В предыдущих разделах с помощью графов флористического сходства местообитаний и типов местообитаний изображена ландшафтно-экологическая структура растительного покрова нескольких районов северной Евразии. Мы считаем, что флористическая структура растительного покрова, понимаемая как система отношений таксономического сходства - различия, выявленная на множестве фитоценозов или их типологических объединений в пределах одного элементарного ландшафтного района с одной конкретной флорой, в немалой степени адекватна интегральной экологической структуре этого района в понимании В. Б. Сочавы (1978).
Математически графы представляют собой абстрактные конструкции, состоящие из множества вершин (в нашем случае фитоценозов, местообитаний или типов местообитаний), на которых распределено множество ребер (в нашем случае отношений флористического сходства выше определенного заданного уровня). Каждое множество типов местообитаний, а в окрестностях г. Сыктывкара конкретных местообитаний, довольно равномерно охватывает все основное разнообразие экологического спектра соответствующих ландшафтных районов независимо от того, на каких площадях выражены представленные сочетания экологических факторов. Выявлены структуры растительного покрова разных районов, отличающихся по геоморфологическому строению (горы и равнины), по интенсивности антропогенной трансформации (заповедник и типичные аграрные районы), по зональному положению (тундра, северная тайга, хвойно-широколиственные леса), а также по таксономической и ареалогической структуре конкретных флор.
Уже при описании растительного покрова мы убедились, что метод графов - весьма рациональный способ подачи и анализа структуры сложных многомерных объектов, каким является растительный покров. В последнее время этот метод все чаще используется в геоботанических, ботанико-географических, зоогеографических и этологических исследованиях (Галанин, 1989, 1990). Например, с помощью графов растительный покров можно представить как некоторый пространственный процесс (Роману, 1984; Галанин, 1989, 1990) в абстрактном экологическом пространстве. Растительный континуум легче понимать и анализировать, пользуясь представлениями о пространственных процессах. Однако по-прежнему и в геоботанике, и в зоогеографии при изучении структуры растительного покрова и животного населения преобладают попытки выявить и проанализировать закономерности организации путем типизации и классификации отдельных пространственных состояний объекта, вырезанных из его непрерывной ткани. В настоящей работе мы также сполна отдали дань этому традиционному методу познания, выделяя типы, классы, подклассы и надклассы местообитаний. Разумеется, традиционный способ зализа структуры растительного покрова дает немало, но все же многие свойства остаются как бы «за кадром», в частности такой вопрос, как соотношение дискретности и непрерывности в структуре растительности и растительного покрова. Мы мало задумываемся о том, может ли быть континуальная структура иерархичной, или это свойство только дискретных структур. Наконец, открытым, на мой взгляд, остается вопрос о том, исчерпывается ли разнообразие всех пространственных процессов или континуальных структур шкалой «дискретность -континуальность». Подобные вопросы не схоластические, как может показаться на первый взгляд. Было бы чрезвычайно интересно выяснить, какая структура, дискретная или континуальная, экологически более оптимальна и как это зависит от антропогенного воздействия или климатических вменений.
В целях наглядности можно провести аналогию между изменениями в пространстве и изменениями во времени и представить, что изменения во времени (временные процессы) исследуются и анализируются только с помощью методов дискретного анализа состояний системы, а именно так в подавляющем большинстве случаев изучаются сегодня изменения в пространстве (пространственные процессы). Например, в «процессе движения Земли вокруг Солнца» можно выделить типов и 2 класса состояний. Условно каждый класс можно разбить на 2 подкласса. Анализируя выделенную структуру, мы можем выяснить, что каждый тип, подкласс и класс состояний характеризуются определенным режимом солнечной энергии, приходящейся на единицу поверхности, определенным состоянием в циркуляции атмосферы Земли, определенной температурой воздуха у ее поверхности и т. д. Такой подход, несомненно, позволит вскрыть некоторые закономерности процесса движения Земли, однако «за кадром» останутся вопросы скорости и причинах движения, гравитации, массе Земли массе Солнца, расстоянии между ними, эволюции системы Солнце - Земля и др. Периодическая система химических элементов Д. И. Менделеева чрезвычайно полезна как классификация, но ее значение неизмеримо выросло по мере того, как она стала рассматриваться в качестве процесса трансформации и перехода химических элементов, т. е. в качестве временного процесса. В биологии мы также знаем переход от искусственных систем классификации видов животных и растений (типичных классификаций) к филогенетическим классификациям - к временным процессам.
При изучении ландшафтно-экологической структуры растительного покрова, животного населения и всей биоты в целом непрерывность следует рассматривать, вероятно, не как нежелательную помеху, мешающую выделять четкие типы и классы состояний пространственного процесса, а как важнейшее свойство этого пространственного процесса, имеющее фундаментальное экологическое значение. К сожалению, арсенал методов работы с континуальными структурами (пространственными процессами) сегодня невелик и в основном сводится к построению графов, дендритов, к нахождению числа осей пространства ординации и определению разнообразия состояний в направлении этих осей.
Вместе с тем некоторые свойства графов как моделей пространственных процессов можно анализировать с помощью вероятностных методов (Галанин и др., 1986; Галанин, 1989; Галанин, Носов, 1989). Оказалось, что по аналогии с временными процессами пространственные процессы могут быть равномерными и неравномерными, линейными и циклическими. Одним словом, разнообразие структуры графов нельзя сводить лишь к разному сочетанию дискретности и непрерывности.
Рассмотрим некоторый граф G(V, Е), состоящий из множества вершин V и множества ребер Е. Наш граф неориентирован, так как ребра не имеют направления, и порядок следования вершин в паре, соединенной одним ребром, не имеет значения. Ребро обозначает симметричное отношение двух вершин. Ориентированные графы вместо ребер имеют дуги, которые от ребер отличаются тем, что обозначают несимметричные отношения между вершинами, такими отношениями, например, в геоботанике и флористике могут быть отношения включения. В отличие от ребер, дуги на графах обозначаются стрелками. Граф, содержащий только ребра, называется неориентированным, а граф, содержащий дуги, - ориентированным. В случае попарного флористического сходства, по Жаккару, мы имеем неориентированные графы.
Две вершины графа, соединенные одним ребром, называются смежными. Ребра, имеющие общую вершину, т. е. исходящие из одной вершины, тоже называются смежными. Ребро и любая из его двух вершин инцидентны друг другу. Обычно ребра обозначаются через перечисление двух вершин, соединенных ребром. Так, если ребро связывает вершину u с вершиной v, то оно обозначается (u,v).
Каждый граф можно представить в евклидовом пространстве множеством точек или кружков, которые соответствуют вершинам, и множеством ребер и отрезков, соединяющих определенные пары вершин. Существуют различные способы задания графа.
Пусть u1, u2, ..., un - вершины графа G (v, е), а е1, е2, ..., еm - его ребра. Матрицей смежности,
соответствующей графу G (v, е), называется матрица А=аij, у которой элемент aij равен числу ребер, соединяющих вершины
ui и uj и аij=0, если соответствующие вершины не смежны. В нашем случае матрицы попарного флористического сходства местообитаний
являются матрицами смежности.
В матрице инцидентности В=bij графа G(v, u) элемент bij= 1, если вершина ui инцидентна ребру еj и bij= 0,
если вершина ui и ребро еj не инцидентны. В нашем случае, при анализе флористического сходства, матрицы инцидентности не используются.
Вообще граф G(v,е) можно задать посредством списка всех пар вершин, соединенных ребрами, или путем задания для каждой вершины множества смежных с ней вершин. Например, граф флористического сходства типов местообитаний района Прегольской низменности в этом случае запишется так:
вершины, связанные ребрами:
(1,2) (1,3) (2,3) (1,4) (3,4) (3,15) (16,23) (23,24) (22,23) (22,24) (21,22) (21,25) (22,25) (1-13) (4-14) (12,13) (21,26) (26,27) (6,27) (13,14) (5,13) (6,13) (6,14) (7,14) (12,14) (5,14) (5,7) (16,7) (5,12) (1,14) (4,12) (5,6) (5,8) (6,12) (6,8) (12,9) (12,8) (8,10) (13,10) (8,13) (7,8) (9,8) (10,18) (8,11) (9,11) (4,13) (4,5)
вершины со смежными исходящими ребрами:
1,2,3,4
2,1,3
3,2,1,4,15
4,1,3,13,14,12,5
13,1,4,14,12,7,8,10,6
12,5,14,4,13,6,8,9 8,19,6,7,13,5,12,9
9,12,7,13,5,12,9
9,12,7,8,11
10,13,8,18
11,9,8
18,10
27,6,26,25
26,27,21 |
14,4,1,13,6,7,12,5
6,13,14,6,7,12,8,27
7,13,14,6,5,9,8
5,4,14,13,6,7,8,12
25,27,21,22
21,26,25,22,23
22,25,21,23,24
23,21,22,24,16
24,22,23
16,23
15,3
17
19
20 |
Два графа G(v, е) и Н(w, j) называются изоморфными, если cуществует взаимно однозначное соответствие между множествами вершин V, W и множествами ребер Е, J,
сохраняющее отношение инцидентности. Подграфом G'(v', е') графа G (v, е) называется граф с множеством вершин V'<V и множеством ребер Е'<Е, каждое из которых инцидентно только вершинам из V'. Подграфом G' (v', е'), порожденным подмножеством V'<V, называется граф с множеством вершин V' и набором ребер Е', состоящим из всех ребер графа G, которые соединяют вершины V'. Остовный подграф G'(v', е') содержит все вершины графа G и некоторый набор его ребер Е'—Е. В нашем случае остовным графом относительно данного графа может быть граф, полученный при повышении порога сходства, когда число вершин остается прежним, а число ребер уменьшается, так как часть связей оказывается меньше нового порогового значения.
Последовательность ребер графа типа (u0, u1), (u1, u2), ..., (ui-1, uj),
(ui, ui-1), ..., (ur-1, ur) называется маршрутом, соединяющим вершины u0 и ur. Маршрут замкнут,
если u0=ur. Маршрут называется цепью, если все его ребра различны, и простой цепью, если все его вершины различны. Замкнутая простая цепь
называется простым циклом. Примером маршрута, соединяющего вершины 22 и 23 на графе (рис. 6, Б), будет 22—29—28—23. Это кратчайший маршрут, соединяющий эти две вершины, которые соединяет и более длинный маршрут: 22—29—18—26—21—24— 23. Приведем пример простой замкнутой цепи из того же графа: 29—18—26—21—28—29.
Граф называется связным, если любая пара его вершин соединена хотя бы одним маршрутом со всеми остальными вершинами, в таком графе нет изолированных отдельных вершин или групп вершин - подграфов. Максимальный связный подграф графа G называется компонентой связности. Несвязный граф имеет, по крайней мере, две компоненты связности.
Длина маршрута от вершины ui до вершины uj цепи, простой цепи, цикла или простого цикла равна количеству ребер в порядке их прохождения.
Длина кратчайшей простой цепи, соединяющей вершины ui и uj в графе G, называется расстоянием d(ui, uj) между
ui и uj. В связном неориентированном графе расстояние удовлетворяет аксиомам метрики. Диаметр графа - это его наибольшее расстояние.
Величина min max d(ui, uj) называется радиусом, а вершина uo, для которой mах d(ui, uj) принимает
наименьшее значение, называется центром графа. В несвязном графе может быть много центров, или ни одного, если все вершины изолированы друг от друга, т. е.
когда ребра отсутствуют.
Степенью вершины ui графа G, обозначаемой di, называется число ребер, инцидентных этой вершине. Например, на графе - модели структуры растительного покрова Чаунского стационара - степень вершины 29 d29=6, а степень вершины 7 d7= 1.
Если граф G имеет n вершин и m ребер, то
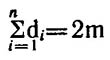
Вершина ui называется изолированной, если di= 0, и концевой, если di= 1. Граф, у которого все вершины имеют одинаковые
степени, равные k, называется регулярным степени k. В полном графе каждая пара вершин соединяется одним ребром. Для графа G(V,Е), не имеющего кратных ребер,
дополнительным графом будет граф G1(V1, Е1) у которого V1=V и вершины смежны в G только в том случае,
когда они не смежны в G1. Граф, дополнительный к полному графу, состоит из изолированных вершин и называется пустым.
Многие характеристики для графа G и его дополнения G1 зависимы.
Каждому графу G можно отнести ряд графов, являющихся производными от G. Так, реберным графом L(G) графа G называется граф, вершины которого соответствуют ребрам графа G и две вершины смежны в L(G) в том и только в том случае, когда соответствующие им ребра графа G смежны. В тотальном графе Т(G) графа G вершины соответствуют элементам графа G, т. е. вершинам и ребрам, и две вершины в Т (G) смежны тогда и только тогда, когда соответствующие элементы в G смежны или инцидентны. Многие свойства графа G при этом переносятся на графы L(G) и Т (G).
С помощью различных операций можно строить граф из более простых графов, переходить от одного графа к более простому, разбивать граф на более простые, в заданном классе графов переходить от одного графа к другому. Наиболее употребительны операции на графах - удаление ребра или нескольких ребер, при этом вершины удаленного ребра или ребер сохраняются; добавление ребра между двумя вершинами графа; удаление вершины с инцидентными ей ребрами (граф, полученный путем удаления вершины v из графа G, обозначается (G—v)); добавление вершины, которую можно соединить ребрами с некоторыми вершинами графа; стягивание ребра - отождествление пары смежных вершин, т. е. удаление пары смежных вершин и добавление новой вершины, смежной с теми вершинами G, которые были смежны хотя бы с одной из удаленных вершин; подразбиение ребра и добавление новой вершины, которая соединяется ребром с каждой вершиной удаленного ребра. Одним словом, теория графов предлагает ряд методов и приемов, которые с успехом можно использовать при анализе структуры растительного покрова.
МЕТОДИКА
Пусть некоторый граф G, моделирующий флористическую структуру растительного покрова, имеет n вершин (u1, u2, u3,
..., un). Все вершины пронумерованы, и мы имеем дело с помеченным графом. На множестве из n вершин распределено множество из m ребер
(е1, е2, е3, ..., еm). Структура графа определяется характером распределения ребер по вершинам, следовательно, характеристика этой структуры должна быть характеристикой этого распределения.
При случайном независимом распределении каждое конкретное ребро еj может занять с равной вероятностью одно из n(n—1)/2 положений, возможных на данном графе, исходя из числа его вершин. Здесь n(n—1)/2 — число всех возможных сочетаний п вершин графа по две, т. е. максимальное число ребер для графа с п вершинами, чтобы сделать его полным графом, когда каждая его вершина связана со всеми остальными вершинами. Если m = n(n—1)/2, то мы имеем полный граф, к которому понятие структуры в нашем смысле вообще неприменимо. Следовательно, отношение 2m/n(n—1) можно использовать как показатель степени вершины полного графа.
Вероятность того, что при случайном независимом распределении ребер по множеству вершин определенное ребро еk займет определенное положение
(u1, u2), будет равна
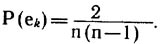
Вероятность же того, что одно из m ребер (любое) займет определенное положение (ui, uj), будет в m раз больше:
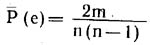
Вероятность обратного события Р (е), когда ни одно из m ребер не займет определенного положения (ui, uj), можно рассчитать
из соотношения
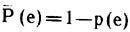
Из определенной вершины в предельном случае может исходить (n—1) ребро, это случай полного графа. Следовательно, среднее ожидаемое число ребер
на одну вершину в данном графе G (число ребер, в среднем инцидентных одной вершине), ожидаемое при случайном независимом распределении ребер, будет равно
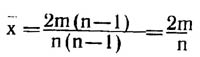
Следовательно, в равномерном графе с n вершинами и m ребрами из каждой вершины должно исходить по 2m/n ребер. В этом случае мы можем говорить о регулярном графе или об абсолютно равномерном пространственном процессе, моделируемом этим графом, который можно назвать равномерным континуумом. Равномерный континуум - это аналог равномерного прямолинейного движения или равномерного процесса во времени. При работе с такими графами использование методов дискретного анализа мало эффективно, так как группы вершин, выделенные в виде типов или классов, не будут естественными, реально существующими, эти группы будут искусственно вырезанными из непрерывной ткани равномерной континуальной структуры.
Графовые модели флористической структуры растительного покрова основаны на выборочных данных. Это - совокупности геоботанических и флористических
описаний пробных площадей, Заложенные в пределах ландшафтного района. В такую выборку могут попасть не все типы местообитаний, и поэтому экологическое
разнообразие района будет представлено неравномерно. При обследовании геоботанических площадей могут быть пропущены виды, могут быть ошибки
в определениях видов, что также отразится в структуре полученного графа. Таким образом, даже при равномерном растительном континууме в связи
с перечисленными моментами в структуре полученного графа возможны случайные отклонения от идеальной модели, при которой степень всех вершин
одинакова и из каждой вершины исходит по 2m/n ребер. В стохастической модели при среднем количестве ребер, исходящих из одной вершины, равной
(х=2m/n), могут быть вершины, из которых исходит большее и меньшее количество ребер. В этом случае степень вершин di - случайно варьирующая величина.
Обозначим возможные значения степени вершины di графа G как события, когда из вершины исходит определенное количество ребер.
При этом полное поле всех возможных событий будет
do, d1, d2, d3, ..., dk, ..., dn-1.
Здесь do — событие, когда вершина полностью изолирована и из кее не исходит ни одного ребра; d1 — из вершины исходит одно ребро
(вершина концевая); d2 — из вершины исходит два ребра {степень вершины равна двум); dk — из вершины исходит k ребер
(степень вершины равна k); dn-1 — из вершины исходит максимально возможное для данного графа число ребер
(вершина соединена ребрами со всеми остальными вершинами данного графа; в неполном графе эта величина не может быть больше m, тогда все ребра исходят из
одной вершины). Событие dy состоит в том, что из вершины исходит среднее ожидаемое число ребер 2m/n. Вероятность этого события должна быть максимальной, отклонения от нее вправо и влево по оси значений должны быть менее вероятными, причем чем больше отклонение от среднего, тем меньше его вероятность. Для равномерного графа со случайными отклонениями распределение вероятностей вершин разной степени должно подчиняться биномиальному закону.
Зная вероятность прямого и обратного события Р(е) и P(е), мы можем построить модель биномиального распределения вероятности событий
do, d1, d2, ..., dn-1. Вероятности событий находятся по разложению бинома Ньютона (р—q)n:
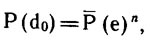
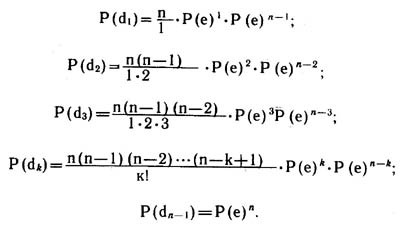
Зная вероятности событий, мы можем рассчитать ожидаемые абсолютные значения числа вершин с той или иной степенью этих вершин (тем или иным числом исходящих из них ребер):
хo = Р(do) • n; x1 = Р(d1) • n; хk = Р(dk) • n.
Полученные результаты удобно изображать в виде графиков статистических распределений числа вершин данного графа в зависимости от степени этих
вершин, т. е. в зависимости от числа исходящих из них ребер. Сопоставление эмпирического ряда частот с теоретическим биномиальным позволяет
ответить на вопрос о том, является ли структура графа регулярной со случайными отклонениями и мы имеем дело с равномерным пространственным
процессом, или же она не регулярна и характеризуется неравномерным распределением ребер по вершинам. Достоверность отклонения эмпирического
распределения от его теоретической биномиальной модели можно оценить с помощью критерия Пирсона х2:
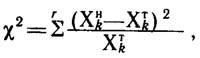
где x2 - величина критерия Пирсона; хнk - число вершин с k исходящими ребрами, наблюдаемое; хтk
- число вершин с k исходящими ребрами, ожидаемое при условии случайного биномиального распределения; т - число сравниваемых образцов или классов
распределения.
В результате распределения ребер по вершинам графа в его структуре в принципе могут образоваться элементы разного типа: связные
компоненты, подграфы, изолированные вершины или изоляты, из которых не исходит ни одного ребра. Следует заметить, что изоляты могут
возникать при случайном распределении ребер по вершинам, когда 2m/n<1. Следовательно, наличие в структуре графа связных компонент, подграфов и изолятов само по себе еще не говорит о дискретности изучаемой структуры. При наличии подграфов и изолятов мы можем говорить о дискретности структуры растительного покрова только в том случае, когда число наблюдаемых подграфов и изолятов достоверно больше ожидаемого при случайном распределении ребер по вершинам.
Простейшим типом связных компонент являются изолированные пары вершин, соединенные между собой одним ребром. На графиках распределения частот вершин в зависимости от степени или числа исходящих из них ребер преобладание структур типа «гантелей» будет выражаться в явном преобладании над ожидаемым числом вершин с одним исходящим ребром. Если в структуре графа «гантелей» больше, чем можно ожидать при случайной модели, то такой пространственный процесс также можно классифицировать как дискретный.
Второй тип связных компонент графа - «звезды»- характеризуется тем, что несколько вершин с одним исходящим ребром связано с одной центральной вершиной, из которой ребер исходит столько, сколько с ней связано вершин. На графиках распределения в этом случае будет наблюдаться явное преобладание вершин с одним исходящим ребром при значительно меньшем количестве вершин с двумя, тремя и более исходящими ребрами. Однако в этом случае число вершин с многими исходящими ребрами (центры «звезд») также будет больше ожидаемого, кривая распределения в предельном случае графа со «звездами» будет двувершинной. Граф, распадающийся на «звезды», если их число больше ожидаемого, также будет отвечать дискретной структуре моделируемого им растительного покрова.
Если связными компонентами графа являются объединения вершин типа линейных последовательностей, «цепочек», «петель» и «треугольников», то на графике распределения будут явно преобладать вершины с двумя исходящими ребрами. Чем больше на графе двухреберных вершин по сравнению с однореберными, тем регулярнее структура изучаемого объекта. Связные компоненты графа типа «плеяд» на графике распределения вершин проявляют себя повышенным числом вершин с очень большим количеством исходящих ребер при заниженном числе вершин с одним и двумя исходящими ребрами. Если тип структуры графа сетчатый, то должно быть повышенное число трех-четырехреберных вершин. Распознавание формы связных компонент графа и их классификация несомненно имеют большое значение при анализе изучаемых структур.
При построении графов нами были использованы флористические связи разного уровня. В связи с этим чрезвычайно интересно выяснить, на каком уровне сходства-различия дискретность и нерегулярность структуры выявляются наиболее полно. Для каждого графа строились теоретические и эмпирические распределения частот вершин разной степени связности при разных ступенях сходства-различия. Учитывая только тесные связи, мы получаем графы с малым числом ребер. На таких графах даже при случайном распределении ребер будет много связных компонент и изолированных вершин. Учитывая наряду с тесными и слабые флористические связи, увеличиваем число ребер и снижаем таким образом вероятность существования случайных изолированных вершин. Связность графа при этом будет всегда увеличиваться, но степень регулярности может оставаться без изменений или даже снижаться.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
С помощью описанной выше методики анализировались графы флористического сходства местообитаний и типов местообитаний семи ландшафтных районов. Результаты представлены в виде таблицы (табл. 1) и графиков (см. ниже).
 |
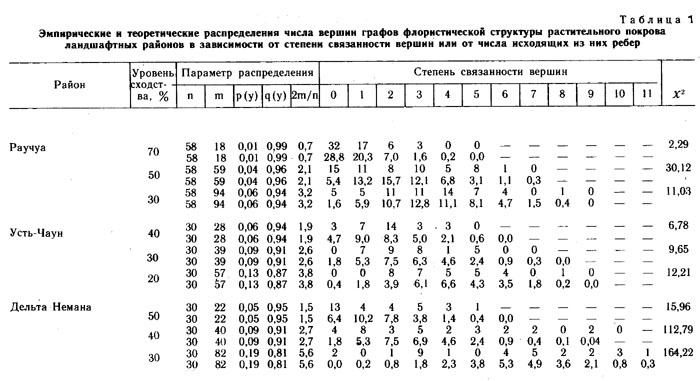
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района реки Раучуа (а - уровень сходства типов местообитаний более 70%, б - более 50%, в - более 30%).
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
На рис. слева изображены кривые распределения числа вершин графа флористического сходства типов местообитаний района Раучуа. Учитывались только ребра, соответствующие флористическому сходству более 70%. При числе вершин (n = 58) и числе ребер (m =18) велика вероятность существования в структуре графа «изолятов» и «гантелей». Эмпирическое распределение вершин в зависимости от числа исходящих ребер (в зависимости от степени смежности вершин) от теоретического отличается очень слабо (х2 = 2,29). Следовательно, учитывая только уровень сходства более 70%, мы получаем граф с регулярной структурой, который моделирует пространственный процесс, весьма близкий к равномерному. Такой тип структуры можно охарактеризовать как равномерный стохастический континуум. |
Приняв более низкий уровень флористического сходства (более 50%), увеличиваем число ребер до 59, в среднем на одну вершину теперь приходится 2,1 ребра, а не 0,7, как в предыдущем случае. Структура графа (рис. б) теперь явно дискретна, так как число изолятов значительно выше, чем ожидается теоретически. Кроме того, значительно больше теоретически ожидаемого число вершин с пятью исходящими ребрами. Структура графа неравномерна. Пространственный процесс, моделируемый этим графом, имеет явно неравномерный характер (х2 = 30,12).
При уровне флористического сходства больше 30% (рис. в) эмпирическая кривая отличается от теоретической (х2 =11,03), но меньше, чем при уровне сходства более 50%. В этом случае можно говорить о слабой неравномерности структуры графа. Дискретности на этом уровне сходства не обнаруживается, хотя вершин с четырьмя, пятью и шестью исходящими ребрами больше, чем ожидается при случайном равномерном распределении.
Причина флористической неравномерности структуры растительного покрова района Раучуа, по-видимому, связана с контрастностью рельефа и резкой континентальностью климата. В таких условиях участки с разным типом экологического режима оказываются флористически резко разграниченными, экологические градиенты или барьеры оказываются столь большими, что они непреодолимы для многих видов конкретной флоры. В структуре графа хорошо заметна иерархичность - прослеживается несколько уровней скоплений типов местообитаний, что также отражается в структуре кривых распределения вершин в зависимости от числа исходящих из них ребер.
Район Раучуа, как многие ландшафтные районы Западной Чукотки, характеризуется давно сложившейся флорой и растительностью, развитие которых в основном происходило преемственно на протяжении нескольких геологических эпох начиная с плиоцена. В этом случае эколого-ценотические амплитуды большинства видов пришли в соответствие с градиентами экологической структуры ландшафта. Дискретность растительного покрова может иметь и ценотический характер, когда в результате межвидовой конкуренции виды как бы «загоняются» в оптимальные для них местообитания и «изгоняются» из местообитаний неоптимальных.
Наиболее оптимален для работы в районе Раучуа уровень флористического сходства типов местообитаний выше 50%, так как разрешающая способность метода в этом случае самая высокая. Говоря о неравномерности и дискретности растительного покрова, выявляемой с помощью расчета флористического сходства, следует всегда отмечать, на каком уровне сходства она выявляется наиболее четко. В связи с-этим есть основание оценивать не только степень неравномерности структуры, но и уровень и глубину этой неравномерности.
 |
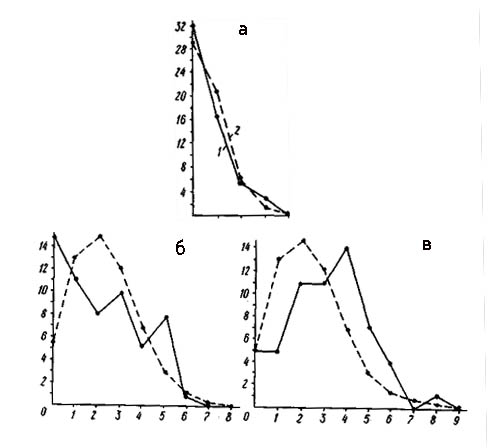
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района Усть-Чаунского стационара (а - уровень сходства более 40%, б - более 30%, в - более 20%).
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
На рис.слева изображены кривые распределения вершин в зависимости от числа исходящих ребер графа флористического сходства типов местообитаний района Усть-Чаунского стационара: а - флористическое сходство, по Жаккару, более 1%. б - флористическое сходство более 30%, в - флористическое сходство более 20%. При уровне сходства более 40% 28 ребер приходится 30 вершин, в среднем по 1,9 ребра вершину. Распределение вершин и ребер в этом случае отличается от случайного незначительно (х2 =6,78), правда, при этом имеет место достоверное превышение над ожиданием фактического числа вершин с двумя исходящими ребрами, говорит о сильно выраженной равномерности структуры графа, так как среднее число ребер на вершину в этом графе равно примерно двум. Неравномерность структуры растительного покрова при таком уровне флористического сходства фактически не выражена.
При уровне сходства более 30% (рис. б) эмпирическое распределение становится почти прерывистым. Здесь больше ожидаемого вершин с одним, двумя, тремя и пятью исходящими ребрами, но много меньше ожидаемого вершин с четырьмя ребрами. В целом этот связный граф моделирует сильно неравномерный пространственный процесс (х2 =9,65). На этом примере мы можем убедиться, что при очень высокой связности структуры графа она может быть сильно неравномерной.
При уровне сходства не более 20% (рис. в) эмпирическая кривая еще более достоверно отличается от теоретической (х2= 12,21), однако при этом сильно увеличивается связность графа, так как вершин с двумя исходящими ребрами значительно больше ожидаемого. Структура графа, а следовательно, и моделируемого этим графом растительного покрова на данном уровне флористического сходства сильно равномерна. Несмотря на то что при учете связей выше 40% множество вершин графа распалось на 8 изолированных подмножеств (компонент), мы можем говорить только об очень слабой неравномерности этой структуры. Это связано с тем, что при малом среднем числе ребер на вершину вероятность случайного распада графа на связные компоненты и изоляты очень велика, а порой просто даже неизбежна - ребер не хватает, чтобы связать все вершины. Сравнивая графики распределений, можно заметить, что наиболее полно и четко неравномерность структуры растительного покрова района Усть-Чаунского стационара выявляется на уровне сходства более 30%. Именно этот уровень был выбран нами при выделении классов местообитаний.
|
Сравнивая флористические структуры растительного покрова районов Раучуа и Усть-Чаунского стационара, можно заметить, что последняя характеризуется значительно более высокой связностью и равномерностью, что несомненно связано с меньшей контрастностью рельефа района Усть-Чауна и меньшей контрастностью местообитаний. Связность структуры растительного покрова в окрестностях стационара, вероятно, может усиливаться за счет постепенного изменения степени затопляемости дельты солеными водами по мере удаления от побережья залива, а этот фактор играет здесь в распределении растений значительную роль.
 |
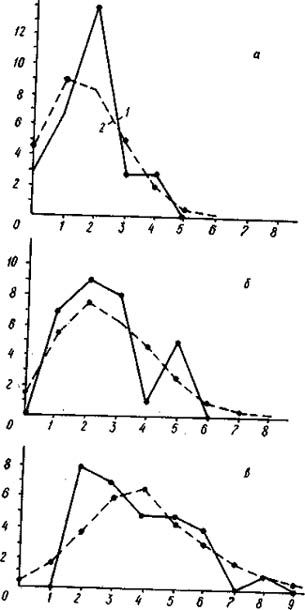
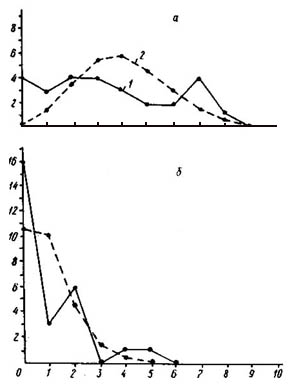
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района дельты р. Немана (а - уровень сходства более 50%, б - более 40%, в - 30%).
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
На рис. слева изображены эмпирические и теоретические распределения вершин графа флористического сходства типов местообитаний района дельты Немана. Отчетливо видно, что при всех ступенях сходства эмпирические распределения резко отличаются от теоретических. При самом высоком уровне сходства - более 50% (рис. а) - граф имеет 22 ребра при среднем числе ребер на одну вершину 1,5. Структура дискретна, так как «изолятов» значительно больше, чем ожидается при случайном распределении, а вершин с одним и двумя ребрами меньше ожидаемого. Структура неравномерна, и степень этой неравномерности весьма значительна (х2 = 15,96), что проявляется не только в ее дискретности, но и в наличии высоко связных компонент.
При снижении уровня сходства до 40% (рис. б) число ребер графа увеличилось до 40 при среднем числе ребер на одну вершину 2,7. Эмпирическое распределение в этом случае резко отличается от теоретического (х2 = 112,79). Видно, что неравномерность структуры графа при снижении порога флористического сходства значительно усилилась. Структура дискретна, так как «изолятов» значительно больше ожидаемого. Больше ожидаемого и концевых вершин с одним ребром. Кроме того, больше ожидаемого вершин с семью и шестью исходящими ребрами, даже имеются две вершины с девятью исходящими ребрами.
Наибольшей величины неравномерность флористической структуры растительного покрова района дельты Немана достигает при учете связей более 30% (рис. в). Число ребер графа увеличилось до 82 при среднем числе ребер на вершину 5,6. При очень высокой насыщенности графа ребрами мы имеем весьма неравномерную структуру, в которой имеются даже «изоляты», что при таком высоком числе ребер очень маловероятно. Больше ожидаемого на графе вершин с тремя, десятью и одиннадцатью исходящими ребрами. |
Вместе с тем на графе отсутствуют вершины с одним ребром и вершины с пятью исходящими ребрами. Таким образом, пространственный процесс, моделируемый данным графом, следует считать крайне неравномерным, причем глубина дискретности и неравномерности очень велика и прослеживается при всех трех уровнях сходства, и даже при снижении уровня сходства и увеличении числа ребер в графе неравномерность его структуры не только не снижается, а, напротив, возрастает.
Резкое отличие флористической структуры растительного покрова района дельты Немана от структуры районов Раучуа и Усть-Чаунского стационара объясняется мощным антропогенным преобразованием растительного покрова, его флоры и ландшафта в целом. Типы местообитаний здесь четко paспадаются на несколько подмножеств (связных компонент), каждое из которых имеет свою степень связности. Именно это и придает графу высокую неравномерность. Наиболее высока степень связности в подмножестве типов местообитаний антропогенного происхождения, в то время как степень связности в подмножестве типов местообитаний с растительностью, близкой к естественной, значительно ниже. Наконец, ряд типов местообитаний с естественной растительностью флористически вообще являются «изолятами». Наличие повышенного количества изолятов отчасти можно объяснить тем, что в ландшафте классы местообитаний, представленные «изолятами», редки и сохранились лишь незначительными островами, так что флористические комплексы этих классов в данном ландшафтном районе далеко неполные. В нетронутом ландшафте с естественным растительным покровом в пределах этих классов, вероятно, также существовало большое флористическое и эколого-ценотическое разнообразие. Возможно, растительный покров полностью уничтоженных типов местообитаний данного класса флористически как раз и мог быть связующим звеном между теперешними «изолятами» и выделенными многокомпонентными классами местообитаний. Следовательно, сильное антропогенное изменение растительного покрова ландшафтного района приводит к распаду относительно непрерывной флористической структуры и появлению большого числа местообитаний, являющихся флористическими изолятами. Ландшафтный район дельты Немана может служить примером глубокой антропогенной модификации экологической структуры не только растительного покрова, но и других компонент биоты. На этом примере можно убедиться в том, что при полном уничтожении естественной растительности и замене ее растительностью антропогенной с низким уровнем ценотической организованности происходит снижение экологического разнообразия почти вдвое. Оставшиеся островки «дикой» природы в таком ландшафте оказываются флористическими изолятами, при этом снижается их способность оптимально перестраиваться при изменении климата, режима грунтовых вод при мелиорации и т. д. Подмножество типов местообитаний антропогенного происхождения имеет очень большую степень связности даже при высоком пороге флористического сходства. Эти местообитания не способны поддерживать флористический потенциал ландшафтного района.
 |
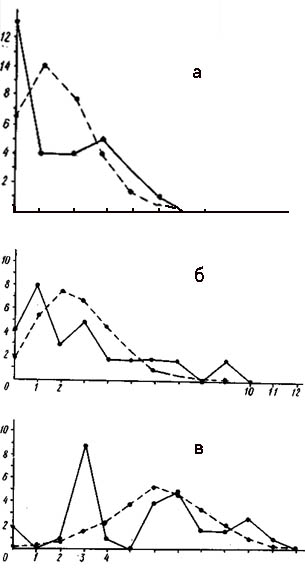
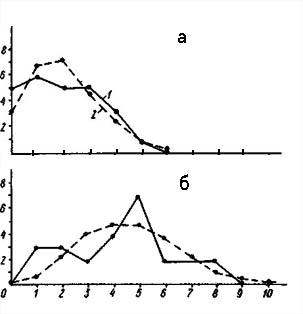
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района Прегольской низменности (а - уровень сходства более 30%, б - более 50%).
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
На рис. слева изображены кривые распределения вершин графа флористического сходства типов местообитаний района Прегольской низменности. При уровне сходства больше 50% граф имеет 27 вершин и 12 ребер при среднем количестве ребер на одну вершину 0,9. Распределение явно дискретно, так как «изолятов» значительно больше ожидаемого. Структура графа достоверно неравномерна (х2= 19,30), и множество вершин распадается на сверхбольшое количество изолятов, две группы, или связных компоненты, и одну «гантелю». В группы объединились типы местообитаний с антропогенным растительным покровом, сильно модифицированные человеком. |
При снижении порога сходства до 30% число ребер увеличилось до 48 при среднем числе ребер на одну вершину 3,6. Структура по-прежнему дискретна («изолятов» больше ожидаемого) и еще более неравномерна (х2= 54,99). При снижении порога сходства неравномерность структуры графа так же, как и в дельте Немана, усилилась. Выделилось подмножество типов местообитаний с относительно коренной растительностью. Это подмножество имеет сетчатый тип распределения ребер по вершинам, что соответствует высокой флористической дифференциации естественного растительного покрова при типологической континуальности его структуры. Подмножество с типичной антропогенной растительностью является типичной плеядой, или высокосвязной компонентой. В районе Прегольской низменности, так же, как и в дельте Немана, имеет место распад экологической структуры растительного покрова. Однако здесь этот процесс выражен несколько в меньшей степени.
 |
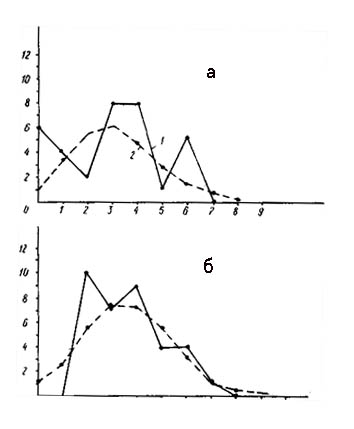
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района Вармийской конечно-моренной гряды (а - уровень сходства белее 50%, б - более 30%).
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
На рис. слева представлены кривые распределения вершин графа флористической структуры типов местообитаний района Вармийской моренной гряды. При пороге флористического сходства 50% граф с 25 вершинами имеет 24 ребра при среднем числе ребер на одну вершину 1,9. Структура графа дискретна, так как «изолятов» на графе больше ожидаемого, а вершин с двумя исходящими ребрами чуть меньше ожидаемого. Пространственный процесс, который моделирует данный граф, равномерный (х2 =2,46). |
При уровне сходства более 30% число ребер увеличивается до 54 при среднем числе ребер на вершину 4,3. «Изолятов» на графе нет, но и вероятность их существования очень мала. О неравномерности структуры говорят повышенное число вершин с одним и двумя исходящими ребрами и сниженное число вершин с тремя исходящими ребрами. В структуре графа четко выражены элементы типа плеяд. Степень насыщенности плеяд ребрами разная, и пространственный процесс довольно неравномерный (х2 =12,00). Высокосвязные компоненты графа соответствуют местообитаниям с антропогенной растительностью. Так же, как и в предыдущих районах, по мере снижения порога флористического сходства неравномерность структуры увеличивается. Нетрудно заметить, что в антропогенных ландшафтных районах глубина неравномерности флористической структуры несколько больше, чем в районах с естественной растительностью.
Сравнивая структуру всех трех аграрных районов между собой, можно заметить, что по степени антропогенной модификации экологической структуры они образуют ряд от крайне антропогенизированного ландшафта польдеров дельты Немана через район Прегольской низменности до ландшафта Вармийской моренной гряды, в котором заметную роль играют островки хвойно-широколиственных и широколиственных лесов и лугов с выработанной ценотической структурой.
 |
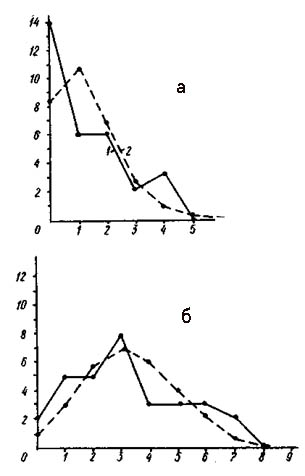
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района хребта Куркуре на Алтае (а - уровень сходства более 40%, б - более 30%).
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
На рис. слева изображены кривые распределения вершин графа флористического сходства типов местообитаний района хребта Куркуре при двух уровнях сходства. При уровне сходства 40% на 34 вершины графа приходится 54 ребра, среднее число ребер на одну вершину 3,2. Пространственный процесс явно дискретен, на графе повышенное число «изолятов» и пониженное число вершин с двумя исходящими ребрами. Неравномерность графа значительно выше ожидаемой. Здесь иного вершин с тремя и четырьмя исходящими ребрами, а также вершин с шестью ребрами, хотя вершин с пятью ребрами меньше ожидаемого. Хорошо заметно, что флористическая связность типов местообитаний альпийского и субнивального поясов значительно выше, чем лесного и субальпийского поясов. Только при уровне сходства более 50% компонента графа, соответствующая верхним поясам, распадается несколько звеньев. |
Снижение порога сходства до 30% увеличивает число ребер до 67, при среднем числе ребер на одну вершину - 3,8. Граф из дискретного и сильно неравномерного превратился в связный слабо неравномерный. При этом величина критерия Пирсона снизилась с 42,71 до 8,43. Исчезли «изоляты», стало меньше ожидаемого вершин с одним ребром (концевые вершины просто исчезли тоже), но столь же резко увеличилось число вершин с двумя исходящими ребрами. Одним словом, снижение порога флористического сходства привело к тому, что плеяды и изоляты оказались связаны между собой. Все добавочные вязи пошли на устранение дискретности и неравномерности исходного графа при пороге 30%. Глубина неравномерности Структуры графа хребта Куркуре невысокая, как и в районах Западной Чукотки. Это подтверждает предположение о большей глубине неравномерности структуры растительного покрова аграрных районов.
 |
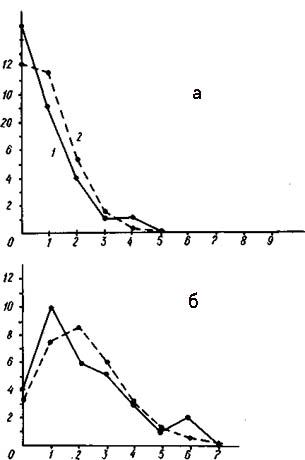
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района окрестностей г. Сыктывкара (а - уровень сходства более 30%, б - более 20%). При расчете сходства учитывались все виды.
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
Распределение вершин графа флористической структуры растительного покрова окрестностей г. Сыктывкара приведено на рис. слева. Напомним, что этот граф был получен путем сравнения не типов местообитаний, а конкретных геоботанических описаний. При уровне сходства более 30% в этом графе на одну вершину в среднем приходится 18 ребер. Структура дискретна, так как изолятов больше ожидаемого. Пространственный процесс в целом несколько неравномерный (х2=11,02). Вершин с четырьмя ребрами больше ожидаемого. При снижении уровня сходства до 20% число ребер возрастает до 50 (при среднем числе ребер на вершину 3,2). Дискретность-графа в этом случае заметно снизилась, а неравномерность структуры несколько увеличилась, хотя и весьма незначительно (х2= 13,40). На этом примере видно, что глубина дискретности и неравномерности структуры растительного покрова в этом районе меньше, чем в районах Калининградской области, но значительно больше, чем в районах с почти ненарушенным растительным покровом. |
Ординация конкретных местообитаний, а не их типов, привела к снижению связности графа, поэтому при анализе использован значительно меньший порог флористического сходства. Обращает на себя внимание, что на графе района окрестностей Сыктывкара значительную роль играют кольца и треугольники, хотя имеются и плеяды. По всей вероятности, это связано с увеличением разрешающей способности метода ординации конкретных местообитаний (геоботанических описаний), а не их типов. Есть основание полагать, что каждый цикл в структуре графа отражает некоторый экологический фактор или градиент этого фактора, вдоль которого и выстраивается цикл конкретных фитоценозов. В связи с сопряженностью данного экологического фактора с другими крайние значения напряженности этого фактора сильно деформируются другими факторами и оказывают сходное влияние на растительность. В этом случае экологический пространственный процесс замыкается. Вообще же проблема пространственных циклов экологических режимов в ландшафте заслуживает самого пристального внимания экологов, почвоведов и физиологов.
Обращает на себя внимание еще одна деталь графиков распределения вершин в зависимости от числа исходящих из них ребер. В ряде случаев эти графики становятся многовершинными, при этом каждая вершина на таком графике соответствует уровню связности определенного типа растительности. Так, антропогенная и рудеральная растительность с невыработанной ценотической структурой имеет самую высокую связность флористической структуры. Напротив, растительность коренных лесов и лугов с выработанной ценотической структурой имеет значительно меньшую флористическую связность - она флористически более дифференцирована.
 |
Распределение числа вершин в зависимости от их степени (числа исходящих из них ребер) графов флористического сходства типов местообитаний района окрестностей г. Сыктывкара (а - уровень сходства более 50%, б - более 30%). При расчете сходства учитывались только обильные виды.
По оси абсцисс - степень вершины, по оси ординат - число вершин с данной степенью; 1 - распределение эмпирическое, 2 - распределение, рассчитанное теоретически при условии, что ребра по вершинам распределяются случайно.
Анализ структуры растительного покрова района окрестностей Сыктывкара, выявленный по присутствию - отсутствию в конкретных местообитаниях только обильных видов (рис.слева) показывает, что данный пространственный процесс слабо дискретен и почти равномерен, во всяком случае более равномерен, чем при расчете сходства-различия по всем видам как обильным, так и не обильным. Величина критерия Пирсона при снижении порога сходства увеличилась, но незначительно (с 3,50 до 4,64). Это показывает, что разрешающая способность метода сходства только по обильным видам ниже, чем метода полного флористического сходства. Структура, выявленная по набору видов доминант, оказалась более континуальной и равномерной.
|
Проведенный анализ структуры графов флористического сходства местообитаний и типов местообитаний ряда ландшафтных районов показал, что метод распределения вершин графов в зависимости от их степени или числа исходящих из них ребер позволяет оценить степень дискретности-континуальности выявляемых структур, степень и глубину неравномерности распределения ребер по вершинам. При этом структура графа анализируется как целое и сравнивается со своей ожидаемой стохастической моделью, что позволяет использовать большой набор методов математической статистики и теории вероятностей, в частности позволяет использовать критерии достоверности.
Сравнение графов флористического сходства типов местообитаний районов с разной степенью нарушенности растительного покрова показало, что при сильном антропогенном преобразовании происходит не только распад ценотической структуры растительного покрова, но и распад структуры ландшафтно-экологической, резко увеличивается ее неравномерность и глубина неравномерности, увеличивается число местообитаний флористических изолятов и полуизолятов, которые, как правило, оказываются островками «дикой природы». Ценотически несформированный растительный покров (антропогенная растительность) имеет меньшее экологическое и, как следствие, меньшее флористическое разнообразие. Снижение экологического и эколого-ценотического разнообразия ландшафтного района и приводит в конечном счете к резкому уменьшению флористического потенциала.
На основе анализа литературных и наших данных мы пришли к выводу о том, что экологические условия местообитания неотделимы от экосистемы и во многом определяются биоценозом и фитоценозом. Как только уничтожается или сильно изменяется растительность, так сразу же на данном участке изменяется и экологический режим, т. е. возникает новое местообитание. В связи с этим сукцессионный процесс следует рассматривать как одновременное изменение и растительности и местообитания. Индикатором характера этих изменений может служить изменение набора видов конкретного контурфитоценоза или микрокомбинации растительности.
Наши данные по району окрестностей Сыктывкара говорят о том, что спорадичность в распределении доминантных видов в ландшафте больше, чем видов малообильных, сопутствующих доминантам. В связи с этим ординация элементов растительного покрова с использованием всего набора видов более экологична, чем ординация с использованием только видов доминантов и эдификаторов. Такая ординация, на наш взгляд, более корректна и позволяет вскрывать более тонкие моменты экологической и ценотической структуры растительного покрова ландшафтного района. Использование в ординации не типов местообитаний или групп ассоциаций, а конкретных описаний контурфитоценозов позволяет детальнее анализировать особенности ландшафтно-экологической структуры растительного покрова.
Одним из перспективных направлений в ландшафтной экологии и геоботанике нам представляется разработка методов анализа континуальных структур с помощью графов флористического сходства. Континуум не безлик, он не означает отсутствия структуры явления. Разные типы связи между элементами и разные структуры подпадают под категорию континуума. Это могут быть цепочки, треугольники, кольца, разнообразные их комбинации, сетчатые структуры и т. д. Представляются весьма удобными трактовка и анализ экологических, флористических и фаунистических структур биотического покрова ландшафта как особого рода пространственных эргодических процессов. Континуальность ландшафтно-экологической структуры растительного покрова высокая. Высокая флористическая связность растительного покрова ландшафта оказывается более свойственной хорошо сложившемуся растительному покрову с выработанной ценотической структурой. Состояние континуума, видимо, наиболее оптимальное экологическое и флористическое состояние растительного покрова. В таком состоянии растительный покров более мобилен, может быстрее перестраиваться при изменении климата, так как во флоре этого района всегда есть виды, которые в новых условиях окажутся адекватными местообитанию. Вместе с тем при изменении условий в контурфитоценозах, несмотря на то, что виды с узкой экологической амплитудой выпадают, в сообществе остаются виды с широкой экологической амплитудой - виды более толерантные, которые и «сшивают» растительный покров ландшафта в континуум - в непрерывную ткань. Эколого-ценотическая индивидуальность видов также очень важное свойство, обеспечивающее флористическую непрерывность растительного покрова и возможность его быстрых и тонких внутриландшафтных перестроек при изменении климатических и эдафических условий.
На основании выполненных исследований можно сформулировать некоторые общие выводы:
1. Графы попарного флористического сходства групп ассоциаций или типов местообитаний являются моделями флористической структуры растительного покрова, отражающими экологическую структуру не только растительного покрова, но и ландшафта в целом.
2. Графовые модели флористической структуры растительного покрова можно использовать при осуществлении ландшафтно-экологического ботанического мониторинга. В этом случае пробные площади должны быть постоянными и хорошо отмаркированными на местности. Периодическое переописание растительности этих площадей раз в 15-20 лет позволит получить временной ряд графовых моделей структуры растительного покрова, который и составит основу мониторинга.
3. Анализ степени равномерности структуры графов показал, что понятиями «континуальность» и «дискретность» свойства растительного покрова не исчерпываются. Дискретность растительности должна пониматься как крайняя степень неравномерности ее структуры, а континуальность, напротив, как крайняя равномерность распределения флористического сходства между, единицами растительности.
4. При оценке неравномерности флористической структуры растительного покрова целесообразно находить и учитывать уровень флористического сходства, при котором неравномерность имеет максимальное значение. Наши данные позволяют утверждать, что наиболее оптимальными для анализа являются графы, в которых на одну вершину в среднем приходится не менее трех ребер.
Читать следующий раздел

