и недоразвитые, мелкие, зеленые, фотосинтезирующие листья. Начало весеннего отрастания видов данной группы совпадает с устойчивым переходом средней температуры воздуха через 0оС (24.III–1.IV). Продолжительность периода активной вегетации хамефитов - 160–180 (200) дней, что практически соответствует безморозному периоду, который продолжается с момента устойчивого перехода средней температуры через 0оС и до первых заморозков - на юге Приморского края с третьей декады марта по третью декаду октября–первую декаду ноября (Василюк, 1975; Деркачева, Русанов, 1990; Туркеня, 1979, 1991).
Криптофиты и гемикриптофиты (Aizopsis aizoon, Hylotelephium viviparum, H. pallescens, Rhodiola и др.) начинают рост ранней весной с момента устойчивого перехода суточных температур через 5оС (13–22 апреля) и заканчивают поздней осенью (19–30 октября). Переход суточных температур через 0оС и 5оС разделяет почти месяц, и, соответственно, почти на месяц позже, чем хамефиты, начинают отрастать очитковые этих жизненных форм (во 2–3-й декаде апреля), что, вероятно, связано с полным оттаиванием почвы. В отдельные годы с поздней весной и быстрым переходом температуры воздуха между 0о и 5оС активная вегетация видов всех жизненных форм начинается почти одновременно.
Период активной вегетации криптофитов и гемикриптофитов из родов Aizopsis и Hylotelephium ограничивается первыми заморозками, а гемикриптофиты из рода Rhodiola, как отмечено выше, заканчивают вегетацию задолго до наступления морозов, что, на наш взгляд, является отражением формирования видов с весенне-летнезеленым феноритмотипом в условиях с коротким вегетационным периодом, обычным для типичных высокогорных и арктических местообитаний представителей рода Rhodiola.
Сравнение сроков развития растений в условиях культуры и естественных местообитаниях подтверждают зависимость начала фазы вегетации от температуры воздуха. Например, различия в наступлении фенофаз у Rhodiola angusta, произрастающей на высоте около 1800 м над ур. моря, в условиях культуры и естественных местообитаниях составляют почти месяц. Отмечена также существенная разница в сроках наступления фенофаз у Rhodiola rosea. Так, например, в условиях Мурманской области начало отрастания этого вида колеблется с 31 мая по 23–25 июля, что связано с полным сходом снега с поверхности почвы (Андреева и др., 1987), начало цветения в условиях г. Владивостока наступает во второй декаде мая, а на вершине г. Облачная – во второй декаде июня. То есть происходит синхронизация эндогенного ритма и существующих условий местообитаний. В суровых условиях высокогорий растения вырабатывают приспособления (осенняя закладка генеративных органов), способствующие ускоренному прохождению цикла развития, однако в годы с низкими летними температурами и частыми снегопадами растения могут вообще не вегетировать, отрастая при наступлении благоприятных условий (Павлов, 1987).
Период активного роста побегов всех видов изученных очитковых ограничивается фазами бутонизации и цветения, только у представителей Rhodiola все три фазы идут одновременно, растения выходят из зимы с полностью сформированными соцветиями и иногда даже с окрашенными бутонами. Для видов родов Aizopsis период активного роста составляет 45-70 дней, родов Orostachys и Hylotelephium - 70-120.
В конце весны один за другим зацветают виды рода Rhodiola, а затем в начале лета рода Aizopsis. Во Владивостоке первыми зацветают Rhodiola rosea subsp. rosea, Rh. rosea subsp. sachalinensis (1-2 декада мая), Aizopsis middendorfiana subsp. sichotensis (1–2-я декада июня), за ним - A. kurilensis и A. middendorfiana subsp. middendorfiana (3-я декада июня–1-я декада июля), затем - A. litoralis, A. aizoon и A. selskiana (1-я декада июля). Позже начинают бутонизировать и цвести виды рода Hylotelephium (2–3-я декада июля) и Orostachys (3-я декада августа). Вероятно, видам двух последних родов для вступления в генеративную фазу необходимы большие суммы активных температур, а для Orostachys - и гораздо меньшая продолжительность дня, чем для представителей других родов. Общая продолжительность цветения каждого вида изученных очитковых составляет 30–50 дней.
Представители родов Aizopsis и Rhodiola, вероятно, имеют органический покой и естественно ограниченный вегетационный период. Независимо от наступления и отсутствия заморозков в условиях культуры виды Rhodiola заканчивают вегетацию к 3-й декаде августа, виды Aizopsis - к 3-й декаде октября. Большинство видов родов Hylotelephium и Orostachys также имеют органический покой, но вынужденно ограниченный вегетационный период, прерываемый заморозками. Представители Aizopsis и Rhodiola имеют четко выраженную зимнюю диапаузу и, даже при выращивании в условиях постоянных, высоких положительных температур (более 10оС) на протяжении всего года, в зимний период уходят на покой. Торможение процессов жизнедеятельности вызвано, на наш взгляд, условиями короткого дня и является приспособлением к условиям сезонного климата с четко выраженной сменой благоприятного и неблагоприятного периодов. Некоторые виды Hylotelephium (H. triphyllum, H. pallescens) способны развиваться, цвести и плодоносить без периода покоя, однако часто наблюдается ослабление цветения и плодоношения, а иногда и полная гибель растений. Hylotelephium pluricaule также может вегетировать при благоприятных условиях в течение круглого года, но для закладки генеративных органов ему необходимо воздействие отрицательных температур. Подобное явление описано В.В. Скрипчинским и Вл.В. Скрипчинским (1961) для раннецветущих растений Северного Кавказа. Очевидно, подобная реакция имеет приспособительное значение, обеспечивая оптимальную ритмику. Сформировавшиеся генеративные органы без воздействия низких температур не могут зацвести.
Разные фазы сезонного развития отличаются по чувствительности к условиям среды и выделяются как критические по отношению к свету, влажности и температуре (Юсуфов, 1996). Основными лимитирующими факторами, контролирующими начало вегетации изученных очитковых, являются, на наш взгляд, продолжительность дня и температура воздуха и почвы, причем в годы с теплой и ранней весной наступление фенофазы ограничивает длина дня, а с холодной и затяжной - температура воздуха. Опыт по выращиванию Hylotelephium telephium в условиях различной длины дня (8 и 16 часов) и различной температуры (от 9 до 21оС) (Hart, Arkel, 1985) показал, что различия в экзогенных факторах вызывают разные реакции у растений. Например, при коротком дне останавливается рост побегов и соцветия не развиваются, однако закладывается в три раза больше почек возобновления, чем у образцов, выращиваемых при длинном дне (ДД). Вторичная ксилема растений ДД содержала значительно большее количество лигнифицированных волокон. Аналогичный опыт по выяснению влияния длины дня на покой Hylotelephium (Sedum) telephium и Rhodiola rosea (Heide, 2001) показал, что покой контролируется главным образом длиной дня, критическая длина дня, активирующая начало роста, - 15 часов.
Критическим фактором для инициации цветения и плодоношения, на наш взгляд, является длина дня. Часто для развития растений важными оказываются не только длина дня, но и характер его сезонного изменения (укорачивающийся или удлиняющийся день). Так, виды родов Aizopsis и Hylotelephium или Rhodiola и Orostachys цветут при примерно одинаковой длине дня, однако Aizopsis и Rhodiola зацветают при удлиняющемся дне, а Hylotelephium и Orostachys - при укорачивающемся.
Сезонные ритмы отражают не только современные условия, но и условия формирования вида в прошлом (Серебряков, 1966). Известно, что все приспособительные особенности растений, в том числе и сезонная ритмика, - результат развития организма в определенных исторических условиях (Серебряков, 1964; Трулевич, 1991). По мнению Т.И. Серебряковой (1976), виды с зимующими листьями - показатель древности ценозов и мягкости климатических условий, в которых сформировался данный феноритмотип. Вероятно, происхождение представителей осенне-зимне-летнезеленых феноритмотипов может быть связано с районами с теплым бессезонным климатом. Формирование же сезонных типов (весенне-летне-осеннезеленых и весенне-летнезеленых), имеющих естественно ограниченный вегетационный период, связано с сезонным циклическим климатом. Органический покой, по мнению В.Н. Любименко (1924), - удачное приспособление к низким температурам умеренного климата. Чередование периодов покоя и роста позволяет растению переносить недостаток воды и тепла, а необходимость индуцирующего воздействия внешней среды для возобновления роста (температуры и продолжительности дня) предохраняет растения от гибели во время некоторых временно благоприятных периодов («бабьего лета», продолжительных оттепелей). Фотопериодический контроль зимнего покоя имеет большое значение в условиях сезонного климата, так как температура может значительно изменяться в разные годы, длина дня всегда остается постоянной и является идеальным сигналом для синхронизации развития растений и сезонных изменений (Heide, 2001).
Свойственный очитковым покой семян также является проявлением эндогенной сезонной ритмичности. В ходе приспособления к условиям местообитаний, прежде всего, происходит синхронизация начальных этапов онтогенеза (прорастание семян, весеннее отрастание и т.д.), обеспечивающая максимально эффективное использование вегетационного периода, всходы развиваются с наступлением благоприятных условий, вырабатываются механизмы, исключающие опережение или запаздывание в прорастании семян (Юсуфов, 1996).
7.6. Возможные пути биоморфологической эволюции в подсемействе Sedoideae
К сожалению, не существует данных, свидетельствующих о времени и месте происхождения Crassulaceae, не известны и ископаемые остатки толстянковых, которые могли бы подтвердить или опровергнуть существующие гипотезы развития семейства. Однако сравнительный филогенетический анализ позволяет предположить, что Crassulaceae произошли примерно 70 млн лет назад (Crepet et al., 2004) в конце мезозойской-начале кайнозойской эры. К этому времени приходится и начало формирования осевых хребтов современных Альп, Кавказа, Гималаев (Монин, 1977), с которым, по нашему мнению, тесно связан процесс диверсификации семейства. Наиболее вероятно, что Sedoideae (sensu Berger) сформировались политопно и автохтонно на разных континентах в результате параллельной эволюции слабодифференцированных предковых форм. В пользу этой гипотезы свидетельствуют филогенетические исследования различных авторов (Ham, 1995; Ham, Hart, 1998; Mort et al., 2001, настоящее исследование), доказавшие полифилитичность подсемейства sensu Berger и предположившие относительную независимость развития его основных ветвей. Вероятно, что восточно-азиатская ветвь Sedoideae выделилась в результате широтных миграций анцестральных видов из районов Средиземноморья или Южной Африки - предполагаемых центров происхождения семейства (Ham, Hart, 1998). Дальнейшая диверсификация происходила, по-видимому, независимо во всех центрах разнообразия подсемейства, о чем свидетельствует состав современных флор очитковых различных континентов (см. гл. 6). Комплекс морфологических, анатомических и физиологических признаков однозначно свидетельствует, что ведущим фактором эволюции как семейства, так и подсемейства явился дефицит влаги.
Мы уже отмечали, что максимальное число видов и наибольшее разнообразие жизненных форм Sedoideae сосредоточены в Гималаях, Альпах и на Ирано-Туранском нагорье, в областях с высокогорными альпийскими и субальпийскими сообществами умеренных широт. Это, на наш взгляд, позволяет предположить, что эволюция Sedoideae может быть тесно связана с процессом орогенеза. Кроме того, большинство из существующих ныне видов приурочено к открытым скальным, каменистым и щебнистым, как правило, хорошо дренированным экотопам, что также свидетельствует в пользу горного происхождения Sedoideae.
Особенности современного распространения позволяют предположить, что интенсивное развитие Rhodiola связано с формированием Гималаев и «восточно-азиатского высокогорного коридора», приходящимся на неоген (примерно 25 млн лет назад) (Монин, 1977).
Максимальное биоморфологическое разнообразие также приходится на области с высокогорными сообществами, характеризующимися резкими колебаниями дневных и ночных температур, небольшим количеством осадков, низким давлением, сильными ветрами и недостаточным количеством питательных веществ в почве. Своеобразные условия и повлекли за собой формирование разнообразных суккулентных, криофильных форм, а высокий уровень солнечной инсоляции способствовал развитию низкорослых и розеточных биоморф. Как отмечено выше (гл. 6), именно разнообразие климатических и эдафических условий горных субтропических стран послужило причиной образования большого числа как видов, так и жизненных форм, встречающихся в этих областях. В процессе поднятия гор предковые формы оказывались в разных высотных поясах, имеющих существенно отличающиеся экологические условия. В результате освоения все новых и новых местообитаний вырабатывались приспособления к существованию в различных экологических нишах. Молодые Sedoideae могли эволюционировать в нескольких направлениях, одни из них осваивали высокогорные участки (Rhodiola, каудексообразующие и моноподиальные биоморфы, моноподиально розеточные модели побегообразования); другие - открытые, сухие с высокой инсоляцией скалистые и каменистые участки (Orostachys, розеточные и полурозеточные биоморфы, розеточные формы роста); третьи - расположенные ниже, более или менее мезофитные местообитания (большинство Hylotelephium и Aizopsis, корневищные биоморфы, симподиально длиннопобеговые формы роста).
По нашему мнению, можно говорить о сопряженности процессов формирования биоморф и видообразования, параллельные ряды биоморф обнаруживаются практически во всех родах, особенно ярко они просматриваются в Hylotelephium и Aizopsis, не проявляющих филогенетического родства (см. гл. 3), сходство биоморф вполне можно объяснить сходством местообитаний, в которых они сформировались.
Следуя концепции Т.И. Серебряковой (1979), можно предположить, что предковыми формами толстянковых были симподиально-полурозеточные (или розеточные) мягкодревесные хамефиты с полициклическими монокарпическими побегами и тирсоидными облиственными соцветиями. Подобные жизненные формы сохранились в родах Aeonium, Sempervivum, Greenovia (Ebel et al., 1991), распространенных главным образом на островах Макаронезии. Однако H. ‘t Hart (1994) предположил, что наиболее вероятной гипотетической предковой формой роста обладает Sedum acre - многолетник с неспециализированными приподнимающимися или распростертыми укореняющимися олиственными гапаксантными цветоносными побегами. Подобный тип довольно обычен среди представителей Sedum s. str. Особенности строения вторичной ксилемы как «полудревесных», так и травянистых Sedoideae (Hart, Koek-Noorman, 1989; см. также гл. 4) показали, что признаки анатомии стеблей свидетельствуют о вторичности одревеснения. Как примитивные, так и эволюционно продвинутые виды имеют идентичное строение вторичной ксилемы, что, по мнению H. 't Hart (1994), подтверждает гипотезу о происхождении Crassulaceae от первично травянистых предков, и то, что одревеснение возникло независимо в нескольких группах как результат приспособления к засушливым условиям.
К подобным выводам привел и филогенетический анализ нуклеотидных последовательностей рибосомной ДНК макаронезийских Sempervivoideae (Mes et al., 1996), показавший, что травянистые Sedum находятся в основании клады. Можно было бы предположить, что базальное положение травянистых форм характерно для семейства в целом, однако наши исследования (Gontcharova, Gontcharov, 2004; см. также гл. 3) продемонстрировали, что травянистые виды занимают позиции в основании Acre- и Aeonium (Макаронезийской)-ветвей филогенетического дерева Crassulaceae, а в Telephium-кладе, включающей, главным образом, восточно-азиатские виды, жизненную форму, близкую к гипотетическому предку sensu H. ‘t Hart (1994), имеет только Phedimus stellatus, помещенный в качестве вершинной ветви Rhodiola-клады, что ставит под сомнение предположение H. ‘t Hart (1994). Подобное положение на древе может указывать как на сохранение предкового состояния признака, так и на вторичное упрощение в результате «отравянения».
Нам представляется вероятным, что преобразования жизненных форм в подсемействе Sedoideae шли от бессезонных мягкодревесных нанофанерофитов или хамефитов через полудревесные биоморфы бессезонного климата, к полудревесным и травянистым сезонного. Общая тенденция биоморфологической эволюции восточно-азиатской ветви Sedoideae заключается, на наш взгляд, в интенсификации процессов жизнедеятельности, всего онтогенеза, отдельных его циклов, которая ведет за собой последующие изменения анатомической структуры, направления роста, появление специальных органов и сокращение длительности жизни надземных осей. Эти тенденции характерны для растений сезонного климата в целом (Cеребряков, 1955, 1962; Скрипчинский, 1977; Хохряков, 1981; Юсуфов, 1996; и др.). Однако мы допускаем возможность не только редукционного (многолетник ® однолетник), но и реверсионного развития (однолетник ® многолетник) в биоморфогенезе Sedoideae.
Усилившееся в конце неогена глобальное похолодание климата (Блютген, 1973; Буданцев, 1983) обусловило, вероятно, формирование сезонных биоморф и определило общую тенденцию к «геофитизации» очитковых, вызвавшую, по-видимому, сначала одревеснение предковых форм, а затем и «отравянение». Возможно, открытые почки возобновления являются своеобразным наследием африканских или средиземноморских предков (плезиоморфным признаком), а происхождение закрытых, характерных для некоторых представителей Rhodiola (см. гл. 4), вероятно, связано с адаптацией к суровым климатическим условиям высокогорий. Специфика коротких вегетационных периодов горных местообитаний выработала у очитковых и целый ряд приспособлений (закладка генеративных органов осенью предыдущего года, весенне-летнезеленый феноритмотип) для оптимального использования благоприятных условий. С коротким вегетационным периодом связано, на наш взгляд, и формирование каудексообразующих гемикриптофитов - современных высокогорных и приполярных видов (Rhodiola rosea). Возможно, что моноподиальное возобновление Rhodiola произошло из симподиального путем торможения развития верхушечной (Böttcher, Jäger 1984), а «каудекс, как орган вегетативного возобновления и запасания питательных веществ, оказался одним из удачных структурных приобретений в процессе исторического развития растений, протекавшего в суровых климатических условиях» (Нухимовский, 1969, c. 77). Корневищные моноподиально нарастающие гемикритпофиты, по-видимому, являются производными каудексообразующих гемимикриптофитов, это подтверждается сходным анатомическим строением (см. гл. 4) и общим типом нарастания.
В условиях похолодания климата выработались приспособления, обеспечивающие успешное возобновление после прохождения неблагоприятных периодов, произошел постепенный переход от хамефитов к гемикриптофитам и криптофитам, постепенное погружение почек возобновления в почву, что обеспечило максимальную защиту органов возобновления от воздействия суровых климатических условий.
Очевидно, что в условиях сезонного климата криптофиты получили преимущественное развитие, так как оказались наиболее приспособленными к переживанию неблагоприятных сезонов и сравнительно короткому вегетационному периоду. Это подтверждается широким распространением и наибольшим числом представителей группы криптофитов среди видов подсемейства, произрастающих в умеренных широтах.
Однако, как уже отмечалось нами, в наиболее суровых условиях высоких арктических широт среди толстянковых преобладают все же не криптофиты, а гемикриптофиты. Вероятно, в Арктике в роли лимитирующего фактора выступают не только низкая температура, но и малая продолжительность вегетационного периода. Гемикриптофиты имеют в этом отношении несомненные преимущества перед геофитами, отрастая до полного оттаивания поверхностного слоя субстрата, а следовательно, более интенсивно используя короткий вегетационный период.
С процессом «геофитизации» связано и основное направление преобразований биоморф восточно-азиатских Sedoideae, шедшее на основе симподиального типа возобновления и приведшее к образованию длиннопобеговых трав. Длиннопобеговые формы роста возникают путем интеркалярного удлинения междоузлий, увеличения их числа при полимеризации (Цвелев, 1977, 1981). Данный процесс, вероятно, был связан с освоением мелкоземистых местообитаний, имевших, по сравнению со скальными, более богатые почвы, большую влажность и меньшую интенсивность инсоляции. Результаты филогенетического анализа (см. гл. 3) позволяют предположить, что длиннопобеговые жизненные формы произошли от полурозеточных. Из топологии построенного нами дерева (см. рис. 9) видно, что именно длиннопобеговость является апоморфией для восточно-азиатских очитковых, а не розеточность и полурозеточность побегов, как это считалось ранее (Ham, Hart, 1998).
Корневищные криптофиты с симподиальным нарастанием, вполне вероятно, могли произойти от полукустарничков (или травянистых хамефитов). На склонах или осыпях многолетние части побегов полукустарничков часто погребаются подвижным субстратом, они способны укореняться в каждом узле побега, образуя эпигеогенные корневища.
Возникновение клубнеобразующих жизненных форм также связано с ростом в засушливых условиях (Clausen 1977; Hart, 1991), необходимостью запасания воды и питательных веществ для успешного переживания неблагоприятных сезонов.
Аридизация климата повлекла за собой общее ускорение развития растений и следующее за ним сокращение жизненного цикла. Она также вызвала преобладающее развитие генеративных органов, усиление семенного размножения, по сравнению с вегетативным, ослабила вегетативное возобновление (Серебряков, 1955). В результате ксерогенеза при сокращении длительности жизни скелетных осей сформировались малолетние и однолетние (терофитные) жизненные формы толстянковых - суккулентнолистовые однолетние травы. Как уже упоминалось, область распространения видов данного типа образует южную границу ареала подсемейства, практически совпадающую с границами между семиаридными и аридными областями Северного полушария. Однолетние жизненные формы рассматривались H. ‘t Hart (1994) как неотенические - максимально приспособленные к аридным условиям.
Диаметрально противоположную гипотезу выдвинул G. Knaben (1966), считавший, что в роде Sedum s.l. эволюционные преобразования шли от однолетних или двулетних форм к многолетним. Подобные выводы автор сделал на основании изучения морфогенеза травянистых хамефитов и одно-, двулетних форм. Он полагал, что у малолетних форм происходит подавление семенного размножения, замена его вегетативным приводит к образованию многолетников, размножающихся практически исключительно вегетативно. Такой реверсивный вариант сценария также кажется нам вполне вероятным, тем более что косвенным подтверждением может служить редукция числа цветков в соцветии и очень редкое цветение рассматриваемых G. Knaben форм.
В основе эволюции биоморф лежат преобразования онтогенеза в целом и его отдельных фаз. Например, при переходе от кустарников к травам идет сокращение ювенильно-виргинильного периода (Скрипчинский, 1977).
Какие же процессы могли привести к преобразованиям, вызвавшим существующее разнообразие биоморф очитковых?
Основные модусы преобразования онтогенеза растений - пролонгация, аббревиация и девиация, а также ретенции, рекапитуляции и инверсии (Тахтаджян, 1954, 1964; Серебрякова, 1983; Недолужко, 1997; и др.). В зависимости от взглядов авторов на основное направление эволюции биоморф - от древесных к травянистым или наоборот, одни и те же факты считаются результатом различных процессов. Так, А.Л. Тахтаджян (1954, 1964) считает, что пролонгации имеют минимальное значение в эволюции растений, а Т.И. Серебрякова (1983), наоборот, что они имеют существенное значение, например, при переходе с системы главного корня целиком на придаточную. А.Л. Тахтаджян считает (1954, 1964) одревеснение нижних частей травянистых растений явлением ретенции (сохранения палеоморфной структуры некоторых частей взрослых растений), а В.А. Недолужко (1997) считал такое одревеснение, не относящимся к онтогенезу вообще, а только сезонным явлением.
Мы полагаем, что в Sedoideae преобразование полукустарничковых биоморф в корневищные, образование «наземных» корневищ и запасающих органов, преобразование биоморф от вегетативно неподвижных к вегетативно подвижным являются результатом терминальных пролонгаций (анаболий). Образование монокарпических форм из поликарпических обычно связывается с неотенией - сочетанием девиации с терминальной аббревиацией (Тахтаджян, 1954, 1964), выражающейся у Sedoideae в акселерации онтогенеза (Юсуфов, 1979). При акселерации происходит перенос закладки генеративных органов на более ранние стадии онтогенеза и аббревиация конечных стадий. Подобный перенос уменьшает зависимость процессов воспроизведения потомства от внешних условий, растения оказываются способными вступать в генеративную фазу при наличии минимальных условий (Юсуфов, 1996). С акселерацией у Sedoideae, вероятно, может быть связано и преобразование полудревесных биоморф в травянистые и дициклических побегов в моноциклические. А.Г. Юсуфов (1979) также считает, что в ходе эволюции семейства толстянковых наблюдается акселерация онтогенеза посредством сочетания базальной девиации с медиальной аббревиацией, выражающаяся в отсутствии покоя семян и проростков, живорождении, сокращении периода смены метамеров. Акселерация онтогенеза выражается также и в упрощении структуры надземного побега, сопровождающемся сокращением жизни монокарпических побегов, усилением «геофитизации».
Оригинальный рисунок А.Р. De Candolle www.illustratedgarden.org/mobot/rarebooks, Rhodiola pinnatifida nature.chita.ru.
Читать далее: заключение и список литературы
