|
Подсемейство Sedoideae (Crassulaceae) флоры Сибири и российского Дальнего Востока (систематика, биоморфология, филогения)
С. Б. Гончарова
Глава 4. Морфолого-анатомические признаки Sedoideae флоры Сибири и российского Дальнего Востока, их таксономическое и эволюционное значение
4.1. Генеративные органы
|
Цветок Sedoideae диплохламидный. Формула цветка *KnCnAn+nGn [n=(3) 4–5 (–8)] (рис. 11, 12). Цветки сидячие или на более или менее длинных цветоножках; актиноморфные, изомерные; гермафродитные у большинства представителей. Однополые цветки характерны только для представителей подрода Rhodiola рода Rhodiola, кроме этого, у представителей секции Rhodiola иногда встречаются функционально однополые цветки (женские или мужские), содержащие зачатки тычинок или пестиков. Подобный тип цветка считается переходным от двуполого к однополому (Левина, 1981). Единственный представитель Sedoideae – Rhodiola stephanii – является однодомным растением (Ohba, 1978). Половые различия цветков использованы Н. Ohba (1978) для внутриродового разделения рода Rhodiola. Так, члены подрода Rhodiola – раздельнополые двудомные растения, подродов Primuloides, Crassipedes (за исключением Rhodiola stephanii) и Clementsia – гермафродитные. Однако, как показывают молекулярно-филогенетические исследования (Mayzumi, Ohba, 2004), подобное разделение оказалось искусственным, признак, вероятно, возникал в роде несколько раз.
Чашечка и венчик. Чашелистики как правило изомерные, редко неравные (Sedum acre); более или менее мясистые, вальковатые, от яйцевидных, до почти шиловидных (Sedum albertii), иногда оканчивающиеся шипиком (Orostachys spinosa, O. japonica), голые, редко опушенные (Aizopsis selskiana). Основания чашелистиков свободные (Aizopsis, Sedum acre) либо срастающиеся (Sedum albertii, Orostachys, Pseudosedum); степень срастания чашелистиков значительно варьирует, самая длинная трубка у представителей рода Rhodiola, она образуется при срастании чашелистиков и оснований завязей (Ohba, 1978). Чашечка не опадает во время плодоношения, что считается важным диагностическим признаком подсемейства (Ohba, 1978).
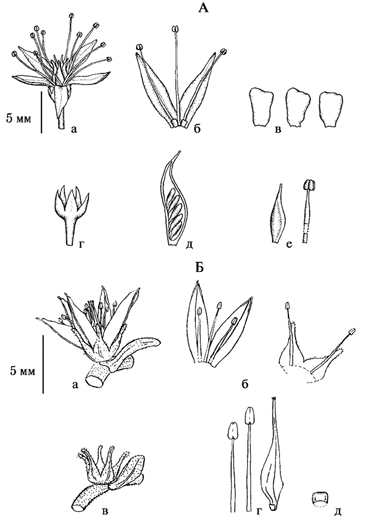
Рис. 11. Строение цветка Sedoideae (по: Гончарова и др., в печати).
А. Hylotelephium pseudospectabile: а – общий вид цветка; б, в, г, д, е – детали строения, б – лепестки, тычинки и нектарники, в – нектарники, г – плодолистик и тычинка, д – чашечка, е – плодолистик с завязями; Б. Aizopsis × pilosa; а – общий вид цветка; б, в, г, д – детали строения цветка, б – лепестки и тычинки, чашелистики и тычинки, в – чашечка, г – тычинки и плодолистик с нектарником, д – нектарники.
|
Рис. 12. Строение цветка представителей родов Orostachys и Rhodiola.
А. Orostachys paradoxa (рис. Т.В. Родновой): а – общий вид цветка; б, в, г, д, е – детали строения: б – лепестки, тычинки и нектарники, в – нектарник и плодолистик, г – листочки чашечки, д – пыльники, е – верхушка лепестка; Б. Rhodiola sp.: а – гермафродитный цветок, б – мужской цветок, в – женский цветок.
Лепестки в числе чашелистиков, свободные или сросшиеся в основании (Orostachys), как правило, длиннее чашелистиков (Hylotelephium, Aizopsis), редко гораздо короче (женские цветки Rhodiola), у представителей рода Pseudosedum и Sedum albertii венчик трубчатый или ворончатый, лепестки срастаются почти до 2/3 их длины. Лепестки желтые (Aizopsis, Sedum acre), зеленоватые или розоватые (Orostachys, Rhodiola), белые, розовые или малиновые (Hylotelephium, Pseudosedum, Sedum albertii). Для представителей родов Aizopsis, Hylotelephium, Orostachys и Rhodiola характерны более или менее четко выраженные «капюшон» и киль (Безделева, 1993; Бялт, 1999а).
Спайнолепестность венчика считалась важным диагностическим признаком еще с момента описания семейства (Candolle, 1801, 1828 а,b), на основании его разделялись не только роды, но и подсемейства. Однако современные исследования продемонстрировали искусственность подобного подхода (Mayzumi, Ohba, 2004), молекулярные и хемотаксономические данные показали, что признак мозаично распространен в монофилитичеcких группах (Hart, et al., 1999).
Нектарники (рис. 11, 12) Sedoideae подпестичные, представляют собой выросты рецептакула, расположенные в основании плодолистиков. Нектарники почти квадратные, прямоугольные или трапециевидные, сердцевидные; их число соответствует числу плодолистиков. Редко в мужских цветках Rhodiola нектарники отсутствуют вообще, либо число их сокращено до 1, очень редко встречаются нектарники, сросшиеся в трубку. H. Ohba (1975) указывал, что в женских цветках Rhodiola лепестки и нектарники очередные, а в мужских и гермафродитных нектарники супротивны лепесткам. У представителей родов Rhodiola и Pseudosedum образуются своеобразные «карманы» между нектарниками и плодолистиками, в которых скапливается выделяющийся нектар.
|
|
Некоторыми авторами морфология нектарников считается «хорошим» устойчивым признаком (Безделева, 1993; Бялт, 1999а), по крайней мере, на родовом или секционном уровне, например, представителям Hylotelephium и Orostachys sect. Orostachys subsect. Orostachys свойственны удлиненные узкие нектарники, а Aizopsis и Orostachys sect. Orostachys subsect. Appendiculata – широкие и короткие. Но эти признаки очень редко используются в диагностических целях, так как разнообразие их не велико.
Андроцей. Все изученные представители Sedoideae имеют обдиплостемонный андроцей – тычинки расположены в два круга, причем наружные тычинки супротивны лепесткам, а внутренние – с ними чередуются, к примеру некоторые Sedum s. str. имеют гаплостемонный (простой) андроцей, тычинки расположены в один круг. Редко, в результате редукции внешнего круга, у представителей Rhodiola встречаются цветки с одним кругом тычинок. Аранжировка тычинок с самого начала истории развития таксономии семейства считалась весомым признаком для разделения не только родов, но и подсемейств, например, все представители подсем. Crassuloideae имеют гаплостемонный андроцей (Berger, 1930).
Тычинки свободные или сросшиеся в основании с лепестками, как правило, тычинки, супротивные лепесткам, срастаются с ними (до 1/3 высоты), очередные лепесткам – свободные, у представителей Pseudosedum прирастают к трубке венчика. Обычно, тычинок в два раза больше, чем лепестков. Тычиночные нити тонкие нитевидные. Пыльники изученных Sedoideae базификсные, почти круглые или эллипсоидные, уплощенные с боков, темно-фиолетовые (почти черные), пурпурные, розовые, кремовые или желтые. Признаки морфологии тычинок имеют, как правило, небольшое таксономическое значение. Однако в отдельных случаях позволяют различить морфологически близкие виды, например, Orostachys malacophylla имеет желтые пыльники, а близкий к нему O. maximowiczii – малиновые, а единственный вид с дорзификсными пыльниками – Rhodiola hobsonii – выделен в монотипную секцию Hobsonia на основании этого признака (Ohba, 1975). Пыльца (рис. 13) изученных представителей Sedoideaе тризонокольпоратная (трехборозднопоровая). Вызревшие пыльцевые зерна широкоэллиптические или эллиптические, округлые на виде сверху и продолговато-сферические в районе экватора, слегка уплощенные, с бороздами, формирующими углы треугольника, тектум струйчато-морщинистый. Размер пыльцевых зерен варьирует от 15 до 25 мкм, самые крупные у представителей рода Hylotelephium – 15,5-25,5x14,0-20,0 мкм в среднем, Aizopsis – 19,5-25,1x14,5-22,1, Orostachys – 20,3-22,8x15,5-18,3. Наши исследования и результаты исследований других авторов продемонстрировали поразительное однообразие и стабильность признаков морфологии пыльцы (Бялт, 1999а; Hart, 1974; Eggli, 1988; Sin, et al., 2002). H. ‘t Hart (1974) отмечал, что пыльца Sedoideaе принадлежит к единому морфологическому типу и образует непрерывные морфологические ряды, что делает эти признаки малозначительными для таксономии подсемейства. Однако стабильность этих признаков подтверждает естественность семейства. Трехбороздные пыльцевые зерна – самый обычный тип пыльцы двудольных (Имс, 1964; Тахтаджян, 1964; Cronquist, 1970), характерен он и для Saxifragaceae, родственных Crassulaceae, однако представители семейства Saxifragaceae имеют в большинстве своем более сложную и разнообразную, чем у Crassulaceae, скульптуру поверхности (Ferguson, Webb, 1970; Kaplan, 1981). Особенности морфологии пыльцы Crassulaceae демонстрируют целый спектр приспособлений ко ксерическим условиям обитания: кольпии (борозды) – приспособления к изменению влажности (гармомегаты); поры – приспособления для ускоренного прорастания пыльцы; скульптурированная экзина – приспособление к опылению насекомыми (Punt, 1986; Thanikaimoni, 1986). |
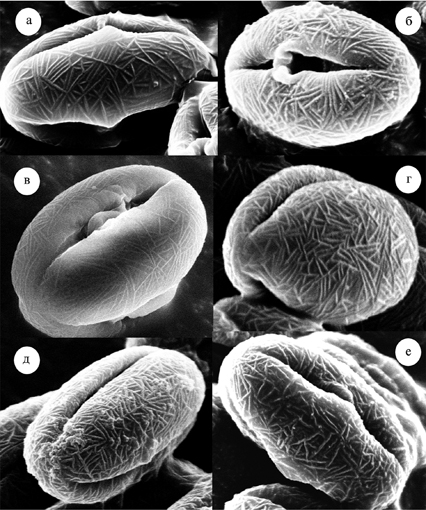
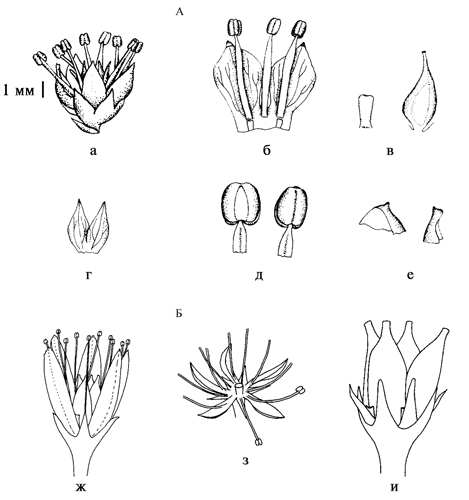
Рис. 13. Морфология пыльцевых зерен Sedoideae.
а – Orostachys spinosa, б – O. malacophylla, в – O. maximowiczii, г – Aizopsis middendorfianа subsp. middendorfiana, д – A. litoralis, е – Hylotelephium pallescens.
|
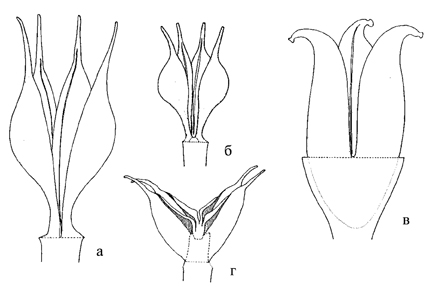
Рис. 14. Типы листовок Sedoideae (по: Ohba, 1978).
а – Orostachys malacophylla; б – Hylotelephium sieboldii; в – Rhodiola tibetica; г – Aizopsis aizoon; а, б, в – ортокарпные; г – киптокарпные.
|
Гинецей, плод. Плодолистики Sedoideae кондупликатные. Плацентация краевая или почти краевая, семязачатки располагаются вдоль брюшного (вентрального) шва. Общий тип плода толстянковых – циклическая многолистовка. Листовочки однолакунные, сросшиеся в основании (гемисинкарпные – Aizopsis, Rhodiola, Pseudosedum, Sedum acre), ножками (некоторые Orostachys) или полностью свободные (апокарпные, Hylotelephium). Плодики вскрываются по вентральному шву.
Зрелые плодолистики Sedum acre и Aizopsis имеют на брюшной стороне вывернутые края, формирующие своеобразные «губы». Завязь верхняя, только представители подрода Rhodiola имеют промежуточную (полунижнюю) завязь, из-за срастания трубки чашечки и оснований завязей (Ohba, 1977, 1978).
Многолистовки Sedoideae разделяют на два типа – киптокарпные и ортокарпные (рис. 14).
|
Киптокарпные характеризуются своеобразным «звездчатым» обликом и «горбатой» внешней стенкой плодолистика (Ohba, 1978; Grulich, 1984). Из изученных представителей такой тип гинецея имеют все представители Aizopsis и Sedum acre. Киптокарпные завязи часто одревесневают. В ортокарпном гинецее плодолистики прямые или немного отогнутые, внешняя стенка не выгнута. Ортокарпные листовки, как правило, имеют пленчатую или похожую на бумагу консистенцию (за исключением Rhodiola). Sedoideae отличается от близких подсемейств (например, Sempervivoideae) отсутствием вентральной полости в многолистовках (Ohba, 1978).
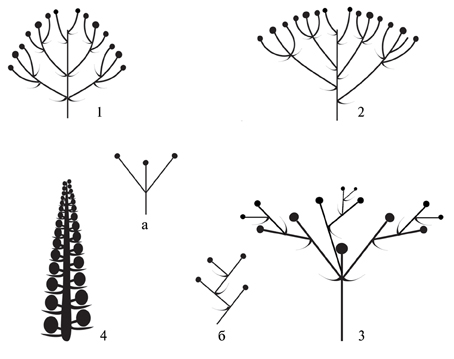
Рис. 15. Типы соцветий дальневосточных Sedoideae (схема).
1,2 – Плейотирс (1 – округлый, 2 – щитковидный) Hylotelephium; 3 – Плейохазий (щитковидный) Aizopsis; 4 – Колосовидное соцветие Orostachys; а,б – парциальные соцветия: а – дихазий, б – извилина.
|
Соцветия изученных Sedoideae верхушечные и в разной степени облиственные, закрытые, открытые или переходного типа; простые или сложные; много- или малоцветковые, иногда редуцированы до единственного цветка.
Цветки изученных Sedoideae собраны в соцветия трех основных типов [терминология и определения по: Федоров, Артюшенко, 1979; Weberling, 1989] (рис. 15):
1. Плейотирс (Hylotelephium, Rhodiola; рис. 15. 1, 2) – сложное соцветие, паракладии которого несут парциальные соцветия цимоидного характера, оно имеет удлиненную главную ось и большое количество разветвленных боковых ветвей, цимоиды располагаются на осях второго и более порядков. Парциальное соцветие – одинарный или двойной дихазий. По типу функционирования апикальных меристем тирсы очитковых относятся к переходному типу, верхушечный цветок верхушечного соцветия редуцирован, паракладии – закрытые. Плейотирсы могут быть щитковидными или округлыми. У представителей рода Hylotelephium соцветия могут насчитывать несколько сотен цветков и достигать диаметра 15-20 см, у представителей рода Rhodiola число цветков может быть редуцировано до одного.
2. Плейохазий (Aizopsis, Sedum, Pseudosedum; рис. 15. 3) – закрытое соцветие, несущее 3-5 (иногда более) равнозначных осей, расположенных почти горизонтально. Под цветком, завершающим главную ось соцветия, развиваются три и более парциальных соцветия. Парциальное соцветие – монохазиальное (извилина, завиток) такой тип соцветия характерен и некоторых представителей рода Rhodiola (Федоров, Артюшенко, 1979). Плейохазии, как и соцветия предыдущего типа, могут быть щитковидными или округлыми.
|
3. Колосовидное (початковидное; рис. 15. 4) открытое соцветие (Orostachys) имеет прямую удлиненную и утолщенную главную ось. Боковые оси короткие или отсутствуют вообще (если есть, то парциальное соцветие – дихазий или двойной дихазий). Соцветие, как правило, простое, редко (у O. maximowiczii) сложное канделябровидное, синфлоресценция может содержать до 50 паракладий.
Строение соцветий имеет существенное значение в систематике как семейства, так и подсемейства. На основании данного признака типы соцветий изученных очитковых четко дифференцируются, признак устойчив, и каких-либо вариаций в пределах вида не обнаружено. Каждый тип соцветия характеризует определенную таксономическую группу, так, к первому типу относятся виды родов Hylotelephium и Rhodiola, ко второму – Aizopsis, изученные виды Sedum, Pseudosedum и Rhodiola, к третьему – Orostachys.
По типу облиственности соцветий среди изученных очитковых можно выделить следующие группы: 1) фрондозно-фрондулезные – соцветие несет неизмененные ассимилирующие листья и листья меньших размеров, морфологически сходные с листьями срединной формации (представители рода Aizopsis – A. aizoon, A. litoralis, A. middendorfiana subsp. middendorfiana, A. middendorfiana subsp. sichotensis, A. kamtschatica); 2) фрондозно-брактеозные – у основания соцветия листья фрондулезные, у верхушки брактеозные, сильно видоизмененные, не сходные с листьями срединной формации (Pseudosedum, Sedum, Aizopsis kurilensis, A. selskiana, большинство представителей родов Hylotelephium и Orostachys); брактеозно-эбрактеозные – нижняя часть соцветия несет прицветники, в верхней части брактеи редуцированы (Hylotelephium erythrostictum, большинство Rhodiola).
Порядок зацветания в парциальных соцветиях типичен для цимозных – первым зацветает верхушечный цветок, затем – боковые. В общем плане в плейохазии цветки распускаются центробежно, в плейотирсе – центростремительно или акропетально, а колосовидном соцветии – акропетально.
Семя. Cеменная кожура у покрытосеменных возникает из интегументов семязачатка и является производной материнского организма (Эсау, 1980). Признаки семенной кожуры стабильны и консервативны, почти не поддаются воздействию внешней среды, что придает им высокое таксономическое значение (Barthlott, 1981, 1984).
В публикациях, касающихся Crassulaceae, описания семян обычно приводятся в качестве иллюстраций к морфологическим характеристикам видов. Первые подробные описания морфологии семян большого числа среднеевропейских Sedum s. l. сделаны H. Fröderström (1930-1935), признаки скульптуры поверхности были использованы автором при разделении видов по секциям и сериям. Он различал голые, гладкие, папиллезные и мамиллезные семена. R.T. Clausen (1984), изучив семена североамериканских видов толстянковых, различал голые, гладкие, ячеистые, сетчатые, бородавчатые, ребристые и с чешуйчатой скульптурой поверхности.
Использование методов электронной микроскопии для изучения архитектуры поверхности семян представителей Sedoideae показало разнообразие признаков морфологии семенной кожуры и их перспективность для систематики (Hart, Berendsen, 1980; Knapp, 1994).
H. 't Hart и W. Berendsen (1980) выделяли 3 типа орнаментации тесты семян Sedum s.l.: однобугорковый (unipapillate), двубугорковый (bipapillate) и многобугорковый (multipapillate) с подтипами и вариантами.
Четыре модели поверхности семян у представителей семейства выделял U. Knapp (1994): лестничная модель (Leitermodell или Kalanchoe-тип) с 7 подмоделями и 3 вариантами; сотовая модель (Wabenmodell или Echeveria-тип) с 5 подмоделями и 2 вариантами; мозаичная модель (Puzzle-Modell или Crassula-тип) также с 5 подмоделями и 2 вариантами; бородавчатая модель (Warzenmodell или Umbilicus-тип) с 2 вариантами.
Из изучаемых нами представителей Sedoideae до наших исследований (Абанькина, Гончарова, 1997; Гончарова, Абанькина, 1999; Колдаева, Гончарова, 2005; Gontcharova, 1999b) имелись данные о морфологии семян лишь для 7: Aizopsis selskiana (Hart, Berendsen, 1980), A aizoon, A. middendorfiana ssp. middendorfiana, Orostachys japonica, O. spinosa, Rhodiola quadrifida и Rh. rosea (Knapp, 1994). Краткие сведения о морфологии семян некоторых представителей рода Orostachys приведены в работе В.В. Бялта (1999а). Однако наши исследования не полностью согласуются с этими данными.
Анатомия семян Crassulaceae изучена довольно полно и обстоятельно (Данилова, 1996; Netolizky, 1926; Corner, 1976; Krach, 1976), но для толстянковых флоры российского Дальнего Востока нами найдены данные только о Aizopsis kamtschaticа (Данилова, 1996) и Rhodiola rosea (Бачаров и др., 2004).
Семена Crassulaceae и Sedoideae, в частности, имеют однотипное строение (Мандрик, Голышкин, 1973; Данилова, 1996; Гончарова, Абанькина, 1999; Бачаров и др., 2004; Колдаева, Гончарова, 2005; Netolizky, 1926; Corner, 1976; Krach, 1976; Knapp, 1994), они образуются из анатропных крассинуцелятных семязачатков и состоят из семенной кожуры, внезародышевых запасающих тканей и зародыша. Зародыш Sedoideae тотальный, семядольно-осевой, лопатовидный, содержит масло и белок (Krach, 1976).
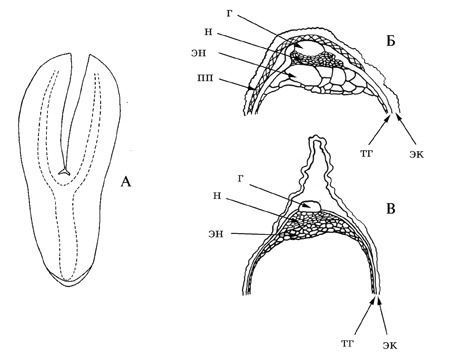
Рис. 16. Строение семени очитковых (по: Колдаева, Гончарова, 2005).
А – зародыш Aizopsis litoralis; Б, В – халазальная область семени (Б – Aizopsis aizoon; В – Hylotelephium ussuriense); тг – тегмен; н – нуцеллус; эк – экзотеста; эн – эндосперм; пп – проводящий пучек; г – гипостаза.
|
Семена изученных видов мелкие, яйцевидные, заостренно-яйцевидные, заостренно-эллиптические; опилковидные; темно-коричневые, коричневые, охристые. Окраска семенной кожуры обусловлена наличием танинов, пропитывающих все слои спермодермы. Микропиле расположено рядом с рубчиком. Микропилярная часть семени может быть округлой или заостренной или окружена «коронкой». Онтогенетические исследования показали, что присутствие «короны» – апоморфный специализированный признак, она появляется спустя неделю после оплодотворения, когда теста уже полностью сформирована (Hart, Berendsen, 1980). Рубчик небольшой, ланцетный или щелевидный, слегка вогнутый, либо не заметен. Рафе либо выражен четко и имеет вид узкого тонкого гребня, проходящего вдоль семени до халазы, либо не выражен. Халазальная часть семени может быть округлой или нести складчатые (Hylotelephium) или крыловидные (Rhodiola) образования, сложенные клетками интегументальной паренхимы. Такие выросты характерны для видов с самыми крупными семенами – Rhodiola alsia, Rh. ishidae, Rh. angusta. Sedoideae – автохоры, и, вероятно, разрастание семенной кожуры и образование крыловидных выростов является приспособлением для распространения семян при помощи ветра.
В области халазы у всех исследованных видов очитков хорошо выражена гипостаза, имеющая вид компактной группы темноокрашенных мелких клеток с утолщенными оболочками и признаками облитерации (рис. 16). Возможно, эта структура – результат развития гаустория, отсутствующего в зрелом семени (Бачаров и др., 2004; Krach, 1976).
Запасные питательные вещества, содержащиеся в целлюлярном эндосперме, практически полностью утилизируются в процессе развития зародыша еще до полного созревания семени, сохраняется, как правило, тонкий бесструктурный слой, окружающий зародыш и образующий «шапочку», покрывающую гипокотиль. В области гипостазы обнаруживаются остатки нуцеллуса.
|
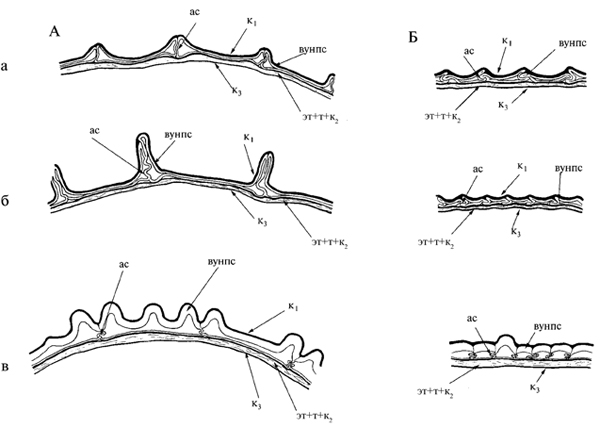
Рис.17. Строение покровов зрелого семени очитковых (по Колдаева, Гончарова, 2005).
А – в поперечном сечении; Б – в продольном сечении; а – Hylotelephium pallescens; б – H. ussuriense; в – Aizopsis selskiana; г – A. aizoon; д – A. litoralis; е – A. middendorfiana subsp. sichotensis; ас – антиклинальная стенка; вунпс – вторичное утолщение наружной периклинальной стенки; к1 – наружная кутикула, к3 – внутренняя кутикула, эк – экзотеста; эт+т+к2 – эндотеста + тегмен + промежуточная кутикула.
|
Зрелая семенная кожура изученных Sedoideae сложена из двух двухслойных покровов (рис. 17). Семена относятся к экзотестальному типу и укреплены по Reseda-типу, на внутренней периклинальной стенке клеток экзотесты образуется более или менее значительное вторичное утолщение. Такой тип утолщения характерен для семян большинства Crassulaceae, Saxifragaceae, Penthoraceae (Кульбаева, 1992; Немирович-Данченко, 1994; Krach, 1976). Укрепленная экзотеста вместе с толстой кутикулой создают механическую защиту. Клеточные стенки не лигнифицированы, состоят из целлюлозы с прослойками кутина и окрашиваются FeCl3 в темно-серый цвет (Цингер, 1958; Krach, 1976). Эндотеста, тегмен и расположенная между ними кутикула, как правило, плотно сжаты вместе и образуют единый бесструктурный темноокрашенный слой. Со стороны эндосперма и зародыша тегмен выстлан хорошо различимой кутикулой.
Различия в степени утолщения периклинальных и антиклинальных стенок, наличие папиловидных выростов и выпячиваний на наружной периклинальной стенке, толщина кутикулы и наличие эпикутикулярных образований обусловливают специфическую скульптуру поверхности семени очитковых.
|
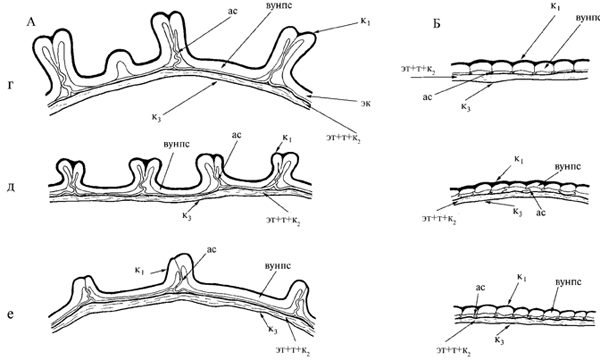
Продолжение рис. 17. г – A. aizoon; д – A. litoralis; е – A. middendorfiana subsp. sichotensis.
|
Экзотеста изученных представителей Sedoideae состоит из правильных продольных рядов поперечно удлиненных или почти изодиаметрических, четырех-, шестиугольных клеток, число рядов клеток не постоянно и варьирует у всех изученных видов. Клетки экзотесты однородны на большей части поверхности. Однако в области рафе клетки не имеют четко выраженной скульптуры и значительно отличаются размерами; клетки, примыкающие к рафе, могут быть изодиаметрическими, а покрывающие его – вытянутыми в меридиональном направлении. Покрывающая экзотесту кутикула часто маскирует скульптуру поверхности семени.
|
Краткая характеристика семян очитковых по родам и секциям
Род Aizopsis
Семена исследованных видов Aizopsis 1000-1500 мкм длиной, удлиненно-яйцевидной формы с заметно суженным микропилярным и округлым халазальным концом, слегка сплюснутые со спинки (рис. 18). Окраска семян темно-коричневая.
Стенки клеток экзотесты представителей рода Aizopsis в разной степени утолщены: наружная периклинальная (НПС) – значительно, внутренняя периклинальная – слабо. Антиклинальные стенки тоньше периклинальных, они сдавлены между последними и образуют S-образные складки (рис. 17). Дистальные антиклинальные стенки приподняты и вместе с прилегающим участком утолщенной НПС образуют 2 дистальных выроста. Проксимальные антиклинальные стенки меньшей высоты, чем дистальные, не приподнятые над поверхностью клетки. Поэтому НПС выглядит выпуклой на поперечном срезе клетки и вогнутой – на продольном.
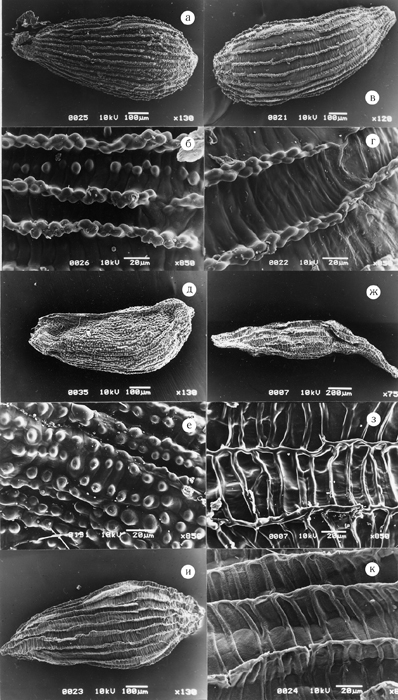
Рис. 18. Семена представителей родов Aizopsis и Hylotelephium.
а, б – Aizopsis litoralis; в, г – A. aizoon; д, е – A. selskiana; ж, з – Hylotelephium pallescens; и, к – H. ussuriense.
Клетки экзотесты некоторых видов рода Aizopsis имеют в центре НПС вырост (выпячивание) в виде бугорка.
Секция Aizopsis. Длина клеток экзотесты в три и более раз превышает ширину. Внешние периклинальные стенки слабовыпуклые. Периклинальные стенки клеток, расположенных около концов семени, могут нести 1-2 одиночно стоящих выпуклых выроста – папиллы (рис. 17). Утолщения дистальных антиклинальных стенок имеют полусферическую или цилиндрическую с округлой вершиной форму. Дистальные антиклинальные стенки в различной степени сомкнуты. В зависимости от степени смыкания антиклинальных стенок ребра могут быть волнистыми или почти ровными. Высота ребер от 25,6 мкм (А. aizoon) до 18,7 мкм (A. middendorfiana subsp. sichotensis). Степень смыкания дистальных антиклинальных стенок, степень их утолщения и высота видоспецифичны. Ребра располагаются почти параллельно от халазального конца семени к микропилярному. Секция Villosae. Длина клеток экзотесты менее чем в три раза превышает ширину. Периклинальные клетки несут один или несколько бугорков (папилл), одинаковых по форме и толщине с утолщениями дистальных антиклинальных клеток. Поверхность семян выглядит бугорчатой. Бугорки расположены продольными рядами, однако за счет бугорков, расположенных на периклинальных стенках, продольные ряды бугорков нечеткие. Вторичная скульптура поверхности не выражена либо слабовыражена, при большом увеличении выглядит слабоморщинистой.
Род Hylotelephium
Семена 900-2000 мкм длиной, ланцетные, уплощенные со спинки или удлиненно-ланцетные, почти цилиндрические в поперечном сечении. Ланцетная форма образуется за счет конусовидного складчатого выроста на халазальном или на обоих концах семени (рис. 18).
Окраска семян светло-коричневая с охристым оттенком, бледная желто-коричневая. Длина клеток экзотесты в три или более раз больше ширины.
Клетки экзотесты представителей рода Hylotelephium имеют равномерно и слабоутолщенные клеточные стенки (рис. 17). Антиклинальные стенки приподнятые, S-образно сдавленные. НПС погруженная, в результате чего она выглядит вогнутой как на продольном, так и на поперечном срезе экзотестальной клетки. В зависимости от особенностей морфологии антиклинальных клеток выделяются следующие типы: 1) дистальные и проксимальные антиклинальные стенки примерно равной высоты, поверхность семени выглядит сетчатой; 2) высота дистальных антиклинальных стенок в два и более раза превышает высоту проксимальных, либо проксимальные антиклинальные стенки клетки не выражены вовсе, дистальные стенки клеток плотно сомкнуты на всю высоту и формируют тонкие продольные ребра. |
|
Максимальная высота ребер отмечена нами для Н. ussuriense – 35,1 мкм, в среднем же высота ребер у представителей рода не превышает 10 мкм.
Вторичная скульптура поверхности спермодермы не выражена либо слабо выражена и при большом увеличении выглядит слабоморщинистой либо имеет вид мелких гранул и хлопьев, покрывающих всю поверхность эпидермы.
Род Orostachys
Семена мелкие 500-900 мкм длиной, удлиненно-яйцевидной или заостренно-яйцевидной формы с заметно суженным микропилярным и округлым халазальным концом, слегка сплюснутые со спинки (рис. 19); охристо-желтые, желтовато-коричневые, светло-коричневые.
Длина клеток экзотесты в 1,5-2 раза больше ширины. Строение клеток экзотесты и мофология поверхности в целом сходны с таковыми семян Hylotelephium с сетчатой скульптурой. Антиклинальные стенки более или менее равномерно утолщенные, приподнимающиеся и равной высоты. Антиклинальные стенки клеток смыкаются на всю высоту. Периклинальные стенки слабовогнутые. Поверхность семени ячеисто-сетчатая.
Вторичная скульптура мелкогранулированная.
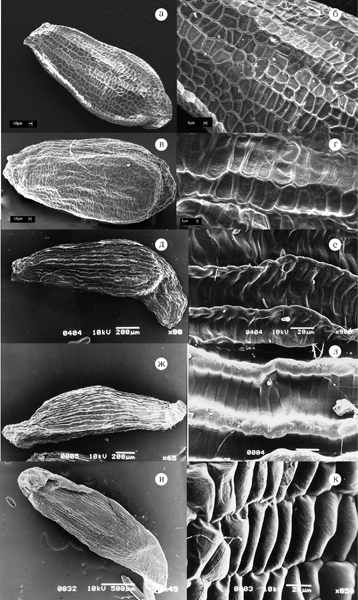
Рис. 19. Семена представителей родов Orostachys и Rhodiola.
а, б – Orostachys malacophylla; в, г – O. japonica; д, е – Rhodiola integrifolia; ж, з – Rh. rosea subsp. rosea; и, к – Rh. ishidae.
Род Rhodiola
Семена до 3000 мкм длиной, ланцетные, уплощенные со спинки, удлиненно-ланцетные, почти цилиндрические в поперечном сечении. На халазальном и микропилярном концах семени либо только на халазальном конце образуются более или менее выраженные конусовидные складчатые или крыловидные выросты (рис. 19). Окраска темно-коричневая. У большинства видов клетки вытянуты перпендикулярно морфологической оси семени, длина клеток более чем в три раза превышает ширину. Редко клетки почти изодиаметрические (Rh. angusta, Rh. ishidae). Наружные периклинальные стенки клеток экзотесты плоские, слабовогнутые либо в различной степени выпуклые.
|
|
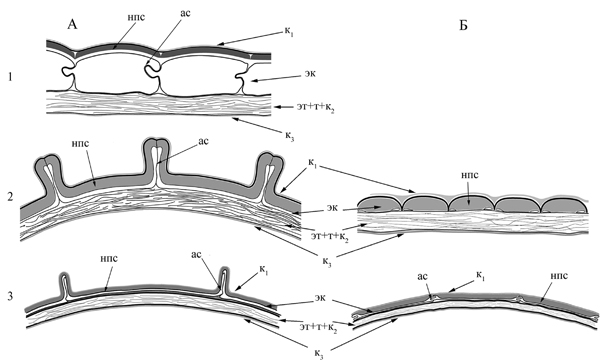
Рис.20. Строение покровов зрелого семени Rhodiola.
А – в поперечном сечении; Б – в продольном сечении; 1 – Rhodiola rosea; 2 – Rh. integrifolia; 3 – Rh. ishidae; ас – антиклинальная стенка; нпс – наружная периклинальная стенка; к1 – наружная кутикула, к3 – внутренняя кутикула, эк – экзотеста; эт+т+к2 – эндотеста + тегмен + промежуточная кутикула.
|
Стенки могут быть более или менее равномерно утолщены (Rh. integrifolia, Rh. rosea subsp. sachalinensis, R. rosea subsp. arctica, Rh. rosea subsp. rosea), или наружная периклинальная стенка за счет мощного вторичного утолщения значительно толще как внутренней периклинальной, так и антиклинальных (Rh. alsia, Rh. ishidae, Rh. angusta, Rh. rosea subsp. rosea) (рис. 20).
Антиклинальные стенки, как правило, тонкие, приподнятые, S-образно изогнуты и образуют складки, создающие видимость утолщения. У семян с сильно утолщенной наружной стенкой экзотесты антиклинальные стенки не выступают над поверхностью клетки, формируя бугорчатый тип поверхности, в том случае, если все стенки более или менее равномерно утолщены, складки антиклинальных стенок часто значительно приподнимаются над поверхностью, а периклинальная стенка проваливается внутрь, формируя ребристую или сетчатую скульптуру семенной кожуры.
Вторичная скульптура мелкогранулированная, слабоморщинистая или не выражена.
|
Таким образом, в зависимости от комбинации признаков мы выделяем следующие типы поверхности семян Sedoideae:
- 1. Поверхность продольноребристая (longitudinally costate) – дистальные антиклинальные клеточные стенки хорошо выраженные, приподнятые и всегда выше проксимальных антиклинальных стенок.
- В пределах данного типа различаются два подтипа:
- 1.1. Поверхность широкоребристая (laticostate) – наружные периклинальные стенки со значительным вторичным утолщением, дистальные антиклинальные стенки значительно утолщены, с округлыми или слегка уплощенными верхушками (Aizopsis aizoon, A. litoralis, A. middendorfiana ssp. sichotensis, Rhodiola rosea subsp. rosea), дистальные антиклинальные стенки в разной степени сомкнутые, что обусловливает большую или меньшую волнистость ребер.
- 1.2. Поверхность узкоребристая (tenuicostate) – наружные периклинальные стенки с незначительным вторичным утолщением, дистальные антиклинальные стенки неутолщенные, полностью сомкнуты (Hylotelephium ussuriense, Rhodiola integrifolia, Rh. rosea subsp. sachalinensis, Rh. rosea subsp. rosea).
- 2. Поверхность сетчатая (reticulate) – клеточные стенки с незначительным, равномерным вторичным утолщением, дистальные и проксимальные антиклинальные стенки приподнятые, равной высоты и толщины, полностью сомкнутые (представители родов Hylotelephium, Orostachys).
- 3. Поверхность многобугорковая (multipapillate) – клеточные стенки со значительным, равномерным вторичным утолщением, периклинальные стенки несут два бугорка (выпячивания), расположенных около дистальных концов клетки. Иногда дополнительные бугорки расположены в центре клетки (Aizopsis selskiana).
- 4. Поверхность бугорчатая (colliculate) – периклинальные стенки со значительным вторичным утолщением, выгнутые, антиклинальные стенки не подняты над поверхностью. Среди представителей Sedoideae подобный тип поверхности спермодермы встречается только в роде Rhodiola (Rh. alsia, Rh. angusta, Rh. ishidae).
Признаки макроскульптуры семенной кожуры стабильны у большинства исследованных образцов. Однако не всегда эти признаки могут быть использованы для дифференциации видов, так как, например, Rh. alsia, Rh. angusta, Rh. ishidae не имеют четких видовых различий по строению семенной кожуры, хотя хорошо различаются по иным признакам.
Признаки семян исследованных образцов Rh. rosea subsp. rosea значительно варьируют. Морфологические характеристики эпидермы слабо реагируют на условия окружающей среды, в которых растение росло (Barthlott, 1981), поэтому разнообразие условий местообитаний, из которых собраны образцы, широкий ареал вида не могут объяснить такое варьирование. На наш взгляд, разнообразие признаков скульптуры семенной кожуры в пределах одного вида может означать либо сборную природу этого таксона, либо неверное определение некоторых образцов. Нами не выявлено четкой взаимосвязи морфологии поверхности семян Rh. rosea subsp. rosea и ее распространения, однако можно отметить, что в европейской части ареала преобладают образцы с широкоребристой поверхностью, азиатские же характеризуются всеми обнаруженными нами типами скульптуры семенной кожуры, но количественно преобладают образцы с узкоребристым типом поверхности.
Rh. integrifolia – один из сложных в систематическом отношении видов, рассматриваемый разными авторами либо как самостоятельный, либо как синоним Rh. rosea subsp. rosea (Ohba, 1981). Однако наши исследования показали стабильность признаков морфологии семян Rh. integrifolia. Хотя семена названного вида и были отнесены к тому же подтипу, что и некоторые образцы семян Rh. rosea subsp. rosea, сочетание признаков скульптуры поверхности семян четко отличает их.
Признаки вторичной скульптуры варьируют у всех изученных видов, наиболее часто встречаются семена с почти гладкой кутикулой, реже – с морщинистой или мелкобугорчатой. Как правило, наиболее скульптурированные эпидермальные поверхности имеют растения, обитающие в более сухих условиях, растения из более влажных мест произрастания часто имеют слабоскульптурированные или гладкие поверхности (Barthlott, 1981).
Изучение морфологии поверхности семян Sedoideae показало, что таксоны рангов секции и серии, как правило, имеют специфичные наборы признаков семенной кожуры (Hart, Berendsen, 1980; Knapp, 1994; Gontcharova, 1999b). Внутри подрода Rhodiola эти признаки варьируют весьма значительно, четко выделяются две группы видов: Rh. angusta, Rh. alsia, Rh. ishidae – с бугорчатой поверхностью и Rh. integrifolia, Rh. rosea subsp. sachalinensis, Rh. rosea subsp. rosea – с ребристыми типами поверхности. На наш взгляд, широкая вариабельность признаков внутри секции отражает сборную природу этого таксона.
H. Ohba (1981), проанализировав комплекс морфологических признаков восточно-азиатских представителей рода Rhodiola, сделал вывод о близком родстве Rh. angusta, Rh. alsia, Rh. ishidae. Наши данные о морфологии семян подтверждают это предположение. Бугорчатая скульптура семенной кожуры, наличие крыловидного выроста выделяют названые виды среди других представителей подрода, что подтверждает верность выделения их в секцию Algida (см. гл. 5).
Выявленные различия в строении семян и скульптуре семенной кожуры Hylotelephium ussuriense и H. pallescens, H. triphyllum типичных представителей серии Hylotelephium подтвердили правильность переноса H. ussuriense из серии Hylotelephium в серию Sieboldia H. Ohba, сделанного нами ранее на основании признаков вегетативной сферы (Gontcharova, 1999a).
Секции Aizopsis (A. aizoon, A. litoralis, A. middendorfiana subsp. sichotensis) и Villosae (A. selskiana) рода Aizopsis характеризуются специфичными наборами морфологических признаков семян, что также подтверждает обоснованность их разделения, предложенного нами ранее (Gontcharova, 1999a).
Полученные нами данные о строении поверхности семени A. selskiana идентичны результатам H. ‘t Hart и W. Berendsen (1980), что еще раз демонстрирует стабильность признаков скульптуры тесты и отсутствие заметного влияния природно-климатических условий произрастания вида на последние.
4.2. Вегетативные органы
Листья более или менее суккулентные, от почти плоских (Aizopsis), до вальковатых, почти круглых в поперечном сечении (Orostachys japonica, Sedum acre, S. albertii). От округлых до узколанцетных, почти линейных. От цельнокрайних до мелкозубчатых. Листья представителей рода Orostachys секции Orostachys подсекции Appendiculatae имеют шиповидные, хрящеватые и бахромчатые придатки.
Характерным отличием анатомического строения листьев суккулентов вообще и толстянковых в частности считается слабая дифференциация мезофилла (или ее полное отсутствие) на столбчатый и губчатый (Борисовская, 1960; Eggli, 1988), что характерно и для большинства изученных нами Sedoideae, обладающих выраженным суккулентным обликом. Однако мезофилл листьев очитковых из более или менее мезофитных местообитаний может быть в большей или меньшей степени дифференциацирован на столбчатую и губчатую паренхиму и иметь строение близкое к типичному – дорзовентральному. Специализированные водоносные ткани, как правило, отсутствуют, вся паренхима выполняет водозапасающую функцию, однако по данным И. Далькэ и др. (2006) листья Sedum acre имеют водозапасающий слой. Клетки мезофилла крупные, тонкостенные, заполнены мелкими хлоропластами. Межклетники могут быть хорошо выражены (Orostachys), или клетки мезофилла плотно прилегают друг к другу (Aizopsis, Hylotelephium). Клетки эпидермы крупные, с кутинизированными стенками, на поверхности часто откладывается более или менее значительный слой воска, определяющий сизую окраску листьев. Антоциановая окраска, характерная для многих видов, определяется окраской отдельных содержащих пигменты клеток. В эпидерме и мезофилле встречаются клетки, содержащие слизь, слизи могут содержаться и в межклетниках (Бялт, 1999а). Листья изученных очитковых, как обычно для всех толстянковых, несут устьица на обеих поверхностях листа, для них характерен анизоцитный устьичный аппарат с тремя околоустьичными клетками разных размеров, однако Н. Ohba (1990) указывает для представителей рода Orostachys секции Orostachys устьица с двумя околоустьичными (побочными) клетками. Морфогенез листа типичен для ксерофитов. Рост листа происходит главным образом за счет деления клеток в его основании, что типично для почечных чешуй (Борисовская, 1960).
Диагностическим признаком родов Rhodiola и Pseudosedum является наличие метаморфизированных чешуевидных листьев на корневищах и каудексах.
Листорасположение (филлотаксис) характеризуется числом листьев в узле и смещением листьев по окружности стебля, выраженным в градусах (Лотова, 2001).
Изученные толстянковые отличаются значительным разнообразием типов листорасположения. Преобладающим типом, как и вообще среди цветковых растений, является очередное (спиральное или рассеянное) листорасположение, при котором от каждого узла побега отходит один лист. В зависимости от угла расхождения (рассеяния или дивергенции) расположение может быть трех-, четырех-, пяти- или многорядным. Угол расхождения листьев некоторых видов Orostachys с листовой формулой 3-5/13 (138о27') может почти достигать «идеального» значения (137о30'29”), при котором достигается максимально эффективное использование доступной световой энергии. Такие листовые формулы довольно типичны для розеточных представителей Crassulaceae, представителей родов Sempervivum, Aeonium, Rosularia и др.
При супротивном листорасположении из узла отходят два листа, угол дивергенции в узле равен 180о. У толстянковых с таким типом филлотаксиса пары листьев расположены под прямым углом относительно друг друга (накрест-супротивно), листья располагаются в четыре ряда на побеге, общий угол дивергенции равен 90о (Hylotelephium ussuriense, H. ewersii, Orostachys paradoxa).
При мутовчатом листорасположении из узла отходят три листа и более. Угол дивергенции равен 120-90-72о в узле. Такой тип довольно обычен у дальневосточных очитковых (Hylotelephium verticillatum, H. viviparum).
Среди изученных представителей наблюдаются и переходные типы, при которых в пределах побега можно обнаружить изменение листорасположения – от мутовчатого (точнее, ложномутовчатого) до очередного (Hylotelephium erythrostictum, H. pallescens) либо от супротивного до очередного (Aizopsis litoralis, Hylotelephium pluricaule).
Структурно-морфологический тип побегов определяется длиной междоузлий и направлением роста побега. Большинство изученных очитковых имеют удлиненные побеги с более или менее равномерно расположенными ассимилирующими и иногда чешуевидными (Rhodiola, Pseudosedum) листьями и хорошо развитыми междоузлиями. Обычны также полурозеточные побеги, имеющие на первых этапах развития укороченные междоузлия, а при переходе в генеративный период образующие удлиненный побег (Orostachys). Среди восточно-азиатских Sedoideae облигатно розеточные побеги (брахибласты) встречаются только у Meterostachys, но они весьма обычны у тропических и субтропических Crassulaceae (Aeonium, Sempervivum). H. Ohba (1978) выделил две категории розеток: розетки, образованные прикорневыми листьями (некоторые Rhodiola, подрод Primuloides, Rosularia и Meterostachys), и розетки, сформированные базальными листьями цветоносных побегов (Orostachys). Растения имеют различные типы побегообразования, в первом случае это – моноподиально розеточные растения со специализированными генеративными побегами, соцветия развиваются из пазушных почек, во втором – симподиально полурозеточные, соцветия развиваются из верхушечной почки. Позже U. Eggli (1988) выдел третий тип розеток, характерный для видов с розеточными пропагулами, обычными в родах Rosularia и Sempervivum. Розетки образуются из пазушных почек, а соцветие – из верхушечной. Розетки у представителей рода Orostachys могут быть либо почти сидячими на коротких неразветвленных побегах (Orostachys aggregata), на длинных разветвленных корневищах (O. paradoxa) или столонах (O. furusei). Розетки изученных очитковых – приземные, однако среди субтропических и тропических толстянковых обычны и надземные (Aeonium, Rosularia).
На наш взгляд, разделение на полурозеточные и безрозеточные Sedoideae весьма условно, так как у хамефитов в процессе морфогенеза побег проходит фазу «розетки» – окрытой почки возобновления, несущей ассимилирующие видоизмененные листья (например, Hylotelephium pluricaule). Такие почки гомологичны, на наш взгляд, розеткам Orostachys, кроме того, у большинства горноколосников во время цветения розетка вытягивается, побег выглядит удлиненным. Главное различие между полурозеточными и безрозеточными побегами толстянковых, по нашему мнению, заключается в продолжительности «розеточной» стадии, у первых розетки сохраняются до начала бутонизации, у вторых – это покоящаяся стадия, побеги начинают активный рост сразу после наступления благоприятного периода.
Побеги изученных очитковых ортотропные (Hylotelephium pallescens, Rhodiola rosea, Aizopsis aizoon и др.) или анизотропные (Hylotelephium ussuriense, Aizopsis middendorfiana и др.). Признак может быть стабильным или значительно варьирует. Например, Hylotelephium pallescens имеет исключительно ортотропные побеги, H. ussuriense – только анизотропные, а Aizopsis selskiana – как ортотропные, так и анизотропные.
Надземные части моноподиальных побегов могут отмирать полностью (у криптофитов и гемикриптофитов), частично или редко сохраняться (у хамефитов). Многолетние надземные части побегов хамефитов могут либо одревесневать (Aizopsis middendorfianа и др.), либо не одревесневать (виды Orostachys, Sedum acre). Однако вторичное одревеснение монокарпических однолетних побегов имеет место и у криптофитов (Hylotelephium triphyllum и др.) (Hart, Koek-Noorman, 1989). Монокарпический побег при полегании обнаруживает способность к укоренению в каждом узле вплоть до соцветия. Плагиотропные части побегов часто укореняются, входя в состав эпигеогенного корневища.
У большинства видов все монокарпические побеги текущего года цветут и плодоносят, однако Pseudosedum lievenii, Sedum acre, S. albertii и Aizopsis hybrida характерен диморфизм побегов. На одном растении имеются как генеративные, так и более или менее многочисленные вегетативные (стерильные побеги). Как правило, на таких побегах листья расположены более плотно, чем на генеративных, а у А. hybrida – на верхушке побега образуют розетку.
Анатомия стеблей. Наши исследования (Гончарова, Царенко, 2004) показали (рис. 21), что стебли хорошо развитых однолетних генеративных побегов на большей их части покрыты эпидермой с сильно утолщенными кутинизированными наружными клеточными стенками. Характерным признаком всех изученных видов является хорошо развитая в стебле кора, почти целиком состоящая из крупных паренхимных клеток, что типично для суккулентов. В периферической части первичной коры почти у всех изученных видов имеется 1-3 слоя клеток пластинчатой колленхимы, образующей непрерывный цилиндр. Число слоев клеток коровой паренхимы варьирует от 6-8 (Aizopsis middendorfiana ssp. middendorfianа) до 28-31 (Orostachys japonica).
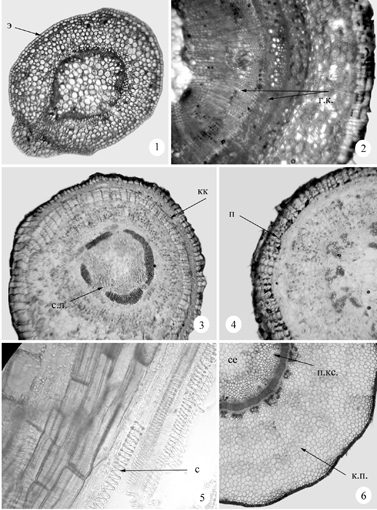
Рис. 21. Анатомия стебля некоторых дальневосточных представителей Sedoideae. 1 – стебель проростка Aizopsis middendorfianа, 2 – многолетний стебель A. middendorfianа; 3 – Hylotelephium pluricaule; 4 – Orostachys paradoxa; 5, 6 – Orostachys japonica; в.л. – волокна либриформа, в.кс. – вторичная ксилема, г.к. – годичные кольца, к – колленхима, ка – камбий, кк – корка, к.п. – коровая паренхима, м.в. – многоклеточные волоски, п – перидерма, п.кс. – первичная ксилема, с – сосуды, се – сердцевина, с.к. – слизевые клетки, с.л. – сердцевинные лучи, ф – флоэма, э – эпидерма.
|
Продолжение рис. 21. 7 – Aizopsis aizoon; 8 – опушение Aizopsis selskiana; 9, 10 – Hylotelephium ussuriense; 11, 12 – Aizopsis selskiana.
|
Клетки коровой паренхимы заполнены зернами крахмала (особенно обильного в конце вегетационного периода), дубильными веществами и слизью. У Orostachys japonica в коровой паренхиме присутствуют млечники, найденные ранее только в корневищах Rhodiola coccinea (Хоружая, 1976). У Aizopsis middendorfiana ssp. middendorfianа внутренние слои коровой паренхимы одревесневают. В основании однолетних отмирающих генеративных побегов изученных Sedoideae эпидерма сменяется перидермой. Многолетние неотмирающие части надземных побегов покрыты 7-11 слоями перидермы. В коре Aizopsis middendorfianа ssp. middendorfianа и Hylotelephium ussuriense нами обнаружены чечевички.
Многолетние стебли Hylotelephium pluricaule покрыты коркой (рис. 21. 3), образование которой связано с неоднократным заложением фелогена (Лотова, 2001).
Одним из характерных признаков анатомического строения стеблей Sedoideae, ранее отмеченным М.Г. Борисовской (1960), является слабое развитие флоэмы и ксилемы. Клетки флоэмы очень мелкие. Флоэма рано стареет вследствие перехода в генеративную фазу. Граница между ксилемой и сердцевиной неровная из-за того, что тяжи первичной ксилемы вдаются в сердцевину. Сердцевина состоит из тонкостенных паренхимных клеток. К концу вегетационного периода стебли однолетних побегов становятся полыми.
Сосуды первичной ксилемы имеют кольчатые и спиральные утолщения.
У большинства изученных видов вторичная ксилема непучкового типа, образует компактный цилиндр и представлена сосудами, волокнистыми трахеидами, волокнами либриформа и скудной тонкостенной паратрахеальной паренхимой. Сосуды округлые, собранные в небольшие группы или узкие цепочки, погружены в волокнистые трахеиды или волокна либриформа. На ранних этапах развития сосуды вторичной ксилемы узкие (значительно уже сосудов первичной ксилемы), на поперечных срезах они почти не различимы среди волокнистых элементов и паренхимы, на более поздних – формируются сосуды с более широкими просветами. Перфорации сосудов простые, вторичные утолщения члеников сосудов кольчатые, спиральные (рис. 21. 5, 10), очень редко лестничные или сетчато-лестничные. Волокна несептированные, с многочисленными порами. Во вторичной ксилеме всех изученных видов нами не обнаружены лучи, отсутствие вторичных лучей отмечалось и всеми предшествующими исследователями у других представителей Crassulaceae (Hart, Koek-Noorman, 1989; Hart, 1994). Стебли Orostachys paradoxa имеют пучковое строение вторичной ксилемы (рис. 21. 4). Пучки разного размера (ксилема состоит их сосудов с кольчатыми и спиральными утолщениями и малым количеством волокон либриформа) разделены более или менее широкими сердцевинными лучами. Ширина первичных лучей варьирует в зависимости от условий произрастания и возраста растения. Так, экземпляры Orostachys paradoxa из естественных местообитаний содержат существенно большее количество паренхимы, чем экземпляры, выращенные в оранжерее. В старых частях многолетних надземных стеблей ксилема сильно паренхиматизирована. До наших исследований сердцевинные лучи указывались для одно, двулетников из sect. Epeteium Boiss. рода Sedum (Борисовская, 1960) и Kalanchoe (Jensen, цит. по: Hart, Koek-Noorman, 1989).
В стеблях однолетних цветоносных побегов образование волокон соответствует конечной стадии вторичного роста и началу отмирания побега, однако в базальных частях стеблей хамефитов вторичный рост продолжается несколько лет, у полукустарничковых хамефитов образуются годичные кольца.
Полученные нами результаты в целом согласуются с исследованиями предшествующих авторов (Борисовская, 1960; Hart, Koek-Noorman, 1989; Hart, 1994). Следует отметить, что анатомическое строение стебля Crassulaceae не типично для обычного строения древесины. Основные отличия строения ксилемы толстянковых – короткие сосудистые элементы, отсутствие вторичных лучей и наличие кольчатых и спиральных утолщений трахеальных элементов вторичной ксилемы.
По мнению К. Эзау (1980), в элементах с более мощным, чем спиральное, утолщением вторичная оболочка откладывается в два этапа. Первым строится спиральный остов (вторичная оболочка первого порядка), затем между кольцами спирали откладывается дополнительная порция вторичной оболочки в виде пластин или тяжей. Вероятно, у представителей Crassulaceae подавлено образование вторичной оболочки второго порядка.
Первичные и вторичные трахеальные элементы в типичных случаях имеют не только различное строение, но и различные функции. Первичная ксилема, в отличие от вторичной, не только проводит водные растворы, но и перераспределяет их между окружающими тканями (Гамалей, 1972). В связи с анатомическими и физиологическими особенностями суккулентов, как крупноклеточная сердцевинная паренхима, так и коровая паренхима выполняют водозапасающую функцию (Борисовская, 1960), ксилема же, сохранившая признаки первичного строения, имеет значительно большую поверхность водопроводящих участков оболочки, что способствует более эффективному перераспределению и использованию воды. A. Berger (1930), S. Carlquist (1962), H. ‘t Hart и J. Koek-Noorman (1989) считали кольчатое или спиральное вторичное утолщение побегов представителей Crassulaceae приспособлением скорее к дефициту воды, чем к высокой температуре. Изученные нами виды распространены главным образом в умеренных районах, чаще всего в мезофитных местообитаниях, однако они сохраняют анатомическую структуру типичную для тропических представителей из ксерических местообитаний.
Хотя признаки анатомического строения не имеют существенного значения для систематики внутри как семейства, так и подсемейства, но их единообразие и специфика еще раз подтверждают, что Crassulaceae – естественная группа.
Длительность жизни. Наиболее многочисленны среди изученных Sedoideae поликарпические многолетние виды с однолетними генеративными побегами (все представители родов Rhodiola, Hylotelephium, большинство Aizopsis, Pseudosedum). Редко полурозеточные побеги Orostachys при отсутствии цветения нарастают моноподиально и становятся полициклическими. Встречаются настоящие монокарпики – двулетники (Orostachys maximowiczii), поликарпические многолетние растения с монокарпическими двулетними розетками (O. aggregata, O. furusei), а также виды, которые в зависимости от условий произрастания могут проявлять себя либо как монокарпики, либо как поликарпики (O. malacophylla, O. spinosa, O. japonica) (Гончарова, 1996а). Как правило, в описаниях видов рода Orostachys указывается, что его представители – монокарпические одно-, двулетние суккулентные травы. Но, как показали наши исследования, различные виды горноколосников проявляют различную степень поликарпичности. Так, O. maximowiczii – типичный двулетник, очень редко образует дочерние вегетативные розетки. О. fimbriata, O. japonica, О. malacophylla, O. spinosa, О. thyrsiflora в равной степени могут развиваться как по первому, так и по второму типу онтогенеза. O. aggregata и O. furussei, О. paradoxa – облигатные поликарпики, формирующие вегетативные розетки на столоновидных неотмирающих побегах, образуя более или менее плотную дерновину.
Нами была проведена серия опытов по стимуляции вегетативного возобновления у монокарпических экземпляров. Результаты показали, что полное или частичное искусственное разрушение генеративной почки приводит к массовому образованию вегетативных розеток, и, следовательно, значительно продлевает жизнь растению. По-видимому, и в естественных условиях губительное влияние некоторых факторов внешней среды (низких температур) на нормальное развитие генеративной сферы может вызывать подобное явление.
Возобновление системы побегов очитковых, как правило, симподиальное. После плодоношения большая часть побега отмирает, из почек возобновления, образующихся в основаниях монокарпических генеративных побегов, развиваются новые побеги (Sedum acre, S. albertii, Pseudosedum lievenii, все виды Hylotelephium, Aizopsis, за исключением A. selskiana).
Моноподиальное возобновление характерно как для монокарпических видов (Orostachys), так и поликарпических (Rhodiola). Orostachys spinosa – многолетник, у которого в течение нескольких лет наблюдается моноподиальный рост вегетативных розеточных побегов и только затем формируется удлиненный генеративный побег (Безделева, 1995). У представителей рода Rhodiola многолетние корневища нарастают моноподиально, боковые цветоносные побеги формируются из почек, образующихся в пазухах чешуевидных либо побеговых листьев (Ohba, 1978).
Смешанное симподиально-моноподиальное возобновление имеет Aizopsis selskiana – в течение ряда лет у молодых генеративных растений постоянно функционирует терминальная почка, возобновление происходит подобно Rhodiola, однако имеет место и симподиальное возобновление, при достижении зрелого генеративного возраста последнее преобладает над моноподиальным.
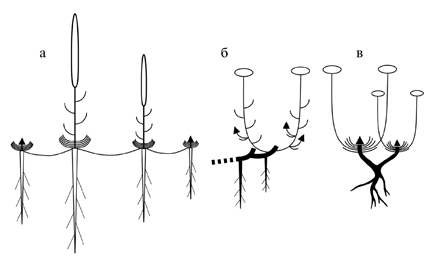
Рис. 22. Модели побегообразования дальневосточных Sedoideae.
а – симподиально-полурозеточная; б – симподиально диннопобеговая; в – моноподиально розеточная.
|
Модели побегообразования (рис. 22) определяются сочетанием структурно-морфологического типа побега и типа его возобновления.
Наиболее обычны среди изученных очитковых виды с длиннопобеговой симподиальной моделью (рис. 22. б), в этом случае растение строится из однотипных, повторяющих друг друга структурных единиц (метамеров различного порядка) – многолетних участков (резидов) и закрытых удлиненных монокарпических моноциклических побегов (Sedum acre, S. albertii, Pseudosedum lievenii, все Hylotelephium и Aizopsis). Реже встречается симподиально полурозеточная модель (многолетние Orostachys, рис. 22. а).
При моноподиальной модели (рис. 22. в) побеговая система построена из повторяющихся элементов структуры двух типов – открытого полициклического и закрытого монокарпического побега (Серебрякова, 1977, 1981; Жмылев и др., 1993, 2002). Среди изученных Sedoideae представители Rhodiola имеют моноподиально-розеточную модель побегообразования с открытой верхушечной почкой (брахиморфной структурой) и удлиненными монокарпическими побегами.
|
Почки возобновления большинства изученных Sedoideae открытые (голые), однако у некоторых представителей Rhodiola [sect. Chamaerhodiola (Ohba, 1978)] – закрытые, защищенные тонкими пленчатыми чешуями. Открытые почки двух типов: 1) розетковидные, большая часть зачаточных фотосинтезирующих листьев которых расположена под значительным углом к оси, часто перпендикулярно, только самые верхние листья смыкаются, прикрывая апекс, такие почки характерны для хамефитов и полурозеточных гемикриптофитов; 2) головчатые, листья плотно прижаты к оси, большинство из них смыкается над апексом, образуя защитный покров, характерны для гемикриптофитов и криптофитов.
Почки располагаются либо в основаниях годичных побегов (Sedum acre, S. albertii, Pseudosedum lievenii, Hylotelephium, Aizopsis), либо на корневищах или каудексах (виды Rhodiola). У некоторых представителей Orostachys – на коротких (O. aggregata) или длинных (O. furusei) столонах. По положению почек возобновления относительно субстрата четко выделяются 3 группы видов: криптофиты (Hylotelephium triphyllum, Aizopsis aizoon и др.), гемикриптофиты (Aizopsis selskiana, виды Rhodiola) и хамефиты (виды Orostachys, Hylotelephium pluricaule, Aizopsis kamtschatica и др.).
Кроме почек возобновления, некоторые виды изученных очитковых несут специализированные почки, выполняющие функции органов вегетативного размножения, о них будет сказано ниже (гл. 6).
Подземные органы изученных Sedoideae представлены гетероризными (аллоризными, морфологически неоднородными) и вторично гоморизными (морфологически однородными) корневыми системами, корневищами, каудексами и метаморфизированными запасающими корнями. Гетероризные корневые системы представлены: стержне-кистекорневыми (первично гетероризные – одно-, двулетние Orostachys, Pseudosedum lievenii; вторично гетероризные – многолетние Orostachys), у которых система главного корня наряду с придаточными корнями функционирует в течение большей части жизни; каудексами (Aizopsis selskiana, Rhodiola rosea), образованными нижними, многолетними, одревесневающими участками побегов, несущими почки возобновления, и главным корнем. Главный корень у Rhodiola rosea может достигать 1 м в длину и 3 см в диаметре (Некратова и др., 1992). От корневищ каудексы отличаются способом отмирания (Васильев и др., 1988).
Принципиальны и анатомические различия. Так, каудекс Rhodiola rosea, в отличие от симподиальных корневищ представителей родов Hylotelephium и Aizopsis, имеет два концентрических слоя пучкового камбия, причем внутренний – «инвертированый» (Хныкина, Зотова, 1966). Такой «инвертированный» камбий продуцирует флоэму по направлению к центру, а ксилему – к периферии органа. По утверждению H. ‘t Hart (1994), наружный камбий является продолжением пучкового камбия первичной стелы, а внутренний происходит из тканей аксилярных олиственных побегов. Интересно отметить, что побеги имеют проводяющую систему непучкового типа, а каудексы – пучкового.
Преобладающим типом корневых систем изученных Sedoideae являются вторично гоморизные корневые системы, характерные и для корневищных растений. Главный корень еще при прорастании семени слабо различим в массе придаточных корней, система главного корня отмирает у большинства видов на первом году жизни и сменяется придаточной бахромчатой или кистевого типа.
Необходимо отметить, что типы корневых систем значительно варьируют в зависимости от местообитаний. Например, для Rhodiola rosea наиболее обычна гетероризная корневая система, с хорошо развитым главным корнем и в разной степени развитыми боковыми и придаточными корнями, однако в условиях скальных экотопов формируется стержневая корневая система, со слабо развитыми боковыми корнями, а в мелкоземистых местообитаниях – кистекорневая (Фролов, Полетаева, 1998).
Корневища изученных Sedoideae надземные (Hylotelephium pluricaule, Sedum acre) или подземные; гипогеогенные, эпигеогенные или смешанные. Они выполняют функции отложения запасов, возобновления и вегетативного размножения. Корневища Rhodiola и Pseudosedum lievenii, несут чешуевидные метаморфизированные листья. Четко выделяются два типа корневищ – моноподиальное, формирующееся посредством медленного нарастания верхушечной почки, и симподиальное с ежегодным перевершиниванием, складывающееся из годичных симподиальных приростов. Первые характерны для представителей рода Rhodiola, вторые для всех прочих родов изученных Sedoideae. В целом, анатомическое строение симподиальных корневищ сходно с таковым побегов, но только в корневищах некоторых видов были обнаружены вторичные лучи (Hart, 1994). Моноподиальные корневища демонстрируют тесное родство с каудексами и, вероятно, являются их производными. Так, в корневищах Rhodiola quadrifida, как и в каудексе Rhodiola rosea, обнаружены два кольца камбия, причем внутренний «инвертированный» (Хоружая, 1976).
Образование запасающих корней довольно редкое явление в Sedoideae. H. ‘t Hart (1994) отмечает, что из почти 500 представителей Sedoideae только 4-5 имеют запасающие корни, в том числе Hylotelephium pseudospectabile и H. triphyllum. Клубневые корни, как и каудексы, и корневища выполняют запасающую функцию и состоят главным образом из крупных паренхимных клеток, заполненных гранулами крахмала и мукополисахаридами (Hart, 1994). Необходимо отметить, что большинство представителей Hylotelephium секции Hylotelephium имеют утолщенные (часто шнуровидные) корни, имеющие сходное анатомическое строение с клубневыми.
Опушение. Среди изученных Sedoideae обычны голые, без опушения виды, лишь четыре представителя имеют более или менее выраженные трихомы: Aizopsis selskiana, A.× pilosa, Sedum albertii и Orostachys paradoxa. Эти виды несут кроющие, обычно одноклеточные неразветвленные трихомы; очень короткие и редкие, заметные только на анатомических препаратах у Orostachys paradoxa или видимые невооруженным глазом мелкие папиллы на листьях у Sedum albertii, или хорошо заметные волоски у A. selskiana и A.× pilosa. Тип опушения, как правило, является стабильным признаком, но у A. selskiana значительно варьирует, встречаются представители не только с различной интенсивностью опушения и длиной волосков, но и с многоклеточными разветвленными волосками (рис. 21. 8).
На основании наличия опушения A. selskiana был выделен в монотипную серию Villosae (Борисова, 1969), а затем нами в секцию Villosae (Gontcharova, 1999a). Считалось, что наличие трихом является уникальным признаком, четко отграничивающим Villosae от типовой секции. Однако позже нами был описан межсекционный гибрид A.× pilosa (Гончарова и др., в печати), также имеющий опушение.
4.3. Эволюционное значение признаков
Все авторы, делавшие попытки установить филогенетические взаимоотношения в семействе на основании морфологических признаков приходили к выводу, что это весьма сложно по причине высокой степени их гомоплазии. К числу таких признаков, неоднократно возникавших в процессе формирования семейства, относят: розеточные биоморфы, однолетность, моноподиальное нарастание системы побегов, каудексы и запасающие корни, вторичное одревеснение побегов, супротивное или мутовчатое листорасположение, редукцию или увеличение частей цветка, срастание лепестков, чашелистиков и образование «цветочной трубки», число кругов тычинок, раздельнополость цветков, форму зрелых листовок, орнаментацию экзотесты (Hart, 1982; Ham, Hart, 1998; Mort et al., 2001; Mayuzumi, Ohba, 2004).
H. Ohba (1989) при выяснении филогенетических взаимоотношений в роде Rhodiola, сделал попытку выявить апомофные и плезиоморфные признаки, а позже H. ‘t Hart и J. Koek-Noorman (1982) и H. ‘t Hart (1992) предложили сочетания таких признаков для семейства в целом (табл. 3). И хотя эти авторы утверждают, что их построения подтверждены молекулярно-филогенетическими исследованиями, на наш взгляд это не всегда очевидно.
Как отмечено выше, анатомическое строение стебля Crassulaceae (наличие спиральных и кольчатых утолщений сосудов) не типично для обычного строения древесины. Объяснением подобного феномена может быть либо гетеробатмия признаков, либо педоморфоз (Jensen, цит. по: Hart, Koek-Noorman, 1989). Теория образования нетипичного вторичного одревеснения в результате педоморфоза была предложена и развита S. Carlquist (1962). В основу теории положено строение вторичной ксилемы, которая содержит признаки первичной.
Таблица 3
Филогенетические признаки Crassulaceae (по: Hart, Koek-Noorman, 1982; Hart, 1992; Ohba, 1989)
Признак |
Состояние признака |
плезиоморфное |
апоморфное |
Число частей цветка |
пятимерный цветок |
полимерные цветки |
Тип андроцея |
обдиплостемонный андроцей |
гаплостемонный андроцей |
Пыльники |
базификсные |
дозификсные |
Тип плодолистика |
киптокарпные плодолистики |
ортокарпные плодолистики |
Тип завязи |
верхняя |
полунижняя |
Срастание частей цветков |
свободные чашелистики и лепестки |
сростные лепестки и чашелистики |
Пол цветка |
гермафродитные |
раздельнополые |
Соцветие |
терминальные |
боковые |
Число цветков и соцветий |
много |
мало, до единичных |
Орнаментация экзотесты |
сетчато-бородавчатая |
ребристая |
Листорасположение |
спиральное |
перекрестно-парное |
Листья |
сохраняющиеся |
опадающие |
«Корневищные» листья |
присутствуют |
отсутствуют |
Вторичная ксилема в стебле |
отсутствует |
присутствует |
Край листа |
цельнокрайние |
зубчатые и пильчатые |
Толщина листа |
плоские листья |
вальковатые листья |
Опушение |
простое (нежелезистое) |
железистое или отсутствует |
Генеративные побеги |
опадающие |
сохраняющиеся |
Тип побега |
удлиненные |
розеточные |
Положение побега в пространстве |
ортотропные |
анизотропные |
Продолжительность жизни растения |
многолетность |
однолетность |
S. Carlquist (1962) описал это явление в том числе и у Crassulaceae. Автор считал, что явление педоморфоза во вторичной ксилеме встречается у высокоспециализированных древесных растений, развившихся от травянистых (или точнее мягкодревесных) предков, таких как розеточные деревья и стеблевые суккуленты. В противоположность камбию родственных травянистых представителей семейства, камбий розеточных деревьев и стеблевых суккулентов долго остается активным, и некоторые признаки строения первичной ксилемы сохраняются во вторичной. Однако вторичная ксилема изученных нами представителей Sedoideae наряду с примитивными признаками имеет и весьма продвинутые – волокна либриформа, простые перфорационные пластинки. На наш взгляд, более верно предположение L. Jensen, что подобное строение вторичной ксилемы Crassulaceae является результатом неодинакового уровня развития и специализации взрослых органов, достигнутого в результате относительной независимости в развитии различных частей организма в процессе эволюции – гетеробатмии.
Вопрос об исходном типе соцветия и направлениях эволюции соцветий до сих пор остается дискуссионным. Главный предмет споров – что считать первичным с одиночный цветок или соцветие? На наш взгляд, этот вопрос должен решаться для каждого конкретного таксона отдельно в контексте истории его формирования и развития. Предлагаемая нами схема преобразований соцветий восточно-азиатских Sedoideae (рис. 23) в целом соответствует схемам, предложенным А.Л. Тахтаджяном (1964), G.L. Stebbins (1974) и Л.И. Лотовой (2001). За исходный тип нами принята облиственная простая закрытая кисть (простой облиственный верхоцветник по А.Л. Тахтаджяну, 1980) – фрондозное монотелическое соцветие с небольшим количеством боковых ответвлений. Подобный тип соцветия встречается в евроазиатском роде Rosularia (Eggli, 1988). Под влиянием разнообразных климатических условий анцестральное соцветие преобразовалось, с одной стороны, до одиночного цветка посредством сокращения количества боковых осей и редукции боковых цветков до 1 (как в подроде Primuloides рода Rhodiola или у некоторых представителей Sedum s. str.); с другой стороны – посредством усложнения ветвления главной оси до сложных многоцветковых соцветий (как у некоторых представителей Hylotelephium).
|
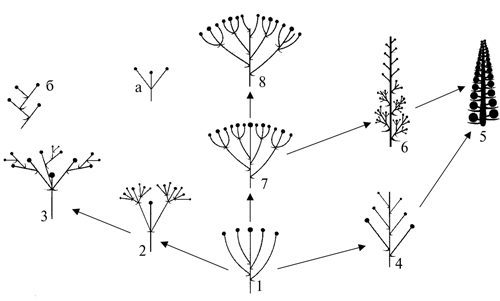
Рис. 23. Возможная схема преобразований соцветий восточно-азиатских Sedoideae. 1. Простой монохазий; 2. Сложный дихазий; 3. Плейохазий; 4. Простая открытая кисть; 5. Простой колос; 6. Сложный колос; 7. Закрытый тирсоид; 8. Открытый тирс. а-б – парциальные соцветия: а – дихазий, б – монохазий (извилина).
|
На наш взгляд, образование малоцветковых соцветий представителей Rhodiola связано с суровыми условиями существования предковых форм, эволюционировавших в условиях высокогорий, – криофилизацией и ксерофитизацией местообитаний, сокращением длины вегетационного периода. W. Troll (1964, 1969) экспериментально установил, что, выращивая растения на обедненном субстрате, можно получить экземпляры с малоцветковыми соцветиями и даже с одиночными цветками. G.L. Stebbins (1974) утверждал, что большое количество малоцветковых соцветий имеет преимущества перед малым количеством многоцветковых, благодаря равномерному распределению питательных веществ, необходимых для формирования диаспор. Однако энергетические ресурсы для зеленых растений не всегда играют роль главного фактора, обусловливающего направление преобразований органов растений, и соцветий в частности.
Несмотря на несомненную важность подобных построений для решения вопросов структурной эволюции растений, необходимо, прежде всего, учитывать биологическую важность преобразований соцветий. Так, R. Wyatt (1982) писал, что эволюционный процесс преобразования соцветий должен быть направлен на улучшение качества опыления и увеличение количества жизнеспособных семян, а также эффективное распространение пыльцы и семян. Архитектура соцветий может сильно влиять на перекрестное опыление растений. Это влияние проявляется как во времени, так и в пространстве (Лотова, 2001). Главное преимущество соцветий перед отдельным цветком не столько в его размерах, сколько в разновременности (гетерохронии) зацветания отдельных цветков, что значительно повышает шансы опыления в неблагоприятных климатических условиях (Лотова, 2001) и вероятность вовлечения большего числа партнеров, участвующих в перекрестном опылении.
Разновременность развития плодов также повышает вероятность их лучшего созревания и оптимального распространения. Наиболее выгодное положение в переменных климатических условиях по отношению к влиянию осадков на опыление и распространение семян занимают полителические соцветия (Wyatt, 1982).
Однако, на наш взгляд, большинство разнообразных признаков морфологии соцветий Sedoideae является селективно нейтральным и относительно функционально равнозначным. «В историческом развитии таких признаков на первый план выступают морфогенетические тенденции неадаптивного характера» (Красилов, 1989, с.192). И хотя морфогенетическая корреляция с отбираемым признаком может придать их эволюции некоторую направленность, она почти не проявляется (Красилов, 1989).
Очередное (спиральное) листорасположение считается примитивным для покрытосеменных (Cronquist, 1970). И, казалось бы, подобные тенденции сохраняются и у Crassulaceae, так как в основании большинства клад располагаются виды с очередным листорасположением, но анализ онтогенеза изученных Sedoideae показал, что все изученные длиннопобеговые представители родов Hylotelephium и Aizopsis в ювенильной фазе имеют 2-5 пар перекрестно-парных листьев, что, на наш взгляд, является отражением «основного биогенетического закона» и подтверждением первичности супротивного листорасположения, по крайней мере, в подсемействе Sedoideae.
Мутовчатое листорасположение – бесспорно апоморфный признак, возникавший несколько раз в подсемействе. С мутовчатым филлотаксисом сопряжена и вивипария Hylotelephium, в этом роде только видам с подобным листорасположением присуща гомофазная вивипария.
Апоморфность ортокарпных листовок Crassulaceae R.C.H.J. van Ham и H. 't Hart (1998) считают бесспорной, однако Н. Ohba (1978) полагает, что более эволюционно продвинуты киптокарпные плодолистики. На филогенетическом дереве Telephium-клады виды, имеющие киптокарпные листовки (Phedimus, Aizopsis), занимают место в серединной части (рис. 9), что позволяет также говорить либо о сохранении предкового состояния признака, либо о вторичном упрощении. Но если принять во внимание общее направление в эволюции плода от апокарпного к синкарпному (Тахтаджян, 1964), то киптокарпные (гемисинкарпные) многолистовки – более эволюционно продвинутый тип плода по сравнению с ортокарпными (апокарпными).
Таким же образом для Sedoideae не подтверждается и апоморфность розеточных, полурозеточных и анизотропных побегов (см. гл. 7.6).
Вальковатые листья с придатками – бесспорный синапоморфный признак, характеризующий Orostachys sect. Orostachys subsect. Appendiculatae.
Число лепестков в цветке более 5 считается R.C.H.J. van Ham и H. 't Hart (1998) апоморфным признаком, а обдиплостемонность – плезиоморфным, можно допустить, что это именно так для Crassulaceae, но у изученных Sedoideae все цветки с многолепестковыми цветками – обдиплостемонные (некоторые Orostachys, Aizopsis kamtschatica), то есть наблюдается сочетание «продвинутых» и «примитивных» признаков. Гаплостемонный андроцей признан более эволюционно продвинутым по сравнению с обдиплостемонным (сохранившим относительно примитивное спиральное расположение частей) (Ohba, 1978). Для изученных Sedoideae, вероятно, апоморфией можно считать редукцию частей цветка, характерную для Rhodiola, например, у типового вида Rhodiola rosea встречаются как 4-мерные гаплостемонные цветки, так и 5-мерные обдиплостемонные и полный спектр переходных форм (Ohba, 1981).
Синапоморфными признаками клады Rhodiola/Pseudosedum является, наличие чешуевидных листьев на корневищах и каудексах.
Таким образом, наши исследования позволяют предположить, что плезиоморфными (предковыми) признаками восточно-азиатских Sedoideae являются: обдиплостемонный андроцей, перекрестно-парные листья, свободные чашелистики и лепестки, отсутствие вторичной ксилемы в стебле, апокарпные листовки, верхние завязи, пятимерный цветок, полурозеточные побеги, терминальные закрытые соцветия, многолетность. Апоморфными (эволюционно продвинутыми) – гаплостемонный андроцей, спиральное листорасположение, сростные лепестки и чашелистики, наличие вторичной ксилемы, гемисинкарпные листовки, полунижние завязи, открытые соцветия, полимерные цветки, удлиненный побеги.
Оригинальный рисунок А.Р. De Candolle www.illustratedgarden.org/mobot/rarebooks, Rhodiola pinnatifida nature.chita.ru.
Читать далее: главу 5
|