ГЛАВА 6
ФЛОРИСТИЧЕСКАЯ СТРУКТУРА РАСТИТЕЛЬНОГО ПОКРОВА ЛАНДШАФТНЫХ РАЙОНОВ
В процессе многолетних ботанико-географических и флористических исследований методом конкретных флор нами было изучено несколько конкретных флор в разных природных зонах и в разных типах ландшафтов страны. В каждом из этих районов тщательно выявлялось таксономическое разнообразие, составлялись геоботанические описания модельных контурфитоценозов, проводилась их типизация. Основная цель этих исследований - установление зависимости таксономической неоднородности контурфитоценозов от физико-географических особенностей ландшафта. Важно установить, насколько таксономическая неоднородность контурфитоценоза зависит от видового состава флоры и ее ареалогической структуры как в разных природных зонах, так и в разных типах ландшафтов в связи изменением характера ведущих экологических факторов и изменением общего разнообразия ландшафтов. Под таксономической хорологической структурой растительного покрова ландшафтного района мы понимаем систему бинарных отношений флористического сходства-различия на множестве описаний элементарных фитогеохор - подразделений растительного покрова в ранге контурфитоценозов и микрокомбинаций, или на множестве типологических объединений элементарных фитогеохор - групп ассоциаций или типов местообитаний. Полевые работы выполнялись с 1971 по 1988 г. на Западной Чукотке сначала в составе Полярной экспедиции БИНа АН СССР им. В. Л. Комарова, а затем в экспедициях ИБПС ДВО АН СССР, с 1973 по 1977 г. полевые работы проводились в Калининградской области, с 1976 по 1978 г. - на Восточном Алтае в Алтайском заповеднике, с 1978 по 1982 г. - в Коми АССР и с 1982 по 1984 г. - в Южном Забайкалье в Сохондинском заповеднике.
В пределах района 10х20км закладывались пробные площадки
размером 100 кв. метров для травянистой и кустарничковой растительности
и размером 400 кв. метровдля лесной и кустарниковой. Каждая пробная
площадь располагалась в наиболее однородной части контурфитоценоза.
Затем все описания подразделялись на группы ассоциаций или типы
местообитаний по совокупности признаков. В один тип местообитаний или
группу ассоциаций объединялись контурфитоценозы, сходные по положению в
рельефе, характеру и интенсивности увлажнения, характеру подстилающих
горных пород, типу почвы, набору доминирующих видов и общему
флористическому составу.
Для каждого типа местообитаний составлялся полный список
встречающихся в нем видов. Далее такие списки использовались при
составлении сводных таблиц распределения видов конкретной флоры по типам
местообитаний. Объем выделенных типов местообитаний, как уже отмечено
выше, приблизительно соответствовал рангу групп ассоциаций, традиционно
выделяемых геоботаниками ленинградской школы. Типы местообитаний в
дальнейшем рассматривались как экологические состояния, или, точнее, как
ансамбли состояний, в которых находится растительный покров данного
ландшафта. На этом множестве типов местообитаний устанавливалась система
бинарных отношений сходства – различия по общности видового состава.
Оценка сходства производилась путем учета присутствия – отсутствия видов
с помощью множественных мер Жаккарда. Результаты записывались в виде
матриц по парного сходства типов местообитаний.
Организация и дальнейший анализ этих матриц выполнялся
графически (Галанин, 1973, 1977, 1979, 1980 а, б). Типы местообитаний
изображались на графах в виде кружков с соответствующими номерами.
Сходные типы по возможности располагались ближе друг к другу и
соединялись линиями, толщина и характер которых соответствовали
различным уровням флористического сходства. Графы изображались на
плоскости, структура их многомерна, поэтому в их изображении на
плоскости имеются искажения: расстояния между кружками не всегда прямо
пропорциональны степени сходства соответствующих типов местообитаний.
Для устранения этих искажений графы необходимо изображать в многомерном
пространстве. Полученные графы можно рассматривать в качестве моделей
хорологической структуры растительного покрова в том случае, если
множество типов местообитаний охватывает все основное экологическое
разнообразие растительного покрова данного ландшафтного района.
В известной степени такие графы отражают и экологическую
структуру ландшафта. Такие модели по существу выступают как
ординационные схемы, а не классификация. В связи с этим они могут
анализироваться не только на предмет наличия скоплений типов
местообитаний, которые мы называем классами и надклассами местообитаний,
но и на предмет наличия рядов сходства типов местообитаний, которые
можно рассматривать как изменения экологических режимов, связанные с
определенными факторами или группами тесно скоррелированных факторов.
Границы классов и подклассов местообитаний мы проводили по ослабленным
уровням флористического сходства типов местообитаний. Некоторые классы
местообитаний оказались представленными несколькими типами
местообитаний, другие двумя типами и даже одним. В пределах классов
местообитаний, представленных многими типами, выделяются подклассы,
центры и ядра.
РАСТИТЕЛЬНЫЙ ПОКРОВ РАЙОНА ДЕЛЬТЫ НЕМАНА В КАЛИНИНГРАДСКОЙ ОБЛАСТИ
Район дельты Немана находится на границе Эстонско-Литовской приморской провинции выпуклых болот широколиственно-хвойных лесов и Южно-Балтийской приморской провинции выпуклых болот и широколиственных лесов Среднеевропейской области хвойно-широколиственных лесов (Кац, 1971).
Флора района носит переходный характер между среднеевропейскими и восточноевропейскими флорами.
Дельтовая низменность расположена в тектонически погружающейся части Приморской равнины. В ледниковое время поверхность плато была выровнена и перекрыта толщей моренных отложений. После отступления последнего валдайского ледника (около 15–18 тыс. лет назад) с линии Инстручских высот (краевые морены) местность приобрела вид холмистой равнины со слабым наклоном к северу. Несколько позднее (13–15 тыс. лет назад), при остановке ледника на линии Клайпедских морен, равнина оказалась затопленной водами послеледникового озера. В результате деятельности этого водоема поверхность моренной равнины была перекрыта наносами, получившими название боровых песков. Затем, по мере отступания ледника и приледникового озера, боровые пески перекрываются древнедельтовыми отложениями. В результате гляциоэвстатического подъема территории усилилась эрозионная деятельность реки, что привело к размыву боровых песков и древнедельтовых отложений. Наконец, подъем сменился погружением и снова началась активная аккумуляция глин. Этот процесс продолжается и в настоящее время. Низменность представляет собой плоскую полого наклоненную на северо-запад равнину. Абсолютные высоты до 5–6 м. Некоторые участки находятся ниже уровня моря, и уровень грунтовых вод здесь регулируется с помощью дамб и насосных станций. Площади польдерных земель в дельтовой низменности весьма значительны.

|
Низменность представляет собой плоскую полого наклоненную на северо-запад равнину. Абсолютные высоты до 5–6 м. Некоторые участки находятся ниже уровня моря, и уровень грунтовых вод здесь регулируется с помощью дамб и насосных станций. Площади польдерных земель в дельтовой низменности весьма значительны.
На территории дельты выделяют три генетические группы почвообразующих пород: современный аллювий, древний аллювий, и болотные отложения (Завалишин, Надеждин, 1961; Рябой, 1979). Высокие и средние части дельты сложены суглинками и песками, нижние, прилегающие к Куршскому заливу, - торфяниками. Все разновидности почв подвержены оглеению, но в разной степени, зависящей от уровня грунтовых вод. В пойме почвы супесчаные, слоистые, слабо гумусированные, дерновые. Почвы высокой части дельты в основном также супесчаные аллювиальные, но глубоко гумусированные. В полосе, прилегающей к заливу, имеются значительные массивы низинных болот с торфяно-глеевыми перегнойными (мощность торфяного горизонта менее 1 м.) и торфяными перегнойными (мощность торфа несколько метров ) почвами. В центральных частях низинных болот встречаются участки олиготрофных болот верхового типа.
|

|
Естественная растительность района дельты Немана сильно изменена в результате многовековой хозяйственной деятельности человека. В меньшей степени антропогенному воздействию подвергались черноольховые крупноосоковые и крупноразнотравные леса. На слегка выпуклых участках низменности встречаются небольшие массивы хвойно-широколиственных лесов. Осушенные участки заняты полями, сеяными лугами и пастбищами. Многие пастбища сильно деградировали в результате перевыпаса крупного рогатого скота.
Климат Калининградской области формируется за счет частых затоков воздуха умеренных широт со стороны Атлантического океана. Он может быть охарактеризован как переходный от морского к умеренно континентальному. Зимой погода относительно теплая (средняя температура января -3,5°С), лето же, напротив, прохладное (средняя температура июля +17°С ). Повышенная влажность способствует развитию низкой облачности и увеличению повторяемости облачных дней. Относительная влажность воздуха зимой и осенью 85–90 %, весной и летом она снижается до 70 %. В среднем за год выпадает около 800 мм осадков. Безморозный период длится 165 дней. Снеговой покров часто неустойчивый и в течение зимы неоднократно разрушается (Очерки…, 1969).
|
Флора ландшафтного района, по нашим данным, насчитывает
400 видов высших сосудистых растений. Ведущими по числу видов семействами
являются сложноцветные (11,5%), злаковые (9,4%), осоковые (7,2%),
гвоздичные (5,1%), розоцветные (4,6%), бобовые (4,3%), крестоцветные
(4,3%), гречишные (3,3%), губоцветные (3,3 %) и лютиковые (2,8%).
Циркумполярных и почти циркумполярных видов во флоре 20%, европейских –
14%, западноевропейских – 2,8%, евроазиатских – 14,3%,
американо-европейских – 4,6 %, европейско-западноазиатских – 35,8%
и видов с неопределенными долготными типами ареалов 8,7%. По широтным
ареалогическим группам ареалов виды флоры распределяются следующим
образом: неморально-бореальные составляют 78,7%, неморальные – 15,2% и
полизональные – 6,1%. Наиболее интересные флористические находки в этом
районе следующие: Platanthera bifolia, Dactylorhiza fuchsii, Betula
humilis, Alnus incana, Cerastium stevenii, Sagina nodosa, Thalictrum
lucidum, Drosera anglica, D. rotundifolia, Rubus chamaemorus, Oxalis
stricta, Viola epipsila, V. palustris, Hyppuris monotropa, Ledum
palustre, Andromeda polifolia, Oxycoccus palustris, Vaccinium
uliginosum, Pedicularis palustris, Petasites spurius.
(Названия видов здесь и далее приведены по С. К.
Черепанову).
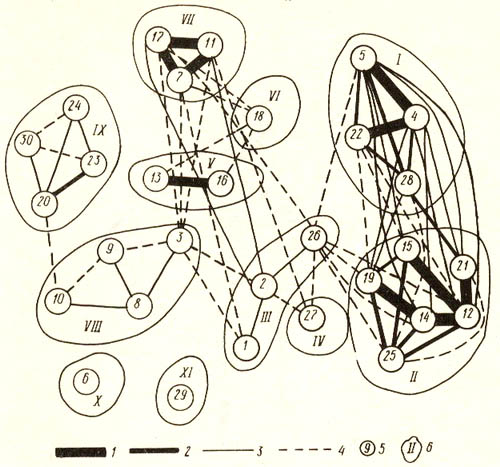 |
Всего в пределах района составлено 300 геоботанических описаний и выделено 30 типов местообитаний, которые характеризуются далее в тексте номерами, соответствующими номерам на рисунках. Результаты попарного сравнения типов местообитаний на предмет флористического сходства по Жаккару представлены в виде графа (см. рис. слева). Всего на графе выделено 11 классов местообитаний. Таким образом, выявленная структура имеет явно иерархический характер.
Граф флористического сходства типов
местообитаний района дельты реки Неман в Калининградской области .
Условные обозначения: 1 – 4 – уровни флористического сходства, по
Жаккару, % : 1 – более 60 процентов; 2 – более 50, но менее 60; 3
– более 40, но менее 50; 4 – более 30, но менее 40 процентов; 5 –
типы местообитаний (характеристика местообитаний дана в тексте); 6
– номера и границы классов местообитаний. |
ОПИСАНИЕ СТРУКТУРЫ РАСТИТЕЛЬНОГО ПОКРОВА С
ПОМОЩЬЮ ГРАФА
I. Класс местообитаний относительно сухих
положительных элементов микрорельефа с песчаными почвами и луговой
растительностью
5. Склоны дамб в лесу. Сухие разнотравно-злаковые
высокотравные луга, изрядно закустаренные. Субстрат песчаный и
супесчаный. Увлажнение нормальное при интенсивном дренаже. Грунтовые
воды залегают на глубине не менее 1,5 м. Основу травостоя обычно
составляют: Phleum pratense, Calamagrostis epigeios, Arrhenatherum
elatius, Dactylis glomerata, Poa pratensis, P. trivialis, Bromus mollis,
Carex leporina, Agropyron repens, Ranunculus acris, Berteroa incana,
Potentilla anserina,Trifolium repens, Vicia cracca, Lathyrus
pratensis,Taraxacum officinale и др.
4. Склоны дамб на открытых местах. Сухие
разнотравнозлаковые низкотравные луга. Субстрат аналогичен предыдущему
типу. Увлажнение периодически недостаточное в связи с интенсивным
дренажем. Здесь наиболее характерны следующие виды: Eguisetum arvense,
Phleum pratense, Arrhenatherum elatius, Dactylis glomerata, Poa
pratensis, Festuca polesica, F. pratensis, F. rubra, Bromus inermis, B.
mollis, Agropyron repens, Polygonum aviculare, Arenaria serpyllifolia,
Dianthus deltoides, Cardaminopsis arenosa, Berteroa incana, Sedum acre,
Potentilla anserina, P. argentea, Medicago falcata , M. lupulina, Lotus
corniculatus, Anthyllis macrocephala, Alchemilla monticola, Daucus
carota, Achillea millefolium, Hieracium pilosella, Plantago
lanceolata.
22. Сухие песчаные бугры и береговые валы естественного
происхождения. Сухие разнотравно-злаковые низкотравные луга. Субстрат
супесчаный и песчаный. Увлажнение недостаточное в связи с интенсивным
дренажем. Наиболее характерные виды: Anthoxanthum odoratum, Holcus
mollis, Phleum pratense, Calamagrostis epigeios, Agrostis tenuis,
Arrhenatherum elatius, Corynephorus canescens, Poa pratensis, Festuca
rubra, Carex hirta, C. praecox, Luzula campestris, Rumex acetosella,
Cerastium arvense, C. holosteoides, Scleranthus annuus, Trifolium
arvense, T. medium, T. repens и др.
28. Склоны насыпей шоссейных дорог. Сухие
разнотравнозлаковые низкотравные луга. Субстрат песчаный и супесчаный.
Увлажнение периодически недостаточное. Здесь характерны следующие виды:
Equisetum arvense, Arrhenatherum elatius, Dactylis glomerata, Poa annua,
P. trivialis, Lolium perenne, Bromus inermis, B. mollis, Carex hirta,
Rumex acetosella, Thlaspi arvense, Descurainia sophia, Sedum acre,
Potentilla anserina, Lotus corniculatus, Vicia sepium, Hypericum
perforatum, Linaria vulgaris, Plantago media и др.
Во всех четырех типах этого класса почвы дерновые,
достаточно гумусированные. Во всех сообществах ценотическая структура
выработана хорошо, эти участки не перепахиваются, практически никогда не
выкашиваются и используются как пастбища.
II. Класс местообитаний осушенной части поймы
Немана, занятой травянистой растительностью
21. Старые заброшенные хутора на слегка возвышенных
участках. Луга среднего уровня с большой примесью рудеральных растений.
Увлажнение нормальное, уровень трофности несколько повышенный. Здесь
обычно встречаются : Alopecurus pratensis, Agrostis stolonifera, Poa
annua, P. pratensis, P. trivialis, Festuca rubra, Lolium perenne, Bromus
mollis, Agropyron repens, Urtica dioica, Rumex confertus, Polygonum
aviculare, P. scabrum, Chenopodium album, Ch. rubrum, Cirsium arvense и
др.

|
12. Оторфованные участки осушаемой поймы. Сеяные злаковые луга на осушенных окультуренных торфяниках. Характерные виды : Phleum pratense, Alopecurus pratense, Agrostis stolonifera, Poa pratensis, Festuca pratensis, Carex nigra, Stellaria graminea, S. media, Barbarea vulgaris, Capsella bursa-pastoris, Erysimum cheiranthoides, Berteroa incana, Vicia sepium, Daucus carota, Myosotis arvensis, Glechoma hederacea, Veronica chamaedrys, Odontites serotina, Plantago lanceolata, Galium mollugo, Cirsium arvense и др.
14. Оторфованные участки осушаемой поймы. Сильно деградировавшие в результате перевыпаса сеяные злаковые луга на осушенных окультуренных торфяниках. От предыдущего типа отличаются только степенью пастбищной дигрессии. Здесь наиболее обычны Phleum pratense, Alopecurus pratensis, Agrostis stolonifera, Deschampsia caespitosa, Poa annua, P. pratensis, Carex nigra, Polygonum aviculare, P. hydropiper, P. scabrum, Ranunculus acris, R. repens, Rorippa islandica, Potentilla anserina и др.
|
15. Cупесчаные участки осушаемой поймы. Сеяные злаковые
луга. Увлажнение нормальное благодаря действию осушительной системы.
Характерные виды: Phleum pratense, Alopecurus pratensis, Agrastis
stolonifera, Poa pratensis, P. trivialis, Festuca pratensis, Rumex
confertus, Polygonum convolvulus, Stellaria graminea, S. media,
Ranunculus repens, Rorippa islandica, Capsella bursa-pastoris,Trifolium
hybridum,T. repens, Vicia sepium, V. tetrasperma, Myosotis arvensis,
Leucanthemum vulgare и др.
19. Супесчаные участки осушенной поймы. Сильно
деградировавшие под воздействием перевыпаса сеяные злаковые луга. От
предыдущего типа отличаются только степенью пастбищной дигрессии. Здесь
наиболее характерны следующие виды: Anthoxanthum odoratum, Phleum
pratense, Agrostis stolonifera, Deschampsia caespitosa, Poa annua, P.
pratensis, Lolium perenne, Carex leporina, Juncus effusus, Rumex
acetosa, R. crispus, Ranunculus repens, Potentilla norvegica, Trifolium
repens, Lamium album, Rhinanthus vernalis, Plantagol anceolata, P.
major, Senecio vulgaris и др.
25. Супесчаные участки осушенной поймы. Поля с посевами
зерновых культур. Сорнополевая растительность. Здесь часты : Equisetum
arvense, Phleum pratense, Festuca pratense, Lolium perenne, Rumex
acetosella, Polygonum aviculare, P. convolvulus, Chenopodium album,
Stellaria media, Spergula arvensis, Fumaria officinalis, Thlaspi
arvense, Barbarea vulgaris, Capsella bursa-pastoris, Potentilla
argentea, Melilotus albus, Galeopsis speciosa, G. tetrahit, Mentha
arvensis, Plantago major, Centaurea cyanus, C. jace, Cirsium arvense,
Vicia cracca.
Ведущие факторы изменения флористического состава
растительности в этом классе местообитаний – интенсивность увлажнения,
характер почв, степень пастбищной дигрессии, осушение, характер посевов.
Тип экологического режима здесь можно классифицировать как аграрный
польдерный.
III. Класс переувлажненных местообитаний неосушенной поймы
с гидро- и гигрофильной растительностью
1. Прибрежные отмели Куршского залива. Прибрежно-водная
растительность. Увлажнение постоянно избыточное, субстрат песчаный,
заиленный. Засоление практически отсутствует, так как Куршский залив
отделен от Балтийского моря длинной косой и имеет почти пресную воду,
приносимую р. Неман. Здесь обитает весьма своеобразный комплекс видов:
Equisetum fluviatile, Typha latifolia, Sparganium simplex, Alisma
plantagoaquatica, Digraphis arundinacea, Phragmites communis,
Calamagrostis canescens, Deschampsia caespitosa, Poa palustris,
Puccinellia distans, Glyceria maxima, Scirpus lacustris, S. sylvaticus,
Eriophorum vaginatum, Eleocharis quinquiflora, Carex acuta, C. otrubae,
Acorus calamus, Iris pseudocorus, Polygonum hydropiper, P. persicaria,
Caltha palustris, Barbarea stricta, Cardamine amara, Comarum palustre,
Lythrum salicaria и др.
2. Неосушенные участки на низком берегу Куршского залива.
Сырые разнотравно-осоковые луга. Увлажнение постоянно избыточное. Почва
супесчаная, слегка оторфованная. Здесь наиболее характерны: крупных
рукавов. Сырые разнотравно-злаково-осоковые луга. Почвы супесчаные
оторфованные оглеенные. Увлажнение постоянно избыточное. Хорошо развиты
аллювиальные процессы. Наиболее характерные виды Equisetum fluviatile,
Digraphis arundinacea, Phragmites communis, Alopecurus pratensis,
Agrostis stolonifera, Poa palustris, Glyceria maxima, Carex acuta, C.
otrubae, C. rostrata, Salix pentandra, S, triandra, S. viminalis, Rumex
crispus, Polygonum persicaria, Coronaria floscuculi, Caltha palustris,
Ranunculus repens, Bidens tripartita, Valeriana exaltata и др.

|
IV. Класс местообитаний пойменных ивняков
27. Неосушенные прирусловые участки в пойме р. Неман и его рукавов. Разнотравно-осоковые пойменные ивняки. Почвы супесчаные, заиленные. Увлажнение периодически избыточное, периодически нормальное. В современном ландшафте ивняки занимают небольшие площади среди окультуренной растительности сеяных лугов. Здесь наиболее обычны следующие виды: Equisetum pratense, Digraphis arundinacea, Phragmites communis, Agrostis stolonifera, Deschampsia caespitosa, Poa trivialis, Carex nigra, C. acuta, Salixa lba, S. dasyclados, S. pentandra, S. triandra, S. viminalis, Urtica dioica, Silene alba, Thalictrum aquilegifolium, Impatiens nolitangere, Lhytrum salicaria, Cuscuta europaea, Myosotis palustris и др.
V. Класс переувлажненных местообитаний осушенной части поймы
13. Вымокаемые участки в виде локальных понижений в микрорельефе среди польдеров на торфянистых почвах. Переувлажненные разнотравно-осоковые луга с очень слабо выработанной ценотической структурой. Здесь характерны: Phleum pratense, Alopecurus geniculatus, Deschampsia caespitosa, Poa pratensis, Festuca pratensis, Puccinellia distans, Carex acuta, C. flava, C. nigra, Juncus bufonius, J. effusus, Polygonumaviculare, P. hydropiper, Ranunculus repens, Comarum palustre, Bidens tripartita и др.
|
16. Вымокаемые участки в виде локальных понижений в
микрорельефе среди польдеров на песчаных и супесчаных почвах.
Переувлажненные разнотравно-осоковые луга. Здесь наиболее характерны:
Alopecurus geniculatus, Deschampsia caespitosa, Poa pratensis, Glyceria
fluitans, Carex flava, C. nigra, Juncus bufonius, J. effusus, Rorippa
islandica, Ranunculus repens, Comarum palustre, Scutellaria
galericulata, Trifolium repens, Galeopsis speciosa, G. tetraphit, Mentha
arvensis, Gnaphalium uliginosum и др.
Следует обратить внимание, что в один класс местообитаний
по флористическому сходству объединяются местообитания с разными
почвами. Причиной этого, по всей вероятности, является то, что
ценотическая структура сообществ в обоих случаях не выработана, так как
участки перепахиваются через 5–7 лет.

|
VI. Класс переувлажненных местообитаний вдоль неглубоких осушительных канав
18. Мелкие пересыхающие канавы среди польдеров. Разнотравно-осоковые травянистые сообщества. Водный режим непостоянный. Временами увлажнение избыточное – вода стоит в канаве, временами канава пересыхает и увлажнение приближается к нормальному. Здесь наиболее обычны Phragmites communis, Alopecurus pratensis, Phleum pratense, Agrostis stolonifera, Deschampsia caespitosa, Dactylis glomerata, Poa palustris, P. trivialis, Glyceria fluitans, Eleocharis quinquiflora, Carex acuta, C. flava, C. pseudocyperus, C. riparia, Juncus effusus, Coronaria flos-cuculi, Ranunculus repens, R. sceleratus, Cardamine amara, Filipendula ulmaria, Potentilla anserina, Lythrum salicaria, Hottonia palustris, Galium palustre.
Польдерные земли в дельте реки Неман в основном используются как пастбища и сенокосы.
|
VII. Класс водных местообитаний осушительных канав и
каналов
7. Широкие и глубокие осушительные канавы в топких
черноольховых лесах. Прибрежно-водная разнотравно-осоковая
растительность. Субстрат торфянистый. Характерные виды: Equisetum
fluviatile, Typha latifolia, Sparganium simplex, Alisma
plantago-aquatica, Sagittaria sagittifolia, Elodea canadensis,
Stratiotes aloides, Hydrocharis morsis-ranae, Glyceria fluitans, G.
maxima, Carex pseudocyperus, C. riparia, C. rostrata, Calla palustris,
Rumex hydrolapatum, Polygonum persicaria, Rorippa amphibia, Cardamine
pratensis, Sium latifolium, Hottonia palustris, Galium aparine и др.
11. Широкие и глубокие осушительные каналы на лугах с
торфянистыми почвами. Прибрежно-водная разнотравно-осоковая
растительность. Здесь наиболее характерны: Typha latifolia, Elodea
canadensis, Stratipotes aloides, Hydrocharis morsis-ranae, Digraphis,
arundinacea, Phragmites communis, Poa palustris, Glyceria fluitans, G.
maxima, Eleocharis quinquiflora, Carex acuta, C. nigra, C.
pseudocyperus, C. riparia, C. rostrata, C. vesicaria, Acorus calamus,
Caltha palustris, Lemna minor, Juncus effusus, Epilobium palustre, Sium
latifolium, Oenanthe aquatica, Bidens tripartita и др.
17.Широкие и глубокие осушительные каналы на лугах с
песчаными почвами. Прибрежно-водная осоково-разнотравная растительность.
Характерные виды: Sparganium simplex, Typha latifolia, Equisetum
fluviatile, Alisma plantago-aquatica, Elodea canadensis, Stratiotes
aloides, Poa palustris, Glyceria maxima, Scirpus sylvaticus, Carex
otrubae, C. riparia, C. rostrata, Acorus calamus, Iris pseudocorus,
Rumex hydrolapatum, Nuphar lutea, Nymphea candida, Rorippa islandica,
Cardamine pratensis, Filipendula ulmaria, Lythrum salicaria, Sium
latifolium, Oenanthe aquatica, Menyanthes trifoliata и др.
VII. Класс местообитаний черноольховых лесов
3. Неосушаемые вогнутые участки дельтовой низменности.
Тонкие крупноосоковые черноольховые леса. Увлажнение постоянно
избыточное застойное. Почвы торфянистые. Здесь наиболее характерны:
Equisetum sylvaticum, Typha latifolia, Sparganium simplex, Alisma
plantago-aquatica, Phragmites communis, Calamagrostis arundinacea, Carex
rostrata, Acorus calamus, Calla palustris, Iris pseudocorus, Alnus
glitinosa, Urtica dioica, Ficaria verna, Barbarea stricta, Ribes nigrum,
R. pubescens, Filipendula ulmaria, Geum urbanum, Viola palustris,
Naumburgia thyrsiflora, Galium palustre.
10. Осушаемые вогнутые участки дельтовой низменности.
Сырые, но не топкие, частично осушенные крупнотравные черноольховые
леса. Почвы торфянистые. Увлажнение избыточное, периодически застойное.
Здесь наиболее обычны следующие виды: Dryopteris cristata, Equisetum
sylvaticumm, Typha latifolia, Sparganium simplex, Phragmites communis,
Calamagrostis arundinacea, C. canescens, C. purpurea, Glyceria maxima,
Scirpus sylvaticus, Carex acuta, C. brunnescens, C. canescens, C.
rostrata, Majanthemum bifolium, Polugonatum multiflorum, Paris
quadrifolia, Betula pendula, Alnus glutinosa, Humulus lupulus, Urtica
dioica, Stellaria nemorum, Tilia cordata, Viola palustris, Lamium album,
Sambucus racemosa и др.
9. Просеки в частично осушенных черноольховых лесах.
Травянистая разнотравная растительность, характерная для черноольховых
лесов. Проведение каналов ее изменило очень незначительно. Здесь обычны:
Dryopteris cristata, Typhal atifolia, Sparganium simplex, Alisma
plantago-aquatica, Calamagrostis arundinacea, Carex pseudocyperus, Alnus
glutinosa, Urtica dioica, Rorippa amphibia, Filipendula ulmaria, Geum
urbanum, Impatiens nolitangere, Lysimachia vulgaris, Naumburgia
thyrsiflora, Galistegia sepium, Solanum dulc-amara, Scrophularia nodosa,
Plantago major и др.
10. Слегка выпуклые осушаемые участки дельтовой
низменности. Черноольховые разнотравные леса с примесью березы, осины и
дуба. Почвы супесчаные оторфованные. Увлажнение избыточное, периодически
почти нормальное. Грунтовые воды залегают очень близко к поверхности.
Здесь наиболее обычныследующие виды: Dryopteris cristata, Athyrium
filixfemina, Equisetum sylvaticum, Milium effusum, Calamagrostis
arundinacea, C. canescens, C. purpurea, Carex brunnescens, Majanthemum
bifolium, Polygonatum multiflorum, Paris quadrifolia, Salix aurita, S.
caprea, Populus tremula, Betula pendula, Alnus glutinosa, Quercus robur,
Ulmus laevis, Stellaria nemorum, Moehringia trinervia, Cardamine amara,
Ribes nigrum, R. pubescens, Rubus idaeus, Frangula alnus, Tilia cordata,
Viola palustris, Aegopodium podagraria, Angelica sylvestris, Fraxinus
excelsior и др.
В этом классе местообитаний основным фактором,
обусловливающим изменчивость растительного покрова, является
интенсивность увлажнения. Наиболее гигрофильные условия в местообитании
3, а наименее – в местообитании 10. Антропогенный фактор в изменении
растительности этого класса местообитаний стоит на втором месте.
IX. Класс местообитаний хвойных и хвойно-широколиственных
лесов на повышениях микро- и мезорельефа гряды
20. Невысокие супесчаные и легко суглинистые холмы и гряды. Широколиственные и хвойно-широколиственные леса. Почвы супесчаные дерновые слабоподзолистые. Увлажнение нормальное. Здесь наиболее обычны следующие виды: Dryopteris cristata, Athyrium filixfemina, Pteridium aquilinum, Pinus sylvestris, Pricea abies, Milium effusum, Lerchenfeldia flexuosa, Festuca altissima, Luzula palescens, Majanthemum bifolium, Polygonatum multiflorum, Paris quadrifolia, Salix aurita, S. caprea, Populus tremula, Betula pendula, Quercus robur, Ribes nigrum, Oxalis acetosella и др.
23. Относительно высокие песчаные холмы и гряды – остатки древних дюн, которые образовались значительно раньше Куршской косы. Сухие разнотравно-кустарничковые зеленомошные сосновые леса. Почвы песчаные оподзоленные. Увлажнение нормальное, периодически слегка недостаточное из-за интенсивного дренажа. Здесь обычны : Dryopteris cristata, Athyrium filix-femina, Pteridium aquilinum, Pinus sylvestris, Picea abies, Lerchenfeldia flexuosa, Festuca altissima, Majanthemum bifolium, Paris quadrifolia, Populus tremula, Corylus avelanna, Quercus robur, Ulmus laevis, Ribes pubescens, Sorbus aucuparia, Rubus idaeus, Fragaria vesca, Geumurbanum, Prunus padus, Oxalis aceyosella, Euphorbia cyparissias, Frangula alnus, Viola canina, Calluna vulfaris, Vaccinium myrtillus и др.
30. Невысокие легко суглинистые и супесчаные холмы и
гряды. Ельники кисличные и кислично-зеленомошные. Почвы подзолистые.
Увлажнение нормальное. Здесь наиболее обычны: Dryopteris cristata,
Athyrium filix-femina, Lycopodium clavatum, Picea abies, Lerchenfeldia
flexuosa, Festuca altissima, Majanthemum bifolium, Polygonatum
multiflorum, Paris quadrifolia, Betula humilis, Quercus robur,
Moehringia trinervia, Sorbus aucuparia, Rubus idaeus, Chamaenerion
angustifolium, Ledum palustre, Vaccinium uliginosum и др.
24. Просеки среди широколиственных и
хвойно-широколиственных лесов. Первая стадия зарастания. Увлажнение
нормальное. Почвы суглинистые и супесчаные оподзоленные. Здесь обычно
встречаются Pteridium aquilinum, Pinus sylvestris, Picea abies,
Lerchenfeldia flexuosa, Salix caprea, Betula pendula, Quercus robur,
Arenaria serpyllifolia, Sorbus aucuparia, Rubus idaeus, Medicago
falcata, Vicea sepium, Oxalis stricta, O. acetosella, Frangula alnus,
Trientalis europaea, Galeopsis speciosa, Melampyrum nemorosum, Vaccinium
myrtillus, V. vitis-idaea, Trifolium hybridum, Medicago lupulina, Urtica
dioica, Chamaenerion angustifolium и др. В этом классе местообитаний
основными факторами изменения растительности являются интенсивность
увлажнения и характер подстилающих пород.
X. Класс местообитаний гидрофильной растительности
черноольховых лесов
6. Широкие и глубокие осушительные каналы в топких
черноольховых лесах. Водная растительность. Субстрат торфянистый,
уровень минерализации очень слабый. Выделяются в отдельный класс
местообитаний. Видовой состав растительности очень бедный. Здесь
встречаются: Lemna minor, Nuphar lutea, Nymphaea candida, Hydrocharis
morsis-ranae.
XI. Класс местообитаний олиготрофных сфагновых болот
29. Верховое сфагновое болото с сосной и березой. Обширный
участок олиготрофного болота расположен на невысокой первой надпойменной
террасе р. Неман. Нанорельеф мелкобугорковатый. Древостой находится в
угнетенном состоянии. Здесь встречаются : Pinus sylvestris, Scheuchzeria
palustris, Eriophorum polystachyon, E. vaginatum, Eleocharis palustris,
Betula humilis, B. pubescens, Droseraanglica, D. rotundifolia, Rubus
chamaemorus, Empetrum nigrum, Ledum palustre, Andromeda polifolia,
Calluna vulgaris, Oxycoccus quadripetala, Vaccinium uliginosum. Это
самостоятельный флористически очень своеобразный класс местообитаний.
Несомненно, нуждается как в тщательном изучении, так и в охране.
Выявленная флористическая структура растительного покрова
показывает, что основные градиенты растительности на
ландшафтно-экологическом уровне в этом районе зависят в первую очередь
от интенсивности и характера увлажнения, уровня трофности и степени
дренированности субстрата. Однако заметную роль в дифференциации
растительного покрова играют антропогенные воздействия, такие, как
вырубка леса, распашка, осушение, создание польдерного режима водного и
минерального питания, выпас скота, посев многолетних трав и монокультур,
насыпание дамб и полотен дорог. Примечательно, что поля с
сельскохозяйственными культурами флористически слабо отличаются от
польдерных лугов и пастбищ, образуют с ними один класс
местообитаний.Естественные же или близкие к ним по степени
выработанности ценотической структуры искусственные луга, но давно не
перепахиваемые, образуют самостоятельный класс местообитаний.

|
К сожалению, луга с выработанной ценотической структурой в данном районе встречаются небольшими фрагментами и занимают небольшую площадь.Флористически весьма рыхлый класс местообитаний образуют типы местообитаний нарушенной части дельтовой низменности. Это связано с тем, что растительность этих участков в наименьшей степени подвержена антропогенному воздействию и близка к климаксовому состоянию, т. е. имеет хорошо выработанную ценотическую структуру. Эта растительность находитсяв биогеохимическом равновесии с внешней средой. В этом состоянии растительный покров, вероятно, более «чувствителен» к изменениям внешней среды, и на экологоценотическом уровне такой растительной покров оказывается более тонко организованным. В ненарушенном состоянии флористические градиенты растительности более адекватны экологическим градиентам среды. Это, например, наблюдается в классах местообитаний VIII и IX, где растительность близка к естественному состоянию, менее нарушена человеком.
|
Следует отметить, что сильное окультуривание ландшафта при разумном сохранении участков с практически нетронутой растительностью увеличивает, а не снижает общее экологическое разнообразие ландшафта. В нашем случае воздействие человека привело к образованию не менее 5–6 новых классов местообитаний, не свойственных данному ландшафтному району в его первозданном виде.
Флористическая общность типов местообитаний, возникших в
связи с антропогенным воздействием, между собой значительно ниже, чем
типов местообитаний, занятых естественной растительностью, что
несомненно связано с так называемой синантропизацией конкретной флоры,
появлением в ней комплекса видов, приуроченных к трансформированным
местообитаниям. Синантропными видами, как правило, становятся виды,
приуроченные к участкам с нарушенным растительным покровом, ценотическая
структура которого почти не выработана. В условиях антропогенного
нарушения, благодаря широкой физиологической амплитуде, эти виды
становятся наиболее активными и резко увеличивают свою встречаемость.
Напротив, многие виды, часто встречающиеся в растительном покрове не
нарушенном человеком, с выработанной ценотической структурой, при
разрушении этой структуры становятся редкими и постепенно исчезают
совсем.
Таким образом, не сохранив всех форм ценотической
организации растительного покрова (не сохранив растительность), мы не
можем сохранить и его флору. Эта очевидная истина, к сожалению, все еще
очень слабо учитывается при планировании природоохранных мероприятий и
разработке сетей охраняемых территорий.
Читать следующий раздел. Фотографии взяты из альбома Neregeta Lietuva (Невиданная Литва) Фотограф Marius Jovaisa http://fotki.yandex.ru/users/tomcat11/ или http://www.liveinternet.ru/users/teyty/post123136921/

|
