Подсемейство Sedoideae (Crassulaceae) флоры Сибири и российского Дальнего Востока (систематика, биоморфология, филогения)
С. Б. Гончарова
Глава 6. Географическое распространение Sedoideae
6.1. Географическое распространение и основные экологические особенности очитковых
Представители сем. Crassulaceae широко, но неравномерно распространены по всему земному шару главным образом в субтропических и умеренных областях, с четко выраженной сменой сухого и влажного сезонов. Наиболее обильны толстянковые в Южной Африке, на Мадагаскаре, Канарских островах, Азии, Мексике и Средиземноморье, но редки в Австралии (один род), Океании (два рода) и Южной Америке (Spongberg, 1978). Необходимо отметить, что подсемейства толстянковых имеют достаточно четкую приуроченность в преимущественном распространении к определенным континентам, например, представители Crassuloideae наиболее обычны в Южной Африке, Echeverioideae – Северной Америке, Sedoideae – Евразии.
Существует две гипотезы обоснования такой специфики географии подсемейств: 1 – возникнув в Южной Африке, предки Sedoideae sensu Hart мигрировали в Средиземноморье и уже из этого региона распространились далее по всему Северному полушарию; 2 – возникнув в Средиземноморском регионе, предки Сrassula- и Kalanchoe-клад мигрировали в Африку, а прочие Sedoideae sensu Hart также распространились по Северному полушарию (Ham, 1995; Ham, Hart, 1998; Mort et al., 2001). В обоих случаях могли сформироваться явно выраженные центры диверсификации – Макаронезийский (Aeonium-клада) и Восточноазиатский (Telephium-клада). Происхождение и история эволюции американской ветви семейства (Acre-клада) наименее изучены. Считают, что большая часть ее членов имеет автохтонное происхождение, а их предки попали в Мексику через Макаронезию (Ham, Hart, 1998).
Распространение Sedoideae приурочено в основном к Голарктическому флористическому царству, здесь встречаются около 500-600 видов подсемейства (табл. 4), преимущественно в субтропической и умеренной зонах Евразии. В европейской части ареала, Северной Африке, Малой и Средней Азии распространены около 110 видов. Две трети от общего числа европейских очитковых приурочены к районам, тяготеющим к Средиземному морю (Борисова, 1939; Webb, 1964, Hart, 1987, 1991; Бялт, 2001). Для большинства «европейских» Sedoideae восточные границы ареалов, более или менее соответствуют 60о восточной долготы, что является определенной границей между европейскими и сибирско-азиатскими флорами (Hart, 1982). В Сибири, Центральной и Восточной Азии распространены около 250 видов, ареалы которых, в своем большинстве, также не выходят за пределы региона. В Северной Америке также произрастают около 250 видов Sedoideae, за исключением Rhodiola rosea, R. integrifolia и Sedum villosum, все они эндемичны для континента (Spongberg, 1978; Hart, 1982; Thiede, 1995). Наибольшее число видов здесь сконцентрировано в Мексике. J. Thiede (1995) приводит 102 вида подсемейства, 93 из которых эндемичны для трансмексиканского вулканического пояса. Необходимо отметить, что высокая степень эндемизма характерна и для других подсемейств, представители которых распространены в Мексике, например, Echeveria – 104 (101 эндемик), Villadia – 31 (29), Dudleya – 25 (17) (Thiede, 1995). Можно предположить, что Мексика является одним из современных центров разнообразия и видообразования семейства. Поразительны отмеченные здесь процессы гибридизации – более 1100 нототаксонов являются продуктом гибридизации (естественной и искусственной) 165 видов родов Echeveria, Sedum, Pachyphytum, Graptopetalum и др. C.H. Uhl (1976) предположил, что толстянковые Мексики (подсем. Sedoideae и Echeverioideae) представляют собой единый биосистематический компариум.
Американские, азиатские и европейские виды Sedoideae образуют три четко очерченные и, предположительно, древние группы (Hart, 1982, 1987).
Таблица 4
Роды Sedoideae с приблизительным числом видов и распространением
Род |
Общее число видов |
Распространение |
Примечание |
Sedum s.str. |
470 (400*) |
Северное полушарие, Южная Америка, Северная, Центральная и Восточная Африка |
*Eggli et al ., 1995 |
Orostachys |
15 (22*) |
Центральная и Восточная Азия |
*Бялт, 1999б |
Hylotelephium |
30 |
Европа, Центральная и Восточная Азия, Северная Америка* |
* Eggli et al ., 1995 |
Rhodiola |
60 (90*) |
Восточная Азия, Гималаи, Сибирь, Европа, Северная Америка |
*Fu; Ohba 2001 |
Phedimus |
5* |
Европа (Ирано-Туранский регион, Кавказ) |
*Гончарова, настоящее исследование |
Aizopsis |
15* |
Восточная Азия, Сибирь |
*Гончарова, настоящее исследование |
Meterostachys |
1 |
Восточная Азия |
|
Sinocrassula |
3 |
Гималаи |
|
Pseudosedum |
10 (2*) |
Центральная Азия, Анатолия, Северный Иран, Кавказ* |
* Eggli et al ., 1995 |
Rosularia |
20 |
Восточная Европа, Гималаи, Алтай |
|
Prometheum |
8 |
Восточная Европа, Центральная Азия |
|
Perrierosedum |
1 |
Мадагаскар |
|
Число видов без звездочки дано по: Mayzumi, Ohba, 2004, со звездочками – по источникам, указанным в примечании.
Сравнение видовых списков очитковых Европы, Америки, западно-азиатских и восточно-азиатских флор (Борисова, 1939; Ohwi, 1965; и др.) показало, что лишь незначительное число видов (около 5) являются общими для них. Вполне вероятно, что процессы видообразования шли независимо в каждой из областей (Grulich, 1984).
В пределах Старого Света виды подсемейства также распространены весьма неравномерно. Большинство видов родов Orostachys, Aizopsis, Hylotelephium и почти все представители Rhodiola (табл. 5) произрастают в Восточной Азии. Наибольшее число видов подсемейства приводится для флоры Китая, здесь встречаются около 210 видов, 117 из которых – эндемики (Fu, Ohba 2001). В Японии подсемейство представлено 39 таксонами видового и внутривидового ранга, из них – 21 эндемик (Ohba, 2001), на российском Дальнем Востоке – 35/11 (Гончарова, 2000б, в), в Сибири – 25/3 (Пешкова, 1994), в Корее – 19 видами (Lee, 1993), Монголии – 14 (Грубов, 1982; Губанов, 1996).
Распространение Sedoideae на территории России и прилегающих регионов
Регион |
|
Rhodiola
(таксонов) |
Hylotelephium (таксонов) |
Orostachys (таксонов) |
Aizopsis (таксонов) |
Meterostachys (таксонов) |
Sedum s.str (таксонов) . |
Pseudosedum (таксонов) |
Источник |
Китай |
207 |
53 |
16 |
8 |
8 (как Phedimus ) |
|
120 |
2 |
Fu; Ohba , 2001 |
Япония |
39 |
2 |
10 |
3 |
3 (как Phedimus ) |
1 |
18 |
|
Ohba, 2001 |
Монголия |
14 |
5 |
3 (как Sedum s.l. ) |
4 |
2 (sensu Sedum s.l. ) |
|
|
|
Грубов, 1982
Губанов, 1996 |
Корея |
18 |
4 |
5 (как Sedum s.l. ) |
3 |
3 (как Sedum s.l. ) |
1 |
6 |
|
Illustrated flora of Korea, 1993 |
Европа |
57 |
2 |
5 (как Sedum s.l.) |
2 |
2 (как Sedum s.l.) |
|
47 (54 * ) |
|
Webb, 1964
* Hart, 1991 |
Восточноевропейс-кая часть России |
43 |
4 |
12 |
3 |
3 |
|
18 (включая Macrosepalum и Petrosedum ) |
1 |
Бялт, 2001 |
Средняя Россия |
3 |
|
5 |
|
4 |
|
1 |
|
Губанов и др., 2003 |
Сибирь |
25 |
7 |
7 (как Sedum s.l.) |
4 |
4 (как Sedum s.l.) |
|
2 |
|
Пешкова, 1994 |
Российская Арктика |
8 |
4 |
2 (как Sedum s.l.) |
|
1 (как Sedum s.l.) |
|
1 |
|
Петровский, 1984 |
Российский Дальний Восток |
35 |
8 |
9 |
8 |
10 |
|
|
|
Гончарова, настоящее исследование |
Примечание. Названия регионов даны в соответствии с таковыми, представленными в региональных флорах.
Анализ распространения евро-азиатских представителей Sedoideae позволяет выделить несколько очагов их видового разнообразия – «Средиземноморский», приуроченный к Иберийскому полуострову, Эгейскому региону и Южным Балканам; «Ирано-Туранский» – к Кавказу, горам Восточной Анатолии и Северному Ирану и «Тибетско-Гималайский» – от Каракорума до Юго-Восточного Китая, Гималаи и Тибет. Южнее названных районов разнообразие видов резко уменьшается, единичные виды встречаются в тропической восточной Африке (5), Вьетнаме (1) (Hart, 1982; Ohba, 1989; Гончарова, 1999). Примечательно, что центры видового разнообразия приходятся на семиаридные субтропические высокогорные области, а южная граница распространения Sedoideae совпадает с границами между семиаридными и аридными областями Северного полушария.
Наиболее часто очитковые встречаются в горных и высокогорных районах. В горах они достигают субальпийских и иногда альпийских высот [в Гималаях до 5100 м, в Пакистане до 5400 м (Вальтер, 1968)]. Довольно обычны представители Sedoideae на морских побережьях и сравнительно немно гочисленны в континентальных равнинных районах. Неравномерность географического распространения свидетельствует как об истории формирования ареалов очитковых, так и о специфике биологии и экологии представителей подсемейства. Большинство видов занимает довольно узкие экологические ниши, они светолюбивы, не переносят застойного переувлажнения почвы и слабо конкурентоспособны. Например, небольшое число Sedoideae, произрастающих на Западно-Сибирской низменности, можно объяснить большим количеством заболоченных местообитаний, а Прикаспийской – наличием массивов подвижных песков (Бялт, 2004а). Только несколько наиболее широко распространенных видов (Orostachys spinosa, Aizopsis aizoon, Hylotelephium triphyllum и некоторые другие) способны занимать широкий спектр местообитаний – от скальных до луговых и даже лесных.
Как сказано выше, максимальное число видов и предполагаемые центры видового разнообразия приходятся на области с высокогорными сообществами, характеризующиеся значительным разнообразием климатических условий, резкими колебаниями дневных и ночных температур, небольшим количеством осадков, выпадающих, как правило, летом (Вальтер, 1968, 1974, 1975). Сухих местообитаний почти не встречается, даже на кажущихся сухими скальных поверхностях или каменистых осыпях, в трещинах скал и между камнями скапливается всегда влажный мелкозем (Вальтер, 1968, 1974, 1975). Значительный источник влаги здесь также – тающий снеговой покров (Толмачев, 1986). Разнообразие климатических и эдафических условий и послужило, на наш взгляд, причиной формирования значительного морфологического разнообразия видов и биоморф, встречающихся в названных областях.
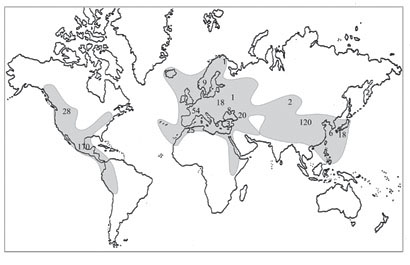
Рис. 28. Распространение рода Sedum.
Арабскими цифрами обозначено число видов, серым цветом – ареал рода.
|
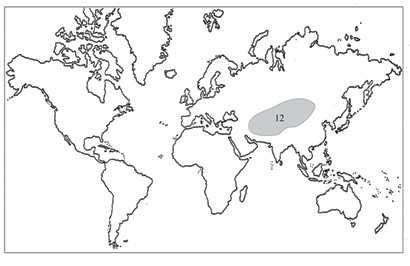
Рис. 29. Распространение рода Pseudosedum.
|
Характеризуя распространение отдельных родов изучаемых Sedoideae, можно отметить, что самый обширный ареал имеет род Sedum (рис. 28). По современным данным (Hart, Bleij, 2005) наибольшее число видов (около 240) произрастают в Америке, около 130 – в Восточной Азии, 100 – в Европе, Северной Африке, Малой и Средней Азии. В Сибири встречаются 2 вида, а на РДВ род не представлен, хотя границы ареала очень близко подходят к российскому Дальнему Востоку, представители рода обычны в северо-восточном Китае, Японии и Корее. Sedum albertii довльно широко распространен в Азии (Ирано-Туранской флористической области) от Улу-Тау до Тибета, по территории «русского» Алтая [долина р. Чибитки (Пяк, 2003)] проходит северная граница ареала вида. Sedum acre – широко распространен по всей Европе, обычен в Северной Африке, Турции, в Западной Сибири проходит восточная граница ареала вида.
Ареал рода Pseudosedum (рис. 29) расположен преимущественно в Центральной Азии (Иране, Афганистане, Таджикистане, Казахстане), Ирано-Туранской флористической области, наиболее многочисленны виды в Памиро-Алайском регионе (Центральноазиатской подобласти), достигая здесь высот 3600 м (Hart, 2005). Во флоре Сибири род представлен одним, наиболее широко распространенным видом – Pseudosedum lievenii. По направлению на восток вид достигает восточных провинций Китая (Ксиньян), а на север – подножий русского Алтая.
Ареал Rhodiola – один из самых больших в подсемействе. Однако широкое распространение определяется ареалом лишь одного вида – Rhodiola rosea s. l., охватывающим арктические районы Нового и Старого Света, Альпы и другие высокогорья Европы, Восточной Азии, восточной части Северной Америки. Большинство же видов Rhodiola сосредоточено в Восточно-Гималайской провинции Восточно-Азиатской флористической области (рис. 30) и Тибетской провинции Ирано-Туранской, которые, вероятно, являются центром происхождения и диверсификации рода. Из почти 60 его представителей 44 распространены в высокогорных районах от Памиро-Алая, Гималаев, Бирмы и Тибета до Юго-Западного Китая (главным образом Юннань и Сычуань) (Ohba, 1989). В европейской и американской частях ареала рода встречаются по 3-4 представителя. Rhodiola iremelica A. Boriss. – единственный вид, эндемичный для Восточной Европы (хр. Зигальга, хр. Крыкты) (Бялт, 2001), а R. rhodantha (A. Gray) Jacobson – для Северной Америки (Скалистые горы) (Spongberg, 1978). По утверждению H. Ohba (1989), ареал рода представляет собой один из примеров, демонстрирующих взаимодействие флор высокогорных и арктических районов.
Из пяти секций подрода Rhodiola представители трех встречаются в Сибири и на РДВ. Секция Chamaerhodiola, распространенная в альпийских и высокогорных районах Евразии от Гималаев (Кашмир, Бутан), Тибета, северо-западного Китая, Монголии, Урала до Сибири (Ohba, 1989), в изучаемом регионе представлена двумя видами – R. coccinea и R. quadrifida.
R. coccinea широко распространена в Азии, от Тянь-Шаня, Памиро-Алая, до Гималаев и Тибета, это один из доминантных видов родиол в альпийско-высокогорных районах Гималаев (Ohba, 1982). Она обычна в Восточном Непале на высотах от 4400 до 4800 м. В Сибири, в пределах русского Алтая проходит северная граница распространения R. coccinea, здесь она также является обычным элементом флоры [особенно на территории Юго-Восточного Алтая] (Пяк, 2003). Ареал R. quadrifida в северо-западном направлении достигает Большеземельской тундры и Полярного Урала (Петровский, 1984; Бялт, 2001).
Большинство представителей секций Rhodiola и Algida встречаются в центрально-азиатских высокогорьях. R. integrifolia распространена по обеим сторонам Берингова пролива, главным образом в арктических и субарктических регионах как Нового, так и Старого Света, преимущественно там, где R. rosea отсутствует (Ohba, 1989). Два подвида: R. rosea subsp. krivochizhinii (о-в Верхотурова) и R. rosea subsp. sachalinensis (о-в Сахалин) – эндемичны для российского Дальнего Востока. R. ishidae и R. angusta распространены исключительно в Северо-Восточной Азии – в Маньчжурской, Сахалино-Хоккайдской и Японо-Корейской провинциях Восточно-Азиатской флористической области, а R. algida – в высокогорьях Сибири и северо-западных районах Монголии и Китая (Алтае-Саянская провинция Бореальной области и Джунгаро-Тяньшанская, Монгольская и отчасти Тибетская провинции Ирано-Туранской).
Виды подрода Crassipedes распространены главным образом в Восточно-Азиатской флористической области, Джунгаро-Тяньшанской и Тибетской провинциях Ирано-Туранской области [от Гималаев до восточных районов Бирмы, в северных районах Тибета и Китая (Ohba, 1989)] и Циркумбореальной (Забайкальской и Северовосточно-Сибирской провинции). В Сибири и на РДВ встречаются три представителя подрода – R. stephaii, R. pinnatifida subsp. pinnatifida и R. pinnatifida subsp. subpinnata. Большая часть ареала R. stephaii находится в Маньчжурской провинции. Вид заходит в область российской Арктики [низовья Пенжины, район Гижигинской губы, (Петровский, 1984)], а на юге достигает северных районов Китая. R. pinnatifida subsp. pinnatifida и R. pinnatifida subsp. subpinnata распространены преимущественно в Алтае-Саянской провинции: на Монгольском Алтае, Хангае и Юго-Восточном Алтае, по территории Русского Алтая проходит северо-западная граница распространения этих видов (Пяк, 2003).
В роде Rhodiola выделяются несколько географических групп (Ohba, 1989). Это виды: 1) распространенные вдоль западной или восточной границы «центрально-азиатского высокогорного коридора», мигрировавшие из арктических районов по направлению на юг к Гималаям, Юго-Западному Китаю и наоборот (R. algida, R. quadrifida и др.); 2) мигрировавшие в направлении с востока на запад и с запада на восток, от Памиро-Алая до Юго-Западного Китая через Гималаи, Тибет и Северную Бирму (большинство видов рода); 3) распространенные по всей Арктике, включая Старый и Новый Свет (Rh. rosea, Rh. integrifolia); 4) распространенные на Дальнем Востоке (R. stephanii, R. angusta, R. ishidae и др.).
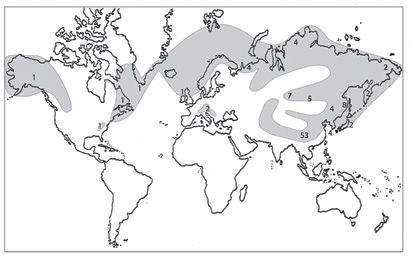
Рис. 30. Распространение рода Rhodiola (по: Ohba, 1989, с дополнениями). Арабскими цифрами обозначено число видов.
|
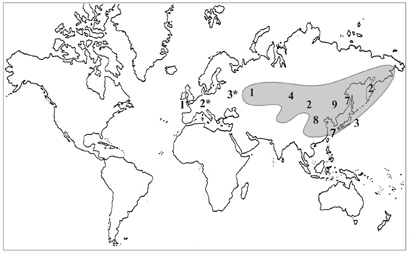
Рис. 31. Распространение рода Aizopsis. (по: Grulich, 1984, с дополнениями). * – заносные и натурализовавшиеся виды.
|
Подобные тенденции в целом характерны и для большинства других восточно-азиатских родов очитковых.
Виды рода Aizopsis имеют преобладающее распространение в пределах Восточно-Азиатской флористической области (рис. 31). Центр разнообразия рода смещен относительно «Тибетско-Гималайского» центра разнообразия подсемейства в направлении российского Дальнего Востока, где встречаются большинство представителей рода. Форму ареала рода определяет ареал секции Aizopsis. А. aizoon (включая Aizopsis aizoon subsp. baicalensis) – самый широко распространенный представитель рода, по направлению на запад он достигает Алтае-Саянской провинции, а на север – едва заходит в Северо-восточносибирскую провинцию Циркумбореальной области. Вид довольно пластичен экологически, что позволяет ему занимать широкий спектр местообитаний – от скальных до луговых. В настоящее время ареал как вида, так и рода значительно расширился из-за активной интродукции и, как следствие, натурализации вида.
В «сибирской» части ареала рода произрастает A. hybridа – единственный представитель рода, большая часть ареала которого лежит вне Восточно-Азиатской флористической области. Этот вид распространен от Урала до Средней Азии и северных районов Монголии, главным образом в Алтае-Саянской провинции Циркумбореальной области, Джунгаро-Тяньшанской и Монгольской провинциях Ирано-Туранской. Дальше всех на север до низовий Колымы, Анюйского нагорья и бассейна р. Пенжина из живучников распространен A. kamtschatica, достигая Арктической провинции Циркумбореальной флористической области. Ареал A. selskiana полностью находится в пределах Восточно-Азиатской флористической области, главным образом в Маньчжурской провинции, в западной части ареала достигая Хэйлунцзяна, Джилина и Ляонина, а в северной – Тернейского района Приморского края.
Пять таксонов эндемичны для флоры РДВ – A. kurilensis (о-ва Шикотан, Кунашир), A. litoralis и A. × pilosa (юг Приморского края), A. middendorfiana subsp. sichotensis [Южный Сихотэ-Алинь (Ворошилов, 1961; Гурзенков, 1967)] и A. pseudohybrida [Амурская обл., Зейский р-н, истоки р. Маи (Ворошилов, Шлотгауэр, 1984)].
Виды рода Hylotelephium также имеют преимущественное распространение в Восточно-Азиатской флористической области (рис. 32), но они более широко распространены, чем представители Aizopsis, и встречаются как в Старом, так и Новом Свете. В «американской» части ареала род представлен единственным представителем – Hylotelephium telephiuoides (Michaux) H. Ohba (эндемичным для Аппалачей). Род ограниченно представлен во всех трех центрах видового разнообразия, однако только один вид Hylotelephium telephium s.l. (включая H. triphyllum) распространен практически по всему Голарктису от Северной Америки до Восточной Азии, от арктических районов Евразии до Северного Китая и Японии (Хоккайдо). Другие виды рода имеют достаточно четко ограниченные ареалы, тяготеющие к одному из центров. В Восточной Азии центр разнообразия рода несколько смещен относительно «Тибетско-Гималайского центра», только один вид Hylotelephium ewersii встречается в Гималаях (Ohba, 1977), наиболее многочисленны виды очитников в Китае (16) и Японии (10). Из 30 таксонов 19 являются эндемичными для различных регионов: 7 видов и подвидов – для относительно небольшой территории Японских островов, 6 – для Китая, 2 (H. pluricaule и H. ussuriense) – для российского Дальнего Востока, 2 (H. populifolium и H. triphyllum subsp. sukaczevii) – Сибири, H. × zhigulienze Tzvel. и H. uralense (Rupr.) V. Byalt – Европы и H. caucasium (Gross.) H. Ohba – Кавказа.
Дальше всех очитников на север наряду с H. triphyllum распространен H. cyaneum. Последний незначительно заходит за северную границу леса на пространстве между р. Колымой и побережьем Берингова моря (Петровский, 1984). Ареал H. cyaneum приурочен к северо-восточной части Циркумбореальной (Северовосточносибирской и Охотско-Камчатской провинциям) и Восточно-Азиатской флористической области (Маньчжурской и Сахалино-Хоккайдской провинциям). H. pallescens распространен главным образом в Забайкальской и Маньчжурской провинциях. Большее число очитников флоры РДВ имеют ареал преимущественно в Маньчжурской, Сахалино-Хоккайдской и Японо-Корейской провинциях Восточно-Азиатской флористической области (H. erythrostictum H. verticillatum H. viviparum, H. pluricaule и H. ussuriense).
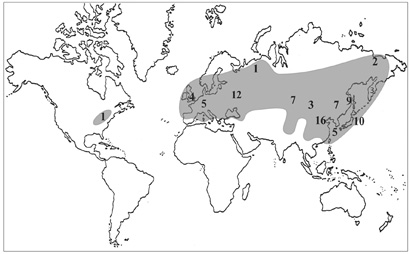
Рис. 32. Распространение рода Hylotelephium.
|
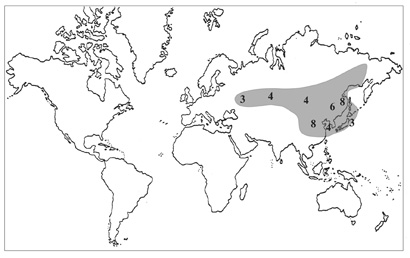
Рис. 33. Распространение рода Orostachys.
|
Представители рода Orostachys встречаются в трех флористических областях: Циркумбореальной, Ирано-Туранской и Восточно-Азиатской, с наибольшим числом видов в последней (рис. 33). Здесь произрастают все известные виды рода (Бялт, 1999а). Как и у Hylotelephium, центр разнообразия рода несколько смещен относительно «Тибетско-Гималайского центра», наиболее многочисленны виды горноколосников в Китае (8/2 – эндемичных), на РДВ – (8/3), Японии (5-8/2). Вне РДВ встречаются лишь 2 представителя Orostachys флоры России – O. thyrsiflora, O. fimbriata. Вполне вероятно, что эти виды, подобно другим представителям «сибирских» Sedoideae, мигрировали вдоль «центрально-азиатского высокогорного коридора» в направлении на северо-восток, тогда как большинство видов рода – на восток. Ареалы O. thyrsiflora и O. fimbriata расположены преимущественно в Ирано-Туранской флористической области (Центральноазиатской подобласти), на территории Сибири проходит северо-восточная граница их распространения.
Среди представителей рода наиболее широко распространены O. malacophylla и O. spinosa. Границы ареала горноколосника колючего в направлении на запад достигают европейской части России [окр. г. Златоуста (Бялт, 2001)] на юго-запад – Северо-Восточного Китая, на север – Магадана, а горноколосника мягколистного – Западной Сибири на западе и северо-восточных провинций Китая на юге.
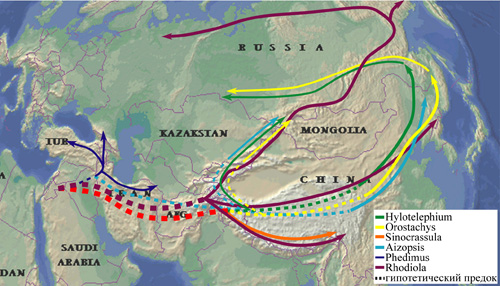
Рис. 34. Предполагаемые пути расселения и диверсификации восточно-азиатских родов Sedoideae. При построении схемы учитывалось современное распространение родов и филогенетические взаимоотношения, установленные при анализе нуклеотидных последовательностей ДНК.
|
Как мы уже отмечали выше, во флоре Сибири и российского Дальнего Востока встречаются 51 вид и подвид Sedoideae, лишь 9 из них являются общими для обоих регионов. Состав толстянковых флоры РДВ имеет явно выраженный восточно-азиатский характер, а Сибири – испытывает явное влияние Ирано-Туранской флоры, представителями которой можно назвать Sedum albertii и Pseudosedum lievenii. Анализируя распространение Sedoideae флоры Сибири и российского Дальнего Востока (рис. 28-33), можно предположить, что как сибирские, так и дальневосточные представители имеют общее происхождение. Различие и сходство видовых составов Sedoideae флор российского Дальнего Востока и Сибири, центром происхождения которых, вероятно, является Тибетско-Гималайский центр разнообразия семейства, обусловлено, на наш взгляд, различием путей миграции видов рис. 34). «Сибирская ветвь» сформировалась при распространении видов в направлении на северо-восток вдоль «центрально-азиатского высокогорного коридора», через северо-западные районы Китая и Монголию (Orostachys thyrsiflora, Aizopsis hybrida, Rhodiola coccinea и др.), вплоть до арктических районов Евразии и через Берингийский мост до северо-западных районов Северной Америки (Rhodiola quadrifida, R. rosea), и с востока на запад из районов РДВ (Aizopsis aizoon, Orostachys spinosa и др.). «Дальневосточная ветвь» – в направлении на восток. Необходимо отметить, что флора толстянковых РДВ имеет большее число эндемичных видов, по сравнению с толстянковыми Сибири (11 и 3, соответственно). Что свидетельствует, вероятно, о более благоприятных эколого-климатических условиях этого региона для развития Sedoideae.
|
6.2. Редкие виды очитковых флоры Сибири и российского Дальнего Востока
Как отмечено выше, редкие виды Sedoideae флоры Сибири изучены очень хорошо, что, к сожалению, нельзя сказать об очитковых российского Дальнего Востока.
По литературным данным 6 видов очитковых являются редкими для флоры Сибири (Редкие и исчезающие растения Сибири, 1980), и еще 6 – для отдельных ее районов (Красная книга республики Алтай (растения), 1996; Красная книга республики Тыва, 1999; Красная книга республики Саха (Якутия), 2000; Красная книга Иркутской области: Сосудистые растения, 2001; Красная книга Томской области, 2002; Красная книга республики Хакасия, 2002; Красная книга Хантымансийского автономного округа, 2003). И только три вида отнесены к редким на РДВ (Харкевич, Качура, 1981; Редкие и исчезающие.., 1983, Перечень объектов…, 2002). Это – Hylotelephium ussuriense, Rhodiola rosea и Rh. angusta. Однако изучение распространения представителей подсемейства флоры РДВ и Сибири показывает, что в списки редких и подлежащих охране видов должны быть включены еще несколько представителей очитковых.
При разделении видов на «редкие» и «обычные» мы следовали следующему определению: «вид редкий – не находящийся под прямой угрозой исчезновения, но встречающийся в таком малом числе особей и/или популяций либо на столь ограниченной территории и в столь специфических местах обитания, что может исчезнуть» (Реймерс, 1990, с. 90). И учитывали два основных критерия: 1) встречаемость – плотность популяции в конкретных местообитаниях, степень концентрированности популяции; и 2) распространенность – число и размер населенных данными организмами участков в пределах всего ареала (Бигон и др., 1989).
Кроме того, среди редких видов также необходимо разделять «собственно редкие виды» – имеющие низкую встречаемость и распространенность в пределах всего ареала и «территориально редкие» – редкие для конкретной части ареала (региона, страны), но обычные в других его частях. В связи с этим, мы считаем необходимым выделить виды: а) рекомендуемые для государственной охраны (редкие для России); б) редкие для Сибири или РДВ; в) редкие для отдельных административных районов. К последней группе нами отнесены виды, представленные в рассматриваемом регионе либо граничными популяциями, либо локусами дизъюнктивного ареала, но обычные в других регионах.
По нашему мнению, виды, эндемичные для флоры региона, также необходимо вносить в списки растений, подлежащих охране, независимо от обилия и размеров ареала (табл. 6).
Таблица 6
Редкие виды Sedoideae флоры Сибири и российского Дальнего Востока
№ |
Вид |
Категория редкости |
Тип редкости [по: Rabinowitz, (Бигон и др., 1989)] |
Охраняемые территории* |
Редкие для России |
1 |
Aizopsis litoralis |
э |
EN |
локально обильный в специфических местообитаниях с ограниченным ареалом |
заповедник «Кедровая падь» |
2 |
Hylotelephium
populifolium |
э |
VU |
локально обильный в специфических местообитаниях с ограниченным ареалом |
|
3 |
Orostachys gorovoii |
э |
LR |
редко встречающийся в специфических местообитаниях и с ограниченным ареалом |
|
4 |
Orostachys paradoxa |
э |
LR |
локально обильный в специфических местообитаниях и с ограниченным ареалом |
|
5 |
Rhodiola algida |
сэ |
VU |
редко встречающийся в специфическом местообитании с ограниченным ареалом |
Катунский и Алтайский заповедники |
6 |
Rhodiola angusta |
|
VU |
редко встречающийся в специфическом местообитании с ограниченным ареалом |
|
7 |
Rhodiola rosea subsp. rosea |
|
EN |
локально-обильный в широком ареале, но специфических местообитаниях |
Сихотэ-Алинский, Олекминский, Катунский и Алтайский заповедники, ресурсный резерват «Бол. Токо» |
|
Редкие для РДВ |
|
|
|
|
8 |
Aizopsis kurilensis |
э |
DD |
|
Южно-Курильский заповедник, заказник «Малые Курилы» |
9 |
Aizopsis middendorfianа subsp. arcuatа |
э |
DD |
|
|
10 |
Aizopsis middendorfiana subsp . sichotensis |
э |
LR |
локально обильный в нескольких местообитаниях и с ограниченным ареалом |
Лазовский, Буреинский и Магаданский заповедники |
11 |
Aizopsis ? pilosa |
э |
LR |
локально-обильный в специфических местообитаниях и с ограниченным ареалом |
|
12 |
Aizopsis pseudohybrida |
э |
DD |
|
|
13 |
Hylotelephium pseudospectabile |
|
VU |
локально обильный с ограниченным ареалом |
|
14 |
Hylotelephium pluricaule |
сэ |
LR |
локально обильный в нескольких местообитаниях и с ограниченным ареалом |
Поронайский заповедник |
15 |
Hylotelephium ussuriense |
э |
LR |
локально обильный в нескольких местообитаниях и с ограниченным ареалом |
Лазовский заповедник |
16 |
Orostachys maximowiczii |
э |
EN |
локально обильный в специфических местообитаниях с ограниченным ареалом |
заповедник «Кедровая падь» |
17 |
Orostachys furusei |
|
DD |
локально-обильный в специфических местообитаниях и с ограниченным ареалом |
|
18 |
Rhodiola rosea subsp. krivochizhinii |
э |
DD |
локально-обильный в специфических местообитаниях и с ограниченным ареалом |
|
19 |
Rhodiola rosea subsp. sachalinensis |
э |
VU |
локально-обильный в широком ареале, но в специфических местообитаниях |
Южно-Курильский заповедник, заказник «Малые Курилы» |
20 |
Rhodiola ishidae |
|
DD |
вероятно, локально-обильный в специфических местообитаниях с ограниченным ареалом |
Южно-Курильский заповедник |
|
Редкие для Сибири |
|
|
|
|
21 |
Aizopsis hybridа |
|
LR |
локально-обильный в специфических местообитаниях с ограниченным ареалом |
|
22 |
Rhodiola quadrifida |
|
VU |
локально-обильный в специфических местообитаниях с ограниченным ареалом |
заповедники Катунский и Алтайский, ресурсный резерват «Бол. Токо» |
23 |
Rhodiola borealis |
э |
LR |
локально-обильный в специфических местообитаниях с ограниченным ареалом |
«Усть-Ленский» заповедник, памятник природы «Походская едома» |
24 |
Rhodiola coccinea |
|
LR |
локально-обильный в специфических местообитаниях с ограниченным ареалом |
|
25 |
Rhodiola pinnatifida ssp. pinnatifida |
сэ |
VU |
локально-обильный в специфических местообитаниях с ограниченным ареалом |
заповедник Катунский |
26 |
Rhodiola pinnatifida ssp. subpinnata |
сэ |
VU |
локально-обильный в специфических местообитаниях с ограниченным ареалом |
заповедник Катунский |
27 |
Sedum albertii |
|
LR |
редко встречающийся в специфическом местообитании с ограниченным ареалом |
|
|
Редкие для отдельных административных районов |
|
|
|
|
28 |
Aizopsis aizoon |
|
LR |
на границе ареала |
редкий для Томской обл. |
29 |
Hylotelephium erythrostictum |
|
DD |
на границе ареала |
редкий для Сахалинской обл., заказник «Малые Курилы» |
30 |
Hylotelephium verticillatum |
|
LR |
на границе ареала |
редкий для Приморского края |
31 |
Orostachys spinosa |
|
LR |
на границе ареала |
редкий для Томской обл. |
Категории риска приводятся по: IUCN Red List Categories (1994).
* Сведения об охраняемых территориях приведены по литературным данным (Жудова, 1967; Нечаева, 1972; Присяжнюк, Форш, 1975; Бромлей, Васильев и др., 1977; Безделева, Харкевич, 1978; Куренцова, 1981; Флора, мико- и лихенобиота Лазовского заповедника, 1990; Флора охраняемых территорий…, 1998; Красная книга республики Алтай (растения), 1996; Красная книга республики Тыва, 1999; Галанин и др., 2000; Красная книга республики Саха (Якутия), 2000; Красная книга Иркутской области: Сосудистые растения, 2001; Красная книга Томской области, 2002; Красная книга республики Хакасия, 2002; Баркалов, Еременко, 2003; Красная книга Хантымансийского автономного округа, 2003).
Особенности биологии и экологии Crassulaceae вообще и Sedoideae в частности определяют специфику распространения видов, только несколько представителей подсемейства можно назвать обычными и широко распространенными (например, Aizopsis aizoon и Hylotelephium triphyllum). Анализ литературных данных и собственные исследования (табл. 6) позволяют нам рекомендовать для государственной охраны семь видов очитковых Сибири и РДВ, три из них эндемичны для флоры РДВ (Aizopsis litoralis, Orostachys gorovoii и O. paradoxa), два – для Сибири (Hylotelephium populifolium, Rhodiola algida). Rhodiola angusta – чрезвычайно редкий вид на РДВ, редок он и на протяжении всего ареала [в Северном Китае (Хэйлунзян) и Северной Корее, на высотах 1700-2600 м]. В Уссурийском районе известно лишь два места произрастания: «Приморский край, Чугуевский район, скальные выходы на вершине г. Облачная (1850 м над ур.м.)», (популяция имеет площадь около 25 м2) и «Никольск-Уссурийский уезд, склон г. Хуалаза, скалистый склон вблизи вершины, по краю обрывов. 3.6.1921. И. Шишкин» (LE). Поиски Rhodiola angusta на г. Криничная (Хуалаза), по экземплярам c которой А.Г. Борисовой (1939) была описана R. komarovii, и на близлежащих вершинах (Снежная, Воробей, Ливадийская), характеризующихся близкими высотами и сходными условиями местообитаний, что и гора Криничная, результатов не дали. В единичных местообитаниях встречаются также Orostachys gorovoii и O. paradoxa. Из видов этой группы наиболее пристальное внимание следует уделить Rhodiola rosea subsp. rosea и Aizopsis litoralis (категория EN). Первый вид подвергается угрозе исчезновения вследствие массовых заготовок, а ареал второго существенно сокращается из-за добычи песка на пляжах и массового отдыха в районах его естественного произрастания.
Для флоры Сибири, основываясь главным образом на литературных данных, мы также предлагаем 7 видов (табл. 6). Это, в основном, виды локально обильные в специфических местообитаниях, в настоящий момент не подвергающиеся серьезной угрозе существованию. Среди них эндемичный вид (Rhodiola borealis) и два субэндемичных подвида (R. pinnatifida ssp. pinnatifida, R. pinnatifida ssp. subpinnata), встречающиеся кроме Сибири и в Северной Монголии. Чрезвычайно редким является лишь Sedum albertii, известный в Сибири только из нескольких местонахождений, однако довольно широко распространенный в Центральной Азии.
Для российского Дальнего Востока редкими являются 7 видов (табл. 6). В большинстве своем это также локально-обильные виды, обитающие в специфических местообитаниях и с ограниченным ареалом. Некоторые из них известны из довольно значительного числа местообитаний, некоторые же – из единичных. Например, для Hylotelephium pseudospectabile известно единственное местообитание на РДВ: «Приморский край, Хасанский район, сопка Приозерная, середина первого пика, скалы. 25, VIII, 2003. В.В. Якубов, С.Б. Гончарова, А.А. Гончаров». Хотя, вполне вероятно, что вид обычен в северных районах Китая и Кореи. На наш взгляд, особое внимание необходимо обратить на охрану Orostachys maximowiczii, так как в настоящее время места произрастания вида в Хасанском районе значительно сокращаются из-за массового отдыха и добычи песка на пляжах.
Большую группу составляют виды (6), данных о состоянии которых недостаточно, и на современном этапе сложно дать оценку их состоянию. Необходимы дополнительные исследования в местах их естественного произрастания. По определению С.С. Харкевича и Н.Н. Качуры (1981, с. 12), виды этой категории составляют временную группу, «изучение их в природе даст возможность выявить их состояние и решить вопрос об их охране, или же об исключении из кадастра подлежащих охране видов».
Для видов других категорий редкости, также определены соответствующие меры охраны. Это, в первую очередь, ограничение или запрет сбора декоративных, лекарственных и иных полезных растений, контроль за состоянием вида в природе. Многие виды, рассматриваемые нами в качестве редких, входят в состав флор охраняемых територий Сибири и российского Дальнего Востока, а, следовательно, в той или иной степени охраняются (табл. 6) (Жудова, 1967; Нечаева, 1972; Присяжнюк, Форш, 1975; Бромлей, Васильев и др., 1977; Безделева, Харкевич, 1978; Куренцова, 1981; Флора, мико- и лихенобиота Лазовского заповедника, 1990; Флора охраняемых территорий…, 1998; Галанин и др., 2000; Баркалов, Еременко, 2003). Как известно, многие очитковые ценнейшие лекарственные растения. Их целебные свойства были известны еще в Древней Греции (Regnat, 2004). Очень популярны толстянковые и в озеленении (Биологическое разнообразие и интродукция суккулентов, 2004). Для таких видов редких растений изучение полезных свойств, их значения в декоративном цветоводстве и садоводстве и перенос в коммерческую культуру – лучшее средство сохранения вида на Земле (Харкевич, 1959, 1971; Цицин, 1976 а,б; Храпко, 1979).
Бесспорно, что на численность вида влияет комплекс факторов, выявить полностью которые практически невозможно. Но, как правило, можно выделить наиболее значимые из них, определяющие причины редкости и сокращения численности. Большинство из предлагаемых для охраны видов не подвергаются непосредственной угрозе исчезновения, их редкость обусловлена, на наш взгляд, особенностями их биологии. Суккуленты вообще и очитковые, в частности, устойчивы к неблагоприятным внешним условиям, они могут расти на кислых, щелочных и нейтральных почвах с недостаточным и избыточным увлажнением, но они, как правило, требовательны к условиям освещения и не способны конкурировать с другими видами в борьбе за свет, именно в этом, по мнению В. Вальтера (1968), и проявляется биологический консерватизм подобных видов – основная, на наш взгляд, причина редкости названных выше очитковых.
Оригинальный рисунок А.Р. De Candolle www.illustratedgarden.org/mobot/rarebooks, Rhodiola pinnatifida nature.chita.ru.
Читать далее: главу 7
