|
Подсемейство Sedoideae (Crassulaceae) флоры Сибири и российского Дальнего Востока (систематика, биоморфология, филогения)
С. Б. Гончарова
Глава 1. Очерк истории изучения сем. Crassulaceae и подсемейства Sedoideae
1.1. История изучения сем. Crassulaceae
Название семейства толстянковых произошло от латинского crassus, что означает толстый, и полностью отражает характерный облик его типичных представителей. Первое известное изображение растения из семейства толстянковых (Kalanchoe citrine Schweinfurth) было обнаружено на одной из стен Большого Храма фараона Тутмоса III (1501-1447 гг. до н.э.) в Египте (Rowley, 1997). Упоминания о растениях Sedum встречаются еще в трудах Плиния (Regnat, 2004), и именно он считается автором имени рода Sedum. Значение его до сих пор является объектом споров, считается, что название произошло либо от sedere (сидеть), либо от sedare (успокаивать, спокойный, тихий) (Краснов и др., 1979). Приверженцы первого варианта делают упор на внешний облик представителей рода, второго – апеллируют к Плинию, применявшему термин sedare к группе растений с успокаивающим действием, в том числе Sedum, Aizoon и Telephium (Regnat, 2004). Автором эпитета Aizoon считают Демокрита (Regnat, 2004), название произошло из греческого языка (aeizvou) и использовалось для родов Sedum и Sempervivum L., оно имеет сходное значение с латинским sempervivum – вечноживущий, вечный. При описании рода Hylotelephium H. Ohba (1978) указывает, что название происходит от сочетания слов ulh (лесной) и Thlejion (суккулентное растение), однако мы склонны согласиться с мнением L.R. Praeger (1921), что растения названы по имени Телефуса, сына Геракла, излечившего раны соком этого растения. Названия других дальневосточных родов более однозначны, Rhodiola произошло от rhodiae – уменьшительного роза из-за специфического, похожего на розу, запаха корня, а Orostachys образовано из двух корней, означающих гора и колос, по особенностям мест обитания и колосовидному соцветию.
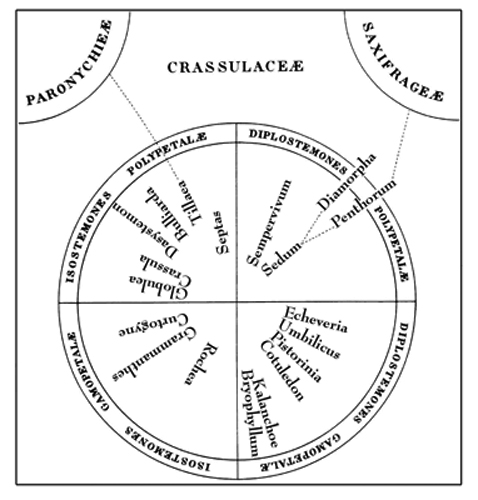
Рис. 1. Схема филогенетических взаимоотношении в сем. Crassulaceae A.P. de Candolle (1828).
|
Формально семейство Crassulaceae описано A.Р. de Candolle в 1805 г. (Lamarck, Candolle, 1805), хотя первая его система была опубликована четырьмя годами ранее (Candolle, 1801). Все последующие исследования подтвердили, что Crassulaceae естественная группа. Спорным оставалось и остается до сих пор лишь положение рода Penthorum Cronov, который считали членом Crassulaceae, Saxifragaceae DC. или олиготипного Penthoraceae Tieghem (Candolle, 1828 a, b; Tieghem, 1898; Engler, 1930). Концепция семейства, предложенная A.Р. de Candolle (1801, 1828 a, b), послужила основой для построений всех последующих исследователей.
В конце XIX–начале ХХ столетия было предложено несколько систем семейства Crassulaceae. Из них наибольшее значение имели классификации A.P. de Candolle (1828 a, b) (рис. 1), S. Schönland (1891), А. Berger (1930), H. Frödersröm (1930-1935).
В первой трактовке A.Р. de Candolle (1801) семейство было подразделено на две группы: «gamopetalae» и «polypetalae» – в соответствии с наличием или отсутствием спайнолепестности. В первую группу вошли роды Cotyledon L., Umbilicus DC. и Kalanchoe Adanson; во вторую – Bulliarda DC., Tillaea L., Crassula L., Sedum и Sempervivum. В «Prodromus Regni Vegetabilis» этот автор (Candolle, 1828а) приводит уже 19 родов с 308 видами в семействе. Автор разделил семейство на две трибы: Crassuleae Legitimae (305 видов из 17 родов) со свободными плодолистиками Crassuleae Anomalae, включив в него два олиготипных рода [Diamorpha Nuttall (1) и Penthorum (2)] со сросшимися плодолистиками. В основу классификации «настоящих» Crassulaceae положены два основных признака: число тычинок и спайнолепестность.
Внутри трибы A.P. de Candolle (1928 a, b) выделил две подтрибы: Isostemonae и Diplostemonae –, которые в свою очередь разделил на две неформальные группы: Gamopetalae (спайнолепестные) и Polypetalae (свободнолепестные).
|
В обработке семейства S. Schönland (1891) для первого издания «Die natürlichen Pflanzenfamilien» число Crassulaceae, известных науке, возросло до 469. Автор не выделял какие-либо группы внутри семейства, что, по мнению U. Eggli (1988), было «шагом назад» по сравнению с обработкой A.Р. de Candolle. Число родов было сокращено до 13, 5 из которых – монотипные. В обработке в виде диаграммы была представлена система семейства, в центре которой был помещен род Sedum. Как подчеркивает H. 't Hart (1995), эволюционные схемы S. Schönland поразительно совпадают с современными филогенетическими построениями, сделанными на основе генетического анализа. S. Schönland (1891) предположил, что род Sedum является предковым для семейства (рис. 2).
|
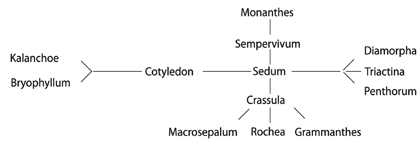
Рис. 2. Схема филогенетических взаимоотношений в сем. Crassulaceae S. Schönland (1891).
|
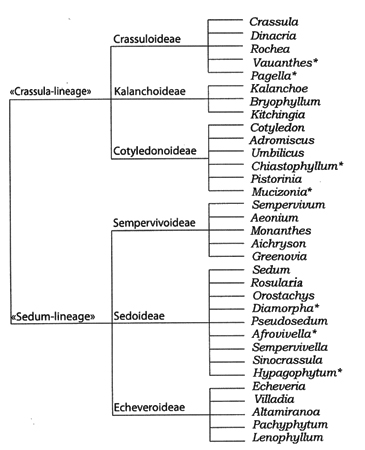
Рис. 3. Схема филогенетических взаимоотношении в сем. Crassulaceae A. Berger (по: Hart, 1995). Звездочкой обозначены монотипные роды.
|
Со времен обработки Crassulaceae, сделанной A. Berger (1930) для второго издания «Die natürlichen Pflanzenfamilien», классификация семейства в основном не изменялась. По сравнению с предыдущими, система была значительно усовершенствована, стала более логичной и простой в использовании, но, тем не менее, оставалась искусственной. Обработка A. Berger и до сих пор является наиболее авторитетной на внутрисемейственном и родовом уровне (Hart, Eggli, 1995; Ham, 1995). В соответствии со взглядами A. Berger семейство содержит 6 подсемейств, 33 рода, около 1400 видов (рис. 3). Подсемейства сгруппированы по три в две группы: Crassula-линия (Crassuloideae, Kalanchoideae, Cotyledonoideae), которая включала таксоны из Южного полушария, и Sedum-линия (Sempervivoideae, Sedoideae, Echeveroideae) – из Северного. Каждое подсемейство содержало один из крупных родов (Crassula, Kalanchoe, Cotyledon, Sempervivum, Sedum и Echeveria DC.), по имени которого и было названо. Внутри названных линий для разграничения подсемейств наибольшее значение придавалось признакам числа и расположения частей цветка, степени сростнолепестности и листорасположения. Например, Crassuloideae включало виды с гаплостемонным андроцеем и несросшимися лепестками чашечки, Kalanchoideae – с четырехчленными цветками, диплостемонным андроцеем и сросшимися лепестками. Внутри Sedum-линии Sempervivoideae характеризовалось полимерными цветками со свободными лепестками и розеточным габитусом, Echeveroideae – пятимерными цветками с частично или полностью сросшимися лепестками (Eggli, 1988; Mort et al., 2001). Sedoideae же, по меткому замечанию С.H. Uhl (1963), явилось «catch-all» таксоном, в который были собраны все роды, не укладывающиеся в приведенные выше характеристики, а в род Sedum – все виды, не укладывающиеся в диагнозы других родов, это и определило все последующие проблемы как подсемейства, так и рода.
Построения H. Fröderström (1930-1935) имели ярко выраженный географический подход. Во введении к монографии рода Sedum H. Fröderström развил совершенно новую революционную теорию о происхождении и эволюции Crassulaceae. Он не принял точку зрения S. Schönland о центральном положении Sedum в филогении семейства и предположил, что предком семейства является космополитный род Tillаea. H. Fröderström полагал, что Sedum – полифилитический таксон, произошедший от двух независимых линий. Эта идея не была признана современными исследователями.
|
Наиболее полная обработка толстянковых флоры CCСР была проведена А.Г. Борисовой (1939, 1969). Основное отличие данной системы от таковой A. Berger (1930) в том, что некоторые секции переведены в ранг родов и разделены на более мелкие таксоны. Для флоры страны А.Г. Борисовой были приведены 129 видов из 10 родов и 5 подсемейств (включая Penthoroideae с двумя видами), описано 28 новых видов.
|
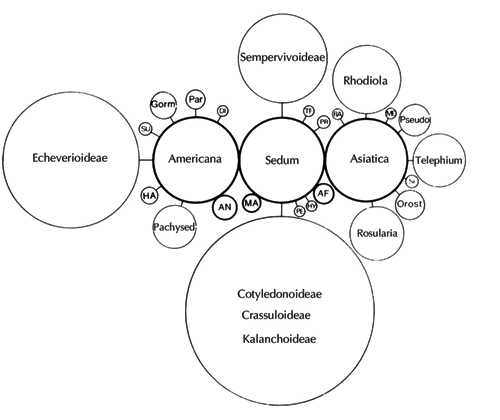
Рис. 4. Схема филогенетических взаимоотношений в сем. Crassulaceae H. ‘t Hart (1982).
Размер кругов пропорционален числу видов в таксоне. SU – Sulcus, Gorm – Gormania, Par – Parvisedum, DI – Diamorpha, HA – Hasseanthus, Pachysed – Pachysedum, AN – Sedum sect. Andina, MA – S. sect. Macaronesia, PE – Perrierosedum, HY – Hypagophytum, TE – Telmissa, PR – Prometheum, BA – Balfouria, ME – Meterostachys, Pseudo – Pseudosedum, SI – Sinocrassula, Orost – Orostachys.
Первой попыткой построения филогенетической классификации Crassulaceae была работа H. 't Hart (1982) (рис. 4), система основывалась главным образом на географическом распространении представителей семейства. Автор сделал вывод, что кроме космополитных Sedum и Tillea, встречающихся на всех трех континентах Северного полушария, все остальные роды приурочены к отдельным континентам или флористическим районам. Используя биогеогафический подход, H. 't Hart, подобно A. Berger, разделил семейство на две ветви: южную, включающую подсемейства Crassuloideae, Cotyledonoideae, Kalanchoideae, и северную, включающую Sempervivoideae, Sedoideae, Echeveroideae. Три самых больших секции Sedum занимают центральное положение в эволюционной схеме рода, каждая секция является эндемичной для региона либо субконтинента. По мнению H. 't Hart, его построения демонстрируют естественность и монофилитичность секций (Hart, Eggli, 1995).
|
Попытка пересмотреть структуру семейства была предпринята А.Л. Тахтаджяном, в «Системе магнолиофитов» (Тахтаджян, 1987) оно разделено на 4 подсемейства: Sedoideae (включающее Sempervivoideae), Kalanchoideae (включающее Cotyledonoideae), Echeveroideae и Crassuloideae, а позже (Takhtajan, 1997) – на три (Crassuloideae, Kalanchoideae и Sedoideae). R.F. Thorn (1992, 2000) также сначала сократил число подсемейств до трех: Sedoideae (включающее Sempervivoideae и Echeveroideae), Cotyledonoideae (включающее Kalanchoideae) и Crassuloideae (Thorn, 1992), а затем до двух: Crassuloideae и Sempervivoideae (по праву приоритета над Sedoideae) (Thorn, 2000), однако эти построения не были признаны.
В 1995 г. в результате работы XXII и XXIII конгрессов «International Organization for Succulent Plant Study» и симпозиума «Evolution and Systematics of the Crassulaceae» была предпринята попытка достигнуть консенсуса в классификации Crassulaceae (Eggli, et al., 1995). В «Consensus Classification» представлены все принимаемые роды, авторы отметили, что, хотя «Consensus…» одобрен не всеми участниками конгресса, он принят в качестве «международного стандарта» для таксономии и номенклатуры семейства. Согласно «Consensus…», Crassulaceae насчитывает 33 рода и около 1300 видов. Хотя количество родов и видов примерно соответствует таковым в системе A. Berger, родовой состав семейства значительно изменен, к примеру, самостоятельными считаются роды: Rhodiola, Hylotelephium, Meterostachys Nakai, Perrierosedum (A. Berger) H. Ohba. Такой «демократический» подход, на наш взгляд, отражает тупиковую ситуацию, сложившуюся в систематике семейства.
С работ R.C.H.J. van Ham и H. 't Hart началась эпоха молекулярно-филогенетического подхода в таксономии Crassulaceae.
R.C.H.J. van Ham (1995) на основании результатов рестриктного анализа ДНК хлоропластов 44 видов из 19 родов выделил 7 клад в составе семейства (рис. 5).
1. Сrassula-клада с высокой поддержкой включает только членов подсем. Crassuloideae в понимании A. Berger. Как отмечено выше, подсемейство характеризуется гаплостемонными цветками, что очень редко среди членов других 6 клад. Ветвь включает около 300 видов, распространенных главным образом в Южной Африке. Базальная дихотомия между Сrassula-кладой и остальной частью семейства отличается наибольшей дивергенцией последовательностей (около 7,3%). 2. Устойчивая Kalanchoe-клада, представленная примерно 250 видами из Kalanchoideae и южно-африканскими родами Cotyledonoideae в понимании A. Berger. Расположение родов Cotyledonoideae в двух разных кладах свидетельствует о том, что подсемейство парафилитично. 3. Telephium-клада включает род Umbilicus подсем. Cotyledonoideae и роды Rhodiola, Hylotelephium и некоторых представителей Sedum s. l. (Phedimus Rafinesque, Aizopsis) подсем. Sedoideae. Группа весьма таксономически разнородна и имеет низкую статистическую поддержку, она объединяет виды из Северного полушария, характеризующиеся «плоскими листьями с зубчатым краем». 4. Sempervivum-клада включает одноименный род из Sempervivoideae в понимании A. Berger и некоторых представителей Sedum s. str. 5. Leucosedum-клада содержит большую группу Sedum s.str., некоторых представителей полифилитичного Rosularia (DC.) Stapf и два средиземноморских рода из подсем. Cotyledonoideae – Mucizonia (DC.) Berger и Pistorinia DC. 6. Aeonium-клада, включающая макаронезийские Sempervivoideae и Sedum s. str. 7. Наиболее крупная и устойчивая Acre-клада объединяет американское Echeverioideae и Sedum s. str. Однако сам автор отмечал, что взаимоотношения в группе, включающей клады с 4 по 7, все же остаются нерешенными, и не исключал влияния «эффекта притяжения длинных ветвей» на подобное положение. Основной вывод, сделанный R.C.H.J. van Ham, говорит о том, что, по крайней мере, 4 из 6 подсемейств в понимании A. Berger – искусственные группы.
|
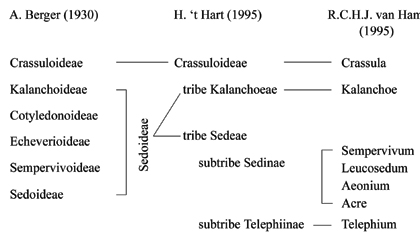
Рис. 5. Внутрисемейственные классификации Crassulaceae A. Berger, H. ‘t Hart и R.C.H.J. van Ham (по: Mayzumi, Ohba, 2004, с изменениями).
|
Другая внутрисемейственная классификация семейства предложена H. 't Hart (1995) [табл. 1], она явилась результатом филогенетического анализа ДНК хлоропластов 44 видов из 19 родов и trnL-F межгенного спейсера 49 видов из 26 родов. Автор сократил число подсемейств до двух: Crassuloideae (250 видов из двух родов) и Sedoideae (более 1100 видов из 28 родов). Crassuloideae в понимании H. 't Hart соответствует традиционному, а Sedoideae включает оставшиеся пять подсемейств в понимании A. Berger. H. 't Hart отмечает, что полученные результаты в целом соответствуют результатам R.C.H.J. van Ham. В обеих филогениях четко выражено базальное разделение между преимущественно африканским Crassuloideae, характеризующимся, как отмечено выше, гаплостемонным андроцеем и супротивными листьями, и остальными Crassulaceae, не имеющими единой уникальной комбинации признаков. Sedoideae, в свою очередь, разделено на две трибы: африканскую Kalanchoeae и Sedeae, приуроченную к Северному полушарию. Kalanchoeae в целом соответствует Kalanchoe-кладе, выделенной R.C.H.J. van Ham (в нее вошли Kalanchoideae в понимании A. Berger и большая часть Cotyledonoideae). H. 't Hart отмечает, что высокая степень поддержки и уникальный набор фенотипических признаков подтверждают естественность трибы Kalanchoeae. Триба Sedeae содержит подтрибы: Telephiinae и Sedinae – в первую входят главным образом восточно-азиатские роды, во вторую – роды, распространенные в Северном полушарии. Последняя подтриба включает в себя около половины всех родов и видов Crassulaceae и, как и в системе A. Berger, является сборным таксоном, в который вошли роды, не укладывающиеся в диагнозы других внутрисемейственных таксонов. Попытки H. 't Hart определить уникальные для подтрибы признаки не увенчались успехом. Хотя, по мнению автора, подтриба Telephiinae четко характеризуется набором уникальных признаков (пятичленными цветками с обдиплостемонным андроцеем и свободными лепестками и т.д.), она является слабо поддержанной.
Таблица 1
Система Crassulaceae по: H. 't Hart (1995)
Таксон |
Число
видов |
Число
родов |
Распространение |
subfam. Crassuloideae |
250 |
2 |
Южная Африка |
subfam. Sedoideae |
|
|
|
tribus Kalanchoeae
|
250 |
5 |
Южная Африка |
tribus Sedeae
|
|
|
|
subtribus Telephiinae
|
150 |
8 |
Азия |
subtribus Sedinae
|
700 |
18 |
Северное полушарие |
Наиболее современной обработкой семейства в мировом масштабе можно назвать «Illustrated Handbook of Succulent Plants: Crassulaceae» (2005) под редакцией U. Eggli. В соответствии с обработкой семейство содержит 1410 видов и 305 подвидов из 33 родов и 23 нотородов. Например, самый крупный род Sedum представлен 428 видами, второй по величине Crassula – 195 видами. В книге авторы придерживаются «Consensus Classification» (Eggli, et al., 1995) с некоторыми изменениями, и концепции семейства H. 't Hart (1995). Однако и эта обработка содержит большое число неточностей, особенно это касается видов Cеверо-Восточной Азии, с большинством из которых авторы знакомы лишь по гербарным образцам или литературным источникам.
Crassulaceae всегда и бесспорно считалось членом либо порядка Saxifragales Dumort. (Тахтаджян, 1966; Hutchinson, 1969; Thorn, 2000), либо неформальной группы Saxifragineae (Cronquist, 1981). Таксон помещался либо в подклассе Rosidae порядке Rosales Perleb, в непосредственном соседстве с семействами Alseuosmiaceae Airy Shaw и Cephalotaceae Dumort. (Cronquist, 1981), либо в подклассе Rosidae, надпорядке Rosanae (Тахтаджян, 1966, 1987; Thorn, 2000). Ближайшими родственными семействами всегда считались Cephalotaceae и Saxifragaceae, тесно связанное с Crassulaceae переходным родом Penthorum, сочетающим в себе признаки обоих семейств. Однако результаты исследований Angiosperm Phylogeny Group (2003) указали на необходимость выделения Saxifragales Lindley из Rosidae и поместили в неформальную группу «Core Eudicots», в качестве сестринской ветви для прочих Rosid. Авторы отмечают, что состав порядка Saxifragales явился одним из «сюрпризов» молекулярно филогенетического анализа покрытосеменных. Анализ указал на сестринские отношения Crassulaceae с Haloragaceae R. Br., Tetracarpaceae Nakai и Penthoraceae.
1.2. История изучения подсемейства Sedoideae и отдельных восточно-азиатских родов
История развития таксономии Sedoideae по сути своей сводится к истории таксономии его центрального рода Sedum s. l., все известные ныне роды подсемейства когда-либо были отнесены к Sedum как подроды, секции или серии. До настоящего времени сохранились два основных взгляда на объем подсемейства (Ohba, 1978). Согласно одному из них Sedoideae делится на довольно большое число родов, и Sedum понимается в «узком» смысле, как, например, в системе А. Berger (1930), различавшего в пределах подсемейства 9 родов. Согласно второму подсемейство включает один крупный род Sedum s. l. (Candolle, 1828а; Praeger, 1921 и др.).
В работах С. Linnaei (1753, 1840) роды разделялись по числу тычинок и плодолистиков, Sedum характеризовался 5-мерными диплостемонными (Decandria Pentagyna) цветками и был разделен на две группы в соответствии с морфологией листьев, «Planifolia» (плосколистные) и «Teretifolia» (вальковатолистные). Род Rhodiola был отделен от Sedum на основании двудомности растений и был отнесен в группу Dioecia Octandria, а единственный известный С. Linnaei (1753) вид горноколосника (Orostachys spinosa (L.) C. A. Mey.) сначала был отнесен к роду Cotyledon в группе Decandria Pentagyna, а затем к роду Crassula в группе Pentandria Pentagyna (Linnaei, 1840). В первом издании «Species Plantarum» С. Linnaei (1753) приводит 15 видов в роде Sedum, один вид – в роде Rhodiola, а в «Systema, genera, species plantarum» (Linnaei, 1840) – уже 19 в Sedum и один в Rhodiola. Интересно, что, хотя роды и помещены в разные группы, расположенные в разных частях системы, С. Linnaei (1840) указывает на их родство.
В 1808 г. F. Fischer в «Catalogus Horti Gorenkensis» представил род Orostachys, однако в работе не был приведен диагноз, и название не считалось валидно опубликованным, что повлекло за собой почти полуторавековую путаницу в определении авторства рода (Byalt, Sokolova, 1999). Почти до конца ХХ века автором рода считался A.P. de Candolle или A. Berger. Авторство F. Fischer было восстановлено С.К. Черепановым (1973), а затем В.В. Бялтом и И.В. Соколовой (Byalt, Sokolova, 1999). Спустя год после первой публикации F. Fischer дал подробное описание рода (1809). Изучив строение цветка, автор сделал вывод о родстве Orostachys и Sedum (из родства S. telephium). В новый род F. Fischer включил 4 вида (в том числе ошибочно – Rosularia serrata (L.) Berger).
В уже упомянутой нами выше работе A.P. de Candolle (1828a) признавал 88 видов в роде Sedum s.l. Род Rhodiola был помещен им в неформальной группе «Planifolia, floribus luteus» вместе с такими видами, как Sedum aizoon L. и S. hybridа L., а Orostachys автор распознавал в качестве секции рода Umbilicus в подсемействе Cotyledonoideae.
Своеобразие морфологии Umbilicus livenii Ledeb. позволило E. Boissier (1872) выделить этот вид в секцию Pseudosedum, а позже А. Berger (1930) придал секции ранг рода. Основным диагностическим признаком Pseudosedum А. Berger (1930) считал спайнолепестный венчик.
В 1921 г. R.L. Praeger в таксономическом обзоре рода Sedum, посвященном главным образом культивируемым видам, различал уже 10 секций, принимая среди них Rhodiola, Telephium и Aizoon.
Настоящим прорывом в систематике как Crassulaceae вообще, так и Sedoideae в частности была работа А. Berger (1930). Однако подсемейство Sedoideae не было им четко очерчено и характерные признаки не выяснены, так что разделить подсемейства Sedoideae, Sempervivoideae, Cotyledonoideae и Echeverioideae было весьма затруднительно (Eggli, 1988). Взяв за основу идеи R.L. Praeger (1918, 1919, 1921), А. Berger значительно развил систему, он принимал в качестве самостоятельных 9 родов в подсемействе (Sedum, Sinocrassula Berger, Orostachys, Rosularia, Sempervivella Stapf, Afrovivella Berger, Pseudosedum, Diamorpha, Hypagophytum Berger). Самый крупный род Sedum включал в себя около 500 видов и разделен на 22 секции, которые в свою очередь были разделены на значительное число серий, к примеру, sect. Sedum содержала 27 серий.
Первая наиболее полная и обстоятельная система рода Sedum s.l. была опубликована H. Fröderström (1930-1935). Основываясь на признаках формы завязи и географическом распространении, он объединил выделяемые А. Berger роды, за исключением Diamorpha и Hypagophytum, в один более крупный Sedum s.l. и разделил его на 9 секций и 51 серию. Автор различал 4 группы, которые были названы Africana, Americana, Asiatica и Eurasiatica, три последние, в свою очередь, разделены в соответствии с морфологией плодолистиков (orthocarpia и kyphocarpia). Rhodiola и Telephium приняты в качестве секций, Orostachys – в качестве группы в sect. «Asiatica Genuina Orthocarpia», а Aizoon – группы в sect. «Asiatica Genuina Kyphocarpia». Концепция H. Fröderström никогда не была принятой и считалась большинством последователей искусственной (Ohba, 1978; Hart, 1982).
Значительный вклад в таксономию Sedoideae был внесен Т. Nakai (1938). Начиная с работ этого автора, Orostachys и Rhodiola окончательно приняты в ранге родов. Т. Nakai впервые выделил из Sedum роды Chamaerhodiola Nakai и Meterostachys Nakai. В 1938 г. Т. Nakai, изучая японских представителей Rhodiola, впервые со времен C. Linnaei четко обозначил род и указал на аксилярную природу генеративных побегов и чешуевидных корневищных листьев, отличающих Rhodiola от других представителей подсемейства.
Важным шагом в упорядочивании внутриродовой классификации Sedoideae была упомянутая выше обработка семейства Crassulaceae А.Г. Борисовой для «Флоры СССР» (1939, 1969). Секции и ряды, представленные данным автором, были основаны главным образом на признаках вегетативной сферы. Справедливость основных построений А.Г. Борисовой, естественность большинства внутриродовых таксонов подтверждают данные хемотаксономических исследований (Гнедков, Шретер, 1977; Шнякина, 1979; Куркин и др., 1986). Taк, П.Н. Гнедковым и А.И. Шретером (1977) установлено, что состав флавоноидных соединений является видоспецифичным и соответствует таксономическому положению секций. Г.П. Шнякиной (1979) при сравнении химического состава 16 видов дальневосточных очитков также отмечено значительное сходство состава химических соединений внутри секций и столь же значительные межсекционные различия.
Значительный вклад был внесен А.Г. Борисовой и в разработку внутриродовой структуры рода Pseudosedum (Борисова, 1933, 1939, 1969; Паланов, 1988). Автором были описаны 3 секции и 10 видов рода. Однако, как указывал А.В. Паланов (1988), А.Г. Борисова не всегда правильно оценивает таксономическое значение признаков строения вегетативных органов, вследствие чего система оказалась искусственно усложненной. Существенные изменения в систему рода были внесены А.В. Палановым (1988), он также впервые указал на наличие «кармановидных» нектарников в основании завязей, что, несомненно, выделяет Pseudosedum из других Sedoideae.
Классификация европейских, северо-африканских и западно-азиатских Sedoideae была предложена V. Grulich (1984). Автор сделал попытку разделить Sedum s. l. на естественные группы и выделил роды: Aizopsis (7 видов), Asterosedum (5), Petrosedum (3), Oreosedum (30). Разделение родов в данной классификации основано главным образом на географическом распространении видов, признаках вегетативной сферы (форме роста, расположении листьев), морфологии соцветий и строении гинецея.
Значительные изменения не только в систему семейства, но и подсемейства внес H. 't Hart. Так, внутри Sedoideae им (Hart, 1995) выделены две трибы: Kalanchoeae (250 видов, 8 родов) и Sedeae. Последняя, в свою очередь, разделена на две подтрибы: Telephiinae (150 видов, 5 родов) и Sedinae (700 видов, 18 родов). Более половины видов Sedinae содержатся в наибольшем по числу видов роде Sedum s.str. (350). Несмотря на его бесспорную полифилитичность, автор сохраняет род Sedum для стабильности номенклатуры. Другие роды варьируют по числу видов от очень больших (Echeveria; 150-200 видов), средних (Aeonium Webb et Berthelot, Dudlea Britton et Rose, Sempervivum; 50-40) и маленьких до монотипных (менее 15 видов). Подтриба Telephiinae равна Telephium-кладе и включает в себя Hylotelephium, Rhodiola, Sedum subgen. Aizoon (Koch ex Schönland) H. Ohba, S. subgen. Spathulatae (A. Boriss.) H. Ohba, Orostachys. Виды этой группы распространены главным образом в Азии и названы H. 't Hart плосколистными азиатскими Sedum. Однако Telephium-клада имеет только 25% бутстреп-поддержку и также является полифилитичной (Mayzumi, Ohba, 2004).
На основании обстоятельного изучения морфологии и географического распространения представителей Orostachys В.В. Бялтом (1999а, 1999б) проведена ревизия видового состава рода мировой флоры. Автор установил, что род включает в себя 22 вида, 1 подвид и 5 разновидностей в двух подродах (Orostachys и Schoenlandia H. Ohba), трех секциях и двух рядах. Однако, на наш взгляд, В.В. Бялт следовал слишком узкой концепции вида, по данным других авторов род насчитывает 10-15 видов (Eggli et al., 1995; Mayzumi, Ohba, 2004).
Значительный вклад в изучение Sedoideae Старого Света и Восточной Азии, в частности, внесен японским ботаником Н. Ohba (1977, 1978, 1995, 2001). Главные признаки, взятые в основу классификации Sedoideae, – форма оснований завязей и зрелых листовок, цвет и степень срастания лепестков, наличие или отсутствие розеток. Автор провел ревизию подсемейства, основываясь на азиатских представителях, пересмотрел разделение таксонов внутри Sedoideae. По мнению Н. Ohba (1978, 1995), в Восточной Азии подсемейство насчитывает около 300 видов из 11 родов. При этом род Sedum толкуется Н. Ohba в узком смысле, а Rhodiola и Orostachys приняты в качестве самостоятельных. Автор значительно сократил число секций в роде Sedum и укрупнил некоторые из них; секции Aizoon и Telmissa приняты за подроды, секция Telephium – за род Hylotelephium, также выделены роды Prometheum, Perrierosedum, предложено выделение рода Balfouria. Автор считает, что Восточная Азия является как центром видового разнообразия, так и центром распространения Sedoideae s. str. H. Ohba (1975, 1978, 1980, 1981, 1982) выполнена и наиболее полная обработка рода Rhodiola. На основании изучения большой коллекции гималайских, тибетских и китайских представителей Rhodiola им проведена ревизия существующих видов, описан ряд новых, пересмотрена внутриродовая таксономия. По мнению H. Ohba, род включает в себя 4 подрода (Rhodiola, Primuloides, Crassipedes и Clementsia), 7 секций и около 60 видов.
Попытка оценить филогенетическое положение восточно-азиатских Sedoideae на основе анализа последовательностей ядерной и хлоропластной ДНК 74 таксонов подсемейства была предпринята S. Mayuzumi и H. Ohba (2004). Авторы отмечали, что все клады, выделяемые ранее R.C.H.J. van Ham и H. 't Hart, за исключением Telephium и Leucosedum, имели высокую статистическую поддержку. Полученные S. Mayuzumi и H. Ohba результаты в целом соответствуют таковым предшествующих исследователей, за исключением взгляда на Telephium-кладу, которая, по их мнению, не является монофилитичной. Все восточно-азиатские Sedum s. str. входят в выделенную R.C.H.J. van Ham Acre-кладу, все прочие роды – в Telephium-кладу. По результатам S. Mayuzumi и H. Ohba, Telephium-клада распадается на 4 подклады, названные в соответствии с основными родами в их составе, – Rhodiola, Hylotelephium, Phedimus и Umbilicus. Важнейший результат исследований S. Mayuzumi и H. Ohba, на наш взгляд, – доказательство искусственности большинства внутриродовых таксонов (секций и серий). Авторы подтвердили самостоятельность родов толстянковых, выделенных из состава Sedum s.l.: Rhodiola, Hylotelephium, Phedimus и Umbilicus – однако не смогли установить монофилетичность Telephium-клады, объединяющей роды предположительно восточно-азиатского происхождения. Было отмечено, что естественность этой группы также трудно обосновать и с точки зрения морфологии, поскольку отсутствуют уникальные (синапоморфные) признаки, отличающие Telephium-кладу от остальных Sedeae sensu Hart (Sedoideae). Кроме этого, S. Mayuzumi и H. Ohba (2004) было установлено, что основные роды, слагающие эту кладу, не являются монофилетическими. Так, Hylotelephium и часть представителей Orostachys, а также Rhodiola и Pseudosedum формируют высокоустойчивые клады на филогенетическом дереве. Результаты молекулярно-филогенетических анализов ставят под сомнение естественность этих таксонов и указывают на необходимость пересмотра родовых границ восточно-азиатских Crassulaceae.
Оригинальный рисунок А.Р. De Candolle www.illustratedgarden.org/mobot/rarebooks, Rhodiola pinnatifida nature.chita.ru.
Читать далее: главу 2
|