Введение
А.В. Галанин © 2012
Проблема ботанического районирования, не смотря на огромное количество исследований в этом направлении, остается до сих пор недостаточно решенной. Об этом свидетельствуют жаркие споры, которые то и дело возникают между специалистами геоботаниками, флористами, лесоведами, экологами. Кроме того, по мере накопления новых данных о распространении видов и сообществ растений, часто приходится пересматривать положение и ранг границ между ботаническими территориальными подразделениями. Если в начале ХХ века ботаники много внимания уделяли концептуальным аспектам этой проблемы, то в конце века интерес к общим вопросам ботанической географии явно угас. Биогеографические концепции, которые активно дискутировались в начале века, к его завершению перешли в разряд «неприкасаемых», их нельзя подвергать сомнению и изменять. «Разрешено» только составлять карты растительности и картосхемы геоботанического районирования на основе этих ботанико-географических аксиом. Всякое ботанико-географическое инакомыслие не допускалось на страницы научных журналов, не говоря об учебных пособиях и учебниках. Одной из таких «неприкасаемых священных коров» ботанической географии, например, стало учение о широтной зональности растительного покрова.
О том, что поверхность Земли и все на ней находящееся, изменяется при продвижении с юга на север, было известно еще в античные времена. А.А. Григорьев (1929) считал, что зональный принцип использовался при физико-географическом районировании России уже в начале XIX столетия, так в географических описаниях России А.Ф. Бишинга, С.И. Плещеева и Е.Ф. Зябловского выделялись природо-хозяйственные зоны. Основоположником учения о природной зональности был русский геолог и почвовед, создатель почвоведения В.В. Докучаев (1899). Приведем почвенные зоны по В.В. Докучаеву: бореальная (тундровая); таёжная (лесная); чернозёмная (преимущественно степная); "аэральная" - зона безводных, субтропических стран (с лёссовой, барханной, каменистой и солонцовой областями); краснозёмная (латеритная) зона тропических стран . В «Краткой географической энциклопедии» (1962) сказано , что дифференциация географической (ландшафтной) оболочки Земли, проявляющаяся в последовательной и определённой смене географических поясов и зон, обусловлена, в первую очередь, изменениями количества лучистой энергии Солнца, падающей на поверхность Земли, в зависимости от географической широты. Там же сказано, что зональность присуща большинству компонентов и процессов природных территориальных комплексов: климатическим, гидрологическим, геохимическим и геоморфологическим процессам, почвенному и растительному покрову и животному миру.
Вот что по этому поводу пишет Е.Н. Лукашова (статья в БСЭ): "Географическая, закономерность дифференциации географической (ландшафтной) оболочки Земли, проявляющаяся в последовательной и определённой смене географических поясов и зон, обусловленной, в первую очередь, изменениями количества лучистой энергии Солнца, падающей на поверхность Земли, в зависимости от географической широты. Зональность присуща и большинству компонентов и процессов природных территориальных комплексов — климатическим, гидрологическим, геохимическим и геоморфологических процессам, почвенному и растительному покрову и животному миру, отчасти образованию осадочных пород. Уменьшение угла падения солнечных лучей от экватора к полюсам вызывает выделение широтных радиационных поясов — жаркого, двух умеренных и двух холодных. Формирование аналогичных тепловых и тем более климатических и географических поясов связано уже со свойствами и циркуляцией атмосферы, на которые большое влияние оказывает распределение суши и океанов (причины последнего — азональны). Дифференциация собственно природных зон на суше зависит от соотношения тепла и влаги, изменяющегося не только по широте, но и от побережий в глубь материков (закономерность секторности), поэтому можно говорить о горизонтальной зональности, частным проявлением которой является широтная зональность, хорошо выраженная на равнинах СССР. Каждому географическому поясу и сектору присущ свой набор (спектр) зон и их последовательность. Зональность проявляется и в закономерной смене высотных зон, или поясов, в горах, что также первоначально обусловлено азональным фактором — рельефом, однако и определенные спектры высотных зон свойственны определенных поясам и секторам. Зональность как горизонтальная, так и глубинная выражена и на океанах (последовательность изменения температуры, солёности, биокомпонентов и т.п.), на которые распространяются географические пояса. Зональность тепловых условий была известна ещё в античное время; для климата и растительности (в т. ч. проявление высотной З.) её установил немецкий учёный А. Гумбольдт (1962). Впервые в качестве основного закона дифференциации крупных природных комплексов зональность определена в 1898 В. В. Докучаевым".
Последователями учения В.В. Докучаева о природной зональности были: А.И. Воейков, Н.М. Симбирцев, Г.Н. Высоцкий, И.М. Крашенинников, К.Д. Глинка, А.Н. Краснов, Г.И. Танфильев, Л.С. Берг и А.А. Григорьев Э. Дригальский, О. Норденшельд, К. Тролль, С.В. Калесник, А.Г. Исаченко, Ф.Н. Мильков и др. (Мильков, 1969 ). Л.С. Берг (1947), А.А. Григорьев (1956, 1957, 1959, 1966), С.В. Калесник (1970) и А.Г. Исаченко (1971), исследуя причины и факторы географической зональности, пришли к заключению, что в формировании зональности наряду с величиной годового радиационного баланса и количества годовых осадков громадную роль играет их соотношение, степень их соразмерности. Отдельные аспекты учения о природной зональности служили предметом изучения в работах Г.Д. Рихтера, К.К Маркова, М.И. Будыко, А.М. Рябчикова, Е.Н. Лукашовой, Д.В. Богданова и др. (Будыко, 1971).
Между растительностью и климатом прослеживается очень тесная взаимосвязь такая, что широкое признание получило понятие биоклиматической зональности, в которой считается, что биотические зоны в точности совпадают с зонами климатическими (Григорьев, Будыко, 1956; Григорьев, 1957). Зоны растительности на картах обычно изображаются полосами, вытянутыми в субширотном направлении, и они не имеют резко выраженных границ. Постулируется, что каждой зоне растительности присущи типические особенности составляющих её компонентов растительных сообществ и процессов, восстанавливающих эти комплексы при нарушениях растительного покрова. Внутри зон, занимающих обширные полосы на поверхности Земли, различают более узкие подразделения - подзоны. Например, в пределах зоны саванн различают подзоны: влажных высокотравных, типичных сухих и опустыненных саванн, которые отличаются друг от друга ритмом сезонного развития всех растений, слагающих растительные сообщества этих подзон. Как правило, ботанические различия в подзонах саванн связанны с различиями в количестве и ритме поступления осадков. В зоне степей выделяют подзоны сухих и типичных степей, что также связано с осадками. В зоне лесов умеренного пояса выделяют подзону тайги (иногда её считают самостоятельной зоной), подзону смешанных лесов и подзону широколиственных неморальных лесов. Деление зоны лесов умеренного пояса на подзоны (часто их рассматривают в ранге самостоятельных зон) связано в основном с теплообеспеченностью территорий, а не с осадками, так как осадков в этих зонах (подзонах) выпадает достаточное количество. Таким образом, следует говорить как минимум о двух основных факторах, дифференцирующих растительный покров на планетарном уровне, это – количество получаемого тепла и количество получаемых осадков.
В каждой зоне и в каждой подзоне растительности, в зависимости от количества осадков, выделяются крупные долготные сектора: приокеанический, континентальный и переходный между ними, которым свойственны свои типы растительности, свои демутационные циклы и своя высотная поясность. В зависимости от положения в том или ином секторе внутри зон и подзон могут быть выделены более мелкие таксономические единицы, например: западно-приокеаническая темнохвойная тайга в Европе, континентальная светлохвойная тайга в Восточной Сибири, западно-приокеаническая темнохвойная тайга в Северной Америке. Для каждой зоны и каждого сектора свойственно определенное соотношение тепла и влаги, поступающих на единицу поверхности Земли. Это соотношение может быть выражено количественно (Григорьев , Будыко, 1956). Например, при значениях радиационного индекса сухости Будыко менее единицы для местности характерно избыточное увлажнение, при такой величине индекса выделяют влажные сектора в разных зонах и подзонах: лесов, лесотундры и тундры. При значениях индекса более единицы для местности характерно недостаточное увлажнение, в этом случае в пределах зон и подзон выделяют сухие сектора: степи, полупустыни и пустыни, а при значениях индекса близких к единице имеет место оптимальное увлажнение местности, в этом случае в пределах зон и подзон выделяют сектора: лесостепи, лиственных и светлых лесов и сухих саванн. Так, устанавливая "периодический закон зональности", А.А. Григорьев отмечал периодическое повторение одинаковых значений радиационного индекса сухости в разных зонах. Например, индекс сухости может быть равен единице и в зоне тундры, и в субтропической зоне, и в экваториальной зоне. Однако при этом годовой радиационный баланс, и годовая сумма осадков в этих зонах будут резко различными. Следовательно, нельзя при характеристике климатических условий зон использовать только один индекс сухости, необходимо указывать количество получаемого тепла и количество выпадающих осадков. Реальная влажность почвы, от которой зависит рост и развитие растений, определяется разницей между количеством осадков выпавших на земную поверхность и количеством осадков испарившихся с этой поверхности.
Следует обратить внимание на то, что при определении положения зон растительности и проведении их границ, по сути, учитываются два основных фактора, - это количество тепла, получаемое от Солнца поверхностью в виде излучения, а также количество осадков, выпадающих на поверхность. Количество получаемого тепла зависит не только от интенсивности излучения, но также и от того, насколько эффективно поверхность земли поглощает излучение и переводит его энергию в тепло. Температура поверхности, а, следовательно, и воздуха (воздух нагревается поверхностью земли) зависит от того, идет ли это тепло на нагревание и повышение температуры или расходуется на переход вещества из одного агрегатного состояния в другое, что, например, происходит при таянии льда или снега, когда тепловая энергия расходуется, а температура не повышается. Количество тепловой энергии, которая заключена в некотором теле, и величина его температуры – это далеко не одно и то же.
Осадки – это также фактор очень сложный, то они выпадают в виде снега, то в виде дождя, то их много зимой и весной и мало летом, то наоборот мало зимой и весной и много в конце лета и осенью. Рыхлые субстраты пропускают воду и в таких местах даже при сильном дожде избытка влаги не бывает. Напротив, если водоупорный горизонт залегает близко к поверхности, то избыточное увлажнение здесь бывает даже при небольшом количестве осадков, это мы наблюдаем в тундре на многолетней мерзлоте. Температура и влажность воздуха тоже влияют на судьбу воды, выпавшей на поверхность. При высокой температуре и малой относительной влажности воздуха выпавшая вода с поверхности и из верхнего горизонта почвы быстро испаряется и наблюдается ее дефицит в почве. В степи и пустыне нередко можно наблюдать, как капли дождя в жарком сухом воздухе испаряются так и не достигнув поверхности. Направление ветров и направление движения циклонов очень сильно влияют на количество выпадающих осадков. Если ветры дуют с моря на сушу, то они приносят на сушу влажный воздух и при его охлаждении в верхних слоях атмосферы пары воды конденсируются в капли дождя или снега, и на сушу выпадают обильные осадки. Если ветры постоянно дуют с суши в сторону моря, то осадков бывает очень мало, тогда пустыня может располагаться даже непосредственно на берегу моря, а не внутри континента.
Все сказанное выше хорошо известно, и показывает, насколько сложна гидротермическая система на Земле. Но именно эта система определяет пространственную дифференциацию биотического, и в первую очередь растительного покрова. Идеальная схема природных зон, учитывающая только годовые суммы температур и суммы осадков, оказывается настолько недостаточной в фитогеографии, что ее использование зачастую теряет всякий смысл. Но мы привыкли к несуразностям такого зонирования и не замечаем, что на одной и той же широте оказываются то тундры (южная оконечность Гренландии), а то широколиственные леса (окрестности города Осло), то дремучие темнохвойные леса (Вологодская область), то степи (окрестности Якутска). Но теория настолько довлеет над нами, что мы не придаем этим фактам существенного значения.
Ситуация становится еще более странной, когда мы пытаемся восстановить пространственную дифференциацию растительного покрова в иные климатические эпохи, например в эпоху последнего вюрмского оледенения, которое случилось 10-30 тыс. лет назад. В то время большое влияние на распределение тепла и влаги по поверхности планеты оказывали ледники в Скандинавии и в Канаде, а также в горах умеренного пояса. Скандинавский ледник, например, перехватывал большую часть влаги, которую приносили воздушные массы с Атлантики на восток. В связи с этим восточнее этого ледника климат в ту эпоху был сухой, жаркий летом и холодный зимой, этим он был похож на климат современной Якутии и Забайкалья. Холодные степи в то время имели большое распространение. Ледники на Чукотском полуострове, в Корякии и на Камчатке также усливали континентальность климата, но уже к западу от них - в Колымском нагорье и в Якутии. Степная растительность тогда и здесь имела большее распространение, степной пояс протягивался от Забайкалья до Чукотки и остров Врангеля, который в то время был частью осушенного шельфа Чукотского моря. Через Берингийский мост суши холодные степи Азии смыкались с холодными степями Северной Америки. Кстати, и сегодня прерии в Северной Америке имеют не широтное, а меридиональное простирание, что связано с экранирующим воздействием на влажные воздушные массы с Тихого океана цепей Кордильер.
Классическая теория природного зонирования говорит о том, что фактор тепла на характер растительности оказывает первостепенное значение, поэтому выделение растительных зон на Земле четко соответствует выделению термических климатических поясов: арктический, бореальный, умеренный, субтропический и тропический. Дифференциации растительного покрова в связи с разной степенью увлажнения обычно отводится второстепенная роль, так в пределах зон широтных растительности выделяются долготные сектора, связанные с разной степенью континентальности климата по мере удаления от океана. Эти подразделения называются не зонами, а областями, подобластями, провинциями, являющимися подразделениями зон.
Однако, если мы рассмотрим любую широтную зону растительности, то в ее пределах увидим столь сильные различия в плакорных растительных сообществах, что они, порой, более значительны, чем различия в плакорных сообществах соседних широтных зон. Например, в пределах зоны бореальных лесов мы имеем темнохвойную еловую тайгу в северной Европе и светлохвойную лиственичную тайгу в Восточной Сибири, которые между собой по типу биологического круговорота и видовому составу различаются больше, чем еловая темнохвойная тайга северной Европы и смешанные елово-широколиственные леса средней Европы, или лиственичная тайга Восточной Сибири и лесостепь Забайкалья.
Фактор «увлажнение» на характер растительности зачастую оказывает большее воздействие, чем фактор «тепло». Здесь не лишне вспомнить правило Либиха о том, что на растения оказывают большее воздействие факторы, находящиеся в минимуме. У каждого вида и каждого сообщества растений существует норма реакции, согласно которой живая система в абстрактном пространстве экологических факторов занимает определенную зону, и эта зона является зоной ее оптимума. Если какой-то из факторов находится за пределами толерантности системы (за пределами зоны оптимума), то никакие другие факторы в избытке не могут компенсировать фактор в минимуме. Правило Либиха было сформулировано для особей растений, но оно справедливо и для локальных популяций видов, а также для растительных сообществ.
Современные флора и р астительность большинства территорий формировались постепенно очень продолжительное время, и их подразделения имеют различный возраст. Некоторые виды и формации влажного тропического леса уже в миоцене существовали в тех же местах, где они произрастают сейчас, но флора и растительность тундровой и таёжной зон на месте их современного распространения имеют только четвертичный возраст. Как правило, родам растений и растительным формациям присущ более древний возраст, чем входящим в них видам и ассоциациям. В процессе развития в масштабах геологического времени соседние территории то конвергировали (сближались) по климатическим условиям, то диввергировали (расходились). При ковергировании климат их становился более сходным, в это время происходил обмен видами и флористическими комплексами (ассоциациями видов или растительными сообществами), флоры территорий становились более сходными вплоть до их полного объединения. При диввергировании климат становился все более различным, и флоры и растительности территорий развивались независимо друг от друга, их сходство на соседних территориях уменьшалось. На диввергировавшие территории приходили разные флористические комплексы, а популяции некоторых видов эволюировали в разных направлениях, становясь разными подвидами и видами.
Изменения климата во времени, как правило, цикличны, цикличны и изменения растительного покрова. Это приводит к тому, что массовые виды доминанты растительного покрова на двух соседних территориях в определенную эпоху могут быть разными (период климатической диввергенции), но флоры в целом этих территорий, учитывая виды малоактивные в ландшафте и реликтовые, будут весьма сходными, так как растительный покров «помнит» эпоху климатической конвергенции, а растительность ее довольно быстро «забывает». Например, современная растительность Даурии, если судить по доминирующим видам, существенно отличается от таковой Маньчжурии, но если судить по всем видам, то флоры Даурии и Маньчжурии весьма сходны – они принадлежат одной Восточноазиатской области. Это было показано нами при сравнительном анализе ряда семейств флоры Даурии и Маньчжурии (Галанин, 2006, Галанин и др., 2009).
Пространственные особенности растительного покрова (флоры и растительности) четко выявляются при геоботаническом и флористическом районированиях. Следует сказать, что полного соответствия между ними нет, так как главным критерием выделения подразделений растительности являются не таксоны, а жизненные формы растений, а при выделения подразделений флоры главными критериями являются общность таксономического состава (видов, родов, семейств). Ареалам типов р астительности соответствуют геоботанические области, а флористическим подразделениям – флористические царства, подцарства и флористические области. Например, на Западно-Сибирской равнине при продвижении с севера на юг прослеживается следующая смена типов р астительности и классов формаций: арктические тундры, субарктические тундры, тундровые редколесья, северная лиственно-пихтовая тайга, средняя кедрово-болотная тайга, южная елово-кедрово-пихтовая тайга, подтаёжные берёзово-осиновые леса, луговые степи, разнотравно-дерновинные степи, сухие типчаково-ковыльные степи.
Геоботанические области обычно делят на провинции, а последние - на геоботанические округа и районы. Сколь либо четких критериев отнесения хорологического подразделения растительного покрова к округу или району не существует. На практике разные авторы используют при ботаническом районировании территории разные критерии. Зачастую об этих критериях ничего вразумительного не сообщается, отчего схемы районирования, полученные разными авторами, получаются несопоставимыми.
На наш взгляд особое место в системе природных растительных зон занимает зона степной растительности. Для степной зоны умеренных поясов характерны: сухой континентальный климат; нередки засухи; количество осадков здесь - 200- 450 мм в год; поверхностный сток незначителен; почвы главным образом - черноземные, а в сухих районах - темно-каштановые и каштановые; растительность – сообщества многолетних злаков и разнотравья. Но существует еще одна степная зона в поясе субтропического климата. Степная субтропическая зона расположена небольшими участками между 25 и 40°. сев. и юж. широты в Северной и Южной Америке, на западе Азии, на юге и юго-западе Австралии. Для этой зоны характерны: климат теплый и сухой, лето жаркое, зима мягкая; осадков - 500- 600 мм в год; почвы - серо-коричневые, иногда засоленные; в составе растительности преобладают дерновинные злаки и кустарники, по долинам рек - галерейные леса. Субтропические степи в Южной Америке называются пампой, в Африке – саванной, в Северной Америке – прерией. До сих пор ботаники четко не определились, степная зона на Земле одна, или их несколько. Если выделять степную зону по фактору «тепла», то их на Земле несколько, а если по фактору «увлажнение», то одна. Но если за критерий принять свойства самой растительности (продуктивность, набор жизненных форм, видовой состав, ритм сезонного развития), то вообще никаких зон степной растительности на Земле выделять нельзя, а можно выделять различные аридные геоботанические области, подобласти и провинции.
Главными характерными особенностями типа растительности должны быть объем, скорость и степень замкнутости биологического круговорота, обеспечиваемого растительными сообществами этого типа. Эти параметры биологического круговорота вещества в наибольшей степени зависят от характера жизненных форм растений, сообитающих друг с другом в конкретных растительных сообществах, или, иначе, – в контурфитоценозах (Галанин, 1989; 1991). Именно поэтому для каждого типа растительности, характеризуя его, геоботаник обязательно называет жизненную форму доминирующих растений. Луга – это сообщества травянистых многолетних мезофитов, чаще всего корневищных многолетников; степи – сообщества травянистых ксерофитов с доминированием плотнодерновинных злаков, короткокорневищных злаков, осок и луковичных эфемероидов; светлохвойные леса – сообщества хвойных листопадных деревьев (лиственницы); темнохвойные леса – сообщества хвойных зимнезеленых деревьев (ели, пихты, псевдотсуги и др.); листопадные широколиственные леса – сообщества широколиственных деревьев, сбрасывающих на зиму листву (дуб, липа, ясень, клен и др.). Каждый тип растительности имеет свою длинную историю, насчитывающую десятки, а иногда и сотни миллионов лет, в течение которой происходила коэволюция совместно обитающих видов растений и животных. Эта эволюция постоянно корректировалась определенными климатическими и экологическими условиями внешней среды, с которыми данный тип растительности находился в относительном биогеохимическом равновесии.
За многие миллионы лет в растительном покрове Земли выработались весьма устойчивые сочетания жизненных форм и видов живых организмов, обеспечивающие относительную биогеохимическую устойчивость ландшафтных экосистем, - для каждого типа элементарного ландшафта сформировался свой особый тип экосистем и свой особый тип растительности. В умеренной зоне, при дефиците воды в течение почти всего вегетационного периода и при относительно хорошем увлажнении весной за счет тающего снега и дождей, не редких в это время года, устойчивым типом растительности являются степи, которые буквально расцветают за счет эфемероидов весной, но летом сильно усыхают. В степях запас фитомассы над поверхностью почвы в 7-8 раз меньше, чем запас фитомассы в корнях, луковицах и корневищах под землей (Cочава, 1986). Отмирающая фитомасса в степи разлагается медленно, поэтому здесь всегда имеется избыток органического вещества и в почве (черноземы) и на ее поверхности (прошлогодняя ветошь).
В умеренной зоне, но при отсутствии устойчивого дефицита воды в течение вегетационного периода, там, где сильно выраженный аллювиальный процесс не позволяет расти деревьям и высоким кустарникам, устойчивым типом растительности являются луга. Для лугов характерно значительно меньшее преобладание подземной фитомассы над надземной, процессы гниения здесь идут значительно интенсивнее, чем в степях. Черноземы под лугами не формируются. Сходным в степях и на лугах является то, что практически вся надземная фитомасса, нарастающая за вегетационный сезон, осенью отмирает, многолетними являются только подземные органы, из которых на следующее лето отрастает новая надземная фитомасса. В отличие от степей, на лугах разница в надземной и подземной фитомассе значительно меньше: в подземной сфере сосредоточено примерно две трети общего запаса фитомассы.
Для типа растительности верховое болото характерно еще более медленное, чем в степях, разложение отмершей фитомассы. Она буквально консервируется здесь на многие десятки и сотни тысяч лет, накапливаясь в виде торфов. Характерной жизненной формой для этого типа растительности являются сфагновые мхи – природные консерванты мертвой органики. Верховые болота образуются в условиях постоянного избыточного увлажнения в течение всего вегетационного периода. Болотные экосистемы характеризуются разомкнутым биологическим круговоротом вещества, чем напоминают экосистемы океана. Есть основание считать, что это очень древний тип экосистем (Галанин, 1989, 1992). Эволюция биосферы, по крайней мере в мезозойскую и кайнозойскую эры, происходила в направлении формирования экосистем, работающих по замкнутому типу и постоянного ускорения оборота вещества в трофических цепях жизни.
Тип растительности темнохвойная тайга отличается тем, что здесь значительная фитомасса накапливается в стволах и корнях деревьев и на многие десятки лет выводится из ежегодного биологического круговорота. Хвоя здесь живет 3-4 (5) лет, что также приводит к существенному замедлению биологического круговорота вещества. По замедленности биологического круговорота темнохвойные леса ближе к степям, но по типу «складирования» органического вещества, исключенного из круговорота, они есть нечто совершенно иное. Склад исключенного из ежегодного круговорота вещества в этих экосистемах содержится не в почве, а в самих растениях, в основном в стволах деревьев. Темнохвойная тайга устойчива в условиях бореального климата с достаточным в течение всего вегетационного периода увлажнением. Светлохвойная тайга от темнохвойной отличается тем, что хвоя здесь отмирает ежегодно, поэтому биологический круговорот интенсивнее, чем в темнохвойной тайге. Интенсификации круговорота вещества в светлохвойной тайге способствует и то, что весной и в начале лета, пока хвоя на деревьях не отросла полностью, под осветленным пологом леса интенсивно развиваются кустарники и травянистые растения. Ежегодно отмирающая наземная фитомасса травянистых растений в светлохвойных лесах – это весьма существенный элемент ежегодного биологического круговорота вещества в экосистемах светлохвойной тайги. Светлохвойная тайга устойчива в условиях резко континентального бореального климата в районах, где зимой выпадает мало снега, почва сильно промерзает так, что нередко образуется многолетняя мерзлота, а весна и начало лета отличаются крайней сухостью.
В аридные фазы климатических циклов активизировался криофильно-степной комплекс видов и начиналась миграция из Центральной Азии в Якутию, на Чукотку и через Беринг ийский мост в Аляску.
В гумидные фазы активизировался другой вектор миграций – с востока на запад.
Восточно-Азиатская ботанико-географическая область вытянута вдоль гумидного вектора и обязана своим возникновением и существованием Тихоокеанскому муссону. А что за хорион вытянут вдоль аридного вектора ? Этого хориона в настоящее время нет, однако он существовал в криоаридные эпохи плейстоцена. Сегодня флористические комплексы и типы растительности того времени встречаются вдоль вектора, обозначенного на карте красной стрелой.

Гумидная и аридная ботанико-географические дуги на карте Севровосточной Азии.
Основные понятия и термины
Особи растений, составляющие растительный покров, неодинаковы, и неодинаковость их двоякова рода. С одной стороны они образуют генетические системы, называемые видами, в которых связаны между собой потоками генетической информации. С другой стороны, расселяясь в пределах ландшафта, особи вступают в сложные взаимоотношения как с самим ландшафтом (средой обитания), так и друг с другом. Главными при этих взаимоотношениях являются процессы биогеохимические и биогеофизические. Системы, возникающие в результате этого взаимодействия – ценоячейки, синузии, контурфитоценозы, микро- и мезокомбинации растительности.
Говоря о ботанико-географической неоднородности растительного покрова мы имеем в виду его неоднородность флористическую и эколого-фитоценотическую. Элементарным звеном сравнения при ботанико- географическом районировании выступает элементарный ландшафтный район или район конкретной флоры в смысле А.И. Толмачева (1974) и одновременно конкретной растительности в смысле А.В. Галанина (1991). В пределах этого района справедливо правило А.И. Толмачева о том, что в пределах одной конкретной флоры на сходных местообитаниях встречаются сходные наборы видов. Выделяя ботанико-географические районы, провинции и подобласти мы следовали этому правилу.
Деление растительного покрова на хорионы (территориальные выделы различного ранга) по флористическому принципу не может совпадать с его подразделением по физиономическому принципу, за исключением тех редких случаев, когда распределение дифференцирующих флористических и биоморфологических (физиономических) признаков скоррелировано положительно.
Многие геоботаники считают, что разнообразие видов в растительном покрове определяется только разнообразием природных условий, которые существуют в той или иной местности в данное время. Роль исторических факторов в современном распространении видов, особенно на ограниченной территории, по их мнению, невелика (Сушкин, 1925; Braun-Blanket, 1928; Миркин, 1985 и др.)
Существует и иная точка зрения, которой придерживались геоботаники, начиная с 1820 г. с работ А. Гумбольдта (Гумбольд, 1962). Эта точка зрения положена в основу классических классификаций растительности В.Н. Сукачева, А.П. Шенникова и др. Например, В.Б. Сочава (1944) подчеркивал искусственность деления географии растений на историческую и экологическую. Современное экологическое состояние любого хориона имеет свою историю; виды, проникшие в район в прежних условиях, в изменившихся климатических или орографических условиях не исчезают сразу. Они снижают свою активность, но сохраняются на немногих местообитаниях, где образуют ассоциации с другими видами, проникающими в данный хорион в связи с изменением климата.
Подтверждением этой точки зрения можгут служить результаты интродукционных исследований, которые показали, что многие виды растений не растут в той или иной местности вовсе не потому, что условия этой местности для них не подходят. Просто исторически сложилось так, что они (эти виды) не могли мигрировать в эту местность (Беликович, Галанин, 2005).
Следует различать разные масштабы пространственно-временных изменений растительного покрова, при этом можно выделить следующие ступени (уровни):
1) Эколого-ценотическая ступень (уровень) имеет дело с пространственными размерами порядка 10 -100 0000 кв. м и временными интервами 10-100 лет. В пределах этой ступени характер и динамика расселения растений определяется не столько характером экотопа (поскольку он весьма однороден), сколько взаимодействием растений с окружающей средой и друг другом. В результате этого взаимодействия происходит трансформация среды обитания растений, в известной степени определяемая генотипом растений. Экотоп трансформируется в биотоп.
2) Ландшафтно-экологическая (вековая) ступень (уровень) имеет дело c пространственными размерами порядка 1-1000 кв. км и временными интервами порядка 100-1000 лет. В пределах этой ступени характер размещения и динамика расселения видов по местообитаниям в основном определяется ландшафтно-экологической неоднородностью экотопов и флорой, - точнее флорпистическим потенциалом растительного покрова или структурой семенного дождя – «дождя диазачатков растений».
Понятие "Конкретная флора" А.И. Толмачева, 1932) связано именно с этим уровнем пространственно-временной организации растительного покрова.
3) Эколого-географическая ступень (уровень) имеет дело с пространственными размерами порядка 1000 -10000 кв. км и временными интервалами порядка 1000-10000 лет; в пределах этой ступени характер размещения и динамика расселения видов в основном определяются климатическими изменениями и колебаниями уровня моря, а также миграциями видов;
4) Микроэволюционная ступень (уровень) имеет дело c пространственными размерами 10000-100000 кв. км и временными интервалами порядка 10000-1000000 лет («время микроэволюции»); в пределах этой ступени динамика растительного покрова определяется крупномасштабными изменениями лика Земли, в том числе тектоникой, крупномасштабными перестройками климата, вымиранием видов, миграцией видов и флор с соседних территорий и образованием новых подвидов и видов - микроэволюцией.
5) Макроэволюционная ступень (уровень) имеет дело с пространственными размерами более 100000 кв. км и временными интервалами 1 000 000 - 100 000 000 лет (время макроэволюции); в пределах этой ступени состояние и динамика растительного покрова определяются мегамасштабными изменениями литосферы, гидросферы и атмосферы, крупномасштабными вымираниями видов, родов и семейств, образованием новых родов и семейств растений.
Считаю, что изучать растительный покров в рамках геоботаники и биогеографии следует не столько как объект, сколько как процесс! В этом суть динамической парадигмы в географии растений и геоботанике. Только в рамках этой парадигмы можно выйти из тупика, в котором оказались эти науки в конце ХХ столетия.
Все живые системы от клетки до биосферы - явления существующие в четырехмерном пространстве-времени, следовательно и изучать их следует как четырехмерные явления.
Как всякая система, растительный покров обладает определенными имманентными свойствами, т.е. его пространственная неоднородность не может полностью совпадать с неоднородностью внешней среды. Каждое новое состояние растительного покрова несет в себе инерцию его прежних состояний. Этот шлейф последействия прослеживается в таксономической и ареалогической структуре флор на протяжении десятков, а иногда и сотен тысяч лет. Поэтому в растительном покрове имеют место реликтовые виды и реликтовые сообщества. Прежде они находились в равновесии с внешней средой региона до тех пор, пока окружающая их среда не изменилась.
К имманентным свойствам растительного покрова относится сверхразнообразие растительных сообществ. При циклических изменениях климата в сообществе в малом обилии накапливаются виды, которые в биологическом круговороте экосистемы в данное время никакой роли не играют, но которые становятся обильными при возвращении прежних климатических условий. Благодаря сверхразнообразию растительное сообщество может быстро перестраиваться, и экосистема постоянно находиться в динамическом равновесии с внешней средой в условиях циклических изменений климата.
Кроме того, благодаря циклическим изменениям климата, в растительном покрове существуют ценотические системы, которые я называю ценотическими маятниками. Ценотический маятник обычно образуют два вида, которые являются содоминантами (соэдификаторами) растительных сообществ, хотя требования этих видов к внешним условиям отличаются.
В одной фазе климатического цикла преобладает вид А, а в другой фазе - вид В. Циклы изменения климата при этом должны быть соизмеримы с длительностью жизни особей. Циклическая динамика климата не позволяет ни одному виду из ценотического маятника полностью вытеснить другой.
В Даурии, например, в верхнем лесном поясе, такой фитоценотический маятник образуют лиственница даурская (Larix dahurica) и кедр сибирский (Pinus sibirica), в Южном Приморье - пихта цельнолистная (Abies holophylla) и кедр корейский (Pinus korajensis), в среднем Сихотэ-Алине - ель аянская (Picea ajanensis) и кедр корейский (Pinus korajensis), в Северо-Восточной Европе - ель сибирская (Picea obovata) и сосна обыкновенная (Pinus sylvestris), в Якутии - сосна обыкновенная (Pinus sylvestris) и лиственница Каяндера (Larix cajanderi).
На Дальнем Востоке главным фактором, дифференцирующим растительный покров, несомненно, является Тихоокеанский муссон. Этот фактор наиболее сильно сказывается в приморских районах, но ослабевает по мере продвижения на запад. Западный предел действия Тихоокеанского муссона – Забайкалье. При циклических изменениях климата бывают периоды, когда западная граница действия этого муссона сдвигается на восток, а в другие периоды климатических циклов, наоборот, на запад. Но абсолютным пределом для Тихоокеанского муссона на западе является и являлся в прошлом меридиан озера Байкал, восточных макросклонов Восточного Саяна и Хангая. На юго-западе зона этого муссона ограничена пустыней Гоби, на севере - зоной, климат которой контролируется Северным Ледовитым океаном.
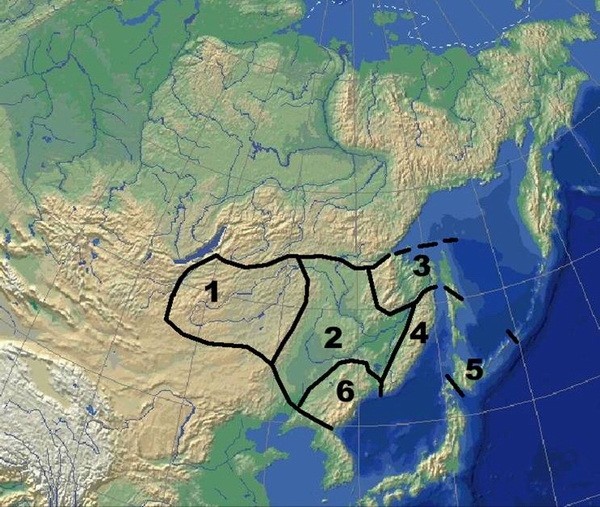
Подобласти Восточно-Азиатской ботанико-географической области бореально- неморально-аридной растительности:
1 – Даурская, или Даурия.
2 – Маньчжурская, или Маньчжурия.
3 – Нижнеамурско-Северосахалинская.
4 – Сихотэ-Алинская.
5 – Сахалино-Южнокурило-Хоккайдская.
6 – Хасанско-Корейская.
|
Выделяя умеренную неморально-бореально-аридную Восточно- Азиатскую ботанико-географическую область, мы проводим ее северную, западную и юго-западную границы по границе распространения Тихоокеанского муссона. Южная граница области примерно совпадает с изотермой среднеянварской темперартуры +1°С, южнее этого рубежа начинается зона субтропиков.
Отдельные подразделения области на уровне подобластей связаны между собой миграционными потоками видов и процессами видообразования.
Именно эта динамика делает Восточно-Азиатскую ботанико-географическую область объективной реальностью, существующей как в пространстве, так и во времени.
Например, граница между лесом и даурской степью-прерией в Южном Забайкалье постоянно сдвигается в зависимости от фазы климатического цикла. В настоящее время идет фаза аридизации климата, отчего активизируется степной компонент растительности, а лесной деградирует.
В периоды с влажным гумидным климатом даурская прерия сменяется лесом. Результаты мониторинга растительности говорят о ксерофитизации растительности Даурии в последнее столетие. По наблюдениям Никиты Петровича Соколова на гольце Сохондо, 250 лет назад климат там был более гумидным, чем сегодня, верхняя граница леса тогда проходила на 150 м выше, а нижняя - на 100 м ниже, чем в настоящее время.
Ритмичность изменения климата Земли, по крайней мере в плейстоцене, - научно доказанный факт. Об этом говорят чередования ледниковых эпох с межледниковьями, установленные палеогеографами.
|
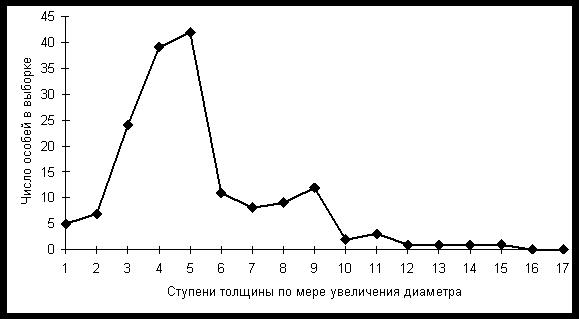
На графике показано частотное распределение особей тиса остроконечного (Taxus cuspidata) в Сихотэ-Алинском заповеднике в зависимости от толщины ствола на высоте 1,3 м от земли (Галанин и др., 2005б).
|
Нами установлено, что толщина ствола четко скоррелирована с возрастом дерева так, что популяция тиса гетерогенна по возрасту деревьев. Пополнение ее новыми особями, как следует из частотного распределения, происходило ритмично. Тис на территории заповедника возобновлялся в гумидные фазы цикла и не возобновляется в аридные.
Измерив годичные приросты тиса в заповеднике, мы убедились, что особи первого пика толщины имеют возраст 250-300 лет, второго – 800-900, самые толстые 1500-1700 лет и более (до 2000 лет ). Благодаря живучести особей и их долголетию, популяции тиса могут использоваться как инструмент для выявления климатических циклов в среднем и северном Сихотэ-Алине. В южной части Приморского края тис неплохо возобновляется и в современную эпоху, и его популяции здесь не могут служить таким инструментом.
О долговечности тиса остроконечного, например, говорит тот факт, что некоторые его особи были придавлены и пригнуты к земле. Однако даже слабого намека на остатки упавших кедров исполинов, пригнувших особи тиса нам обнаружить не удалось.
Похоже, что это частотное распределение отражает вековой 600-700 летний ритм колебаний климата. Такой промежуток времени отделяет два пика на диаграмме.
|
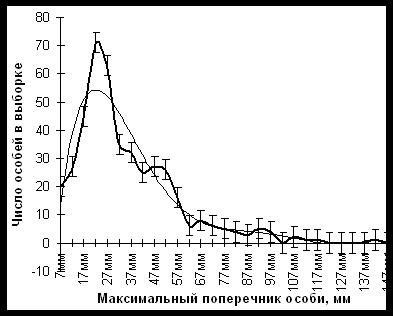
Частотное распределение особей накипного эпилитного лишайника (Rhizocarpon geographycum) в зависимости от их максимального поперечника на горе Сохондо (Южное Забайкалье).
|
На этом графике изображено частотное распределение особей накипного эпилитного лишайника (Rhizocarpon geographycum) в зависимости от их максимального поперечника в субальпийском поясе Сохондинского заповедника. Исследовалась популяция на глыбах каменной россыпи на стационаре Верхний Букукун. Видно, что популяция по размерам особей гетерогенна, а, следовательно, гетерогенна и по их возрасту. Это значит, что пополнение популяции новыми особями представляет собой ритмичный процесс, когда благоприятные для этого лишайника эпохи сменяются неблагоприятными.
Лихенологические исследования в субальпийском поясе Сохондинского заповедника в 2001 г. и исторические материалы позволили выявить размерную структуру популяций лишайников эпифитов, которая также отражает 600-700 летний климатический цикл, в течение которого гумидные эпохи сменялись аридными. Максимумы гумидности в прошлом имели место во II-III веках до новой эры, в V-VI , ХI -XII и XVII-XVIII веках новой эры. Напротив, в I-II , VIII-IX , XIV- XV и XX-XXI веках здесь происходили и происходят процессы аридизации климата. Этот вывод об аридизации климата в XX веке подтверждается и инструментальными наблюдениями на метеостанции "Букукун". Так, с 1949 г. среднегодовая температура здесь повысилась на 1,3°С, а годовое количество осадков снизилось на 20-25%.
|
Деградация пастбищ в аридные фазы климатического цикла заставляла народы Центральной Азии, сначала гуннов, потом тюрков, затем монголов теснить своих восточных, западных и северных соседей. Мощные миграционные волны кочевников из Центральной Азии повторялись примерно через каждые 600 лет.
В гумидные эпохи вектор миграций народов менялся с точностью до наоборот. В XVI-XVII веках (гумидная эпоха) на восток шли русские землепроходцы. В одну из прежних гумидных фаз с Запада на Восток мигрировали племена европеоидов динлинов. Вероятно, имели место западные волны расселения с запада на восток и в более ранние гумидные эпохи.
Кроме векового 600-700 летнего климатического ритма существует 40700-летний ритм, связанный с наклоном земной оси к плоскости земной орбиты (эклиптики). Земная ось раскачивается, и когда наклон оси к плоскости эклиптики уменьшается, в это время сокращаются теплый и холодный климатический пояса, но расширяются области умеренного климата. Похоже, что чередование ледниковых эпох на Земле в плейстоцене происходило именно с такой периодичностью (Галанин, 2001).
Другой ритм с периодом 92000 лет связан с наличием эксцентриситета (вытянутости в виде элипса) земной орбиты, который также циклически изменяется. При этом орбита Земли изменяется от круговой до эллиптической. Если эксцентриситет увеличивается, то летние месяцы в северном и южном полушариях сильно отличаются по количеству получаемой солнечной энергии от южных, если эксцентриситет уменьшается, то все происходит наоборот (Галанин, 2001).
В настоящую эпоху ось вращения Земли направлена на Полярную звезду. Но так было не всегда.
|

Земля вместе с Солнечной системой обращается по эллиптической орбите вокруг ядра Галактики с периодом в 180- 200 (210) млн. лет. За галактический год Солнечная система проходит разные зоны Галактики – рукава и межрукавные пространства, отличающиеся плотностью межзвездного газа и гравитацией. За галактический год происходит пять вспышек в ядре Галактики.
Примерно через каждые 31-32 млн. лет на Земле происходит резкое увеличение радиоактивности и, как следствие, вспышки видо- и родообразования в биосфере.
|
Цикличность во Вселенной порождает развитие материального мира: благодаря цикличности время становится вектором, оно имеет направление из прошлого в будущее. Если бы не было цикличности, не было бы развития, а время, как пространство, было бы не вектором, а скаляром - оно не имело бы направления, то есть понятия "прошлое" и "будущее" потеряли бы смысл, прошлого и будущего просто бы не существовало.

Тыринское озеро в Ононской Даурии на границе с Монголией. На гранитных скалах видны следы ветровой эрозии. Когда-то климат здесь был более аридный - пустыня Гоби в те эпохи приходила сюда. Фото: © А.В. Галанин.
|
Циклоны, зарождаясь в Тихом о кеане, несут влагу на материк.
Но по мере продвижения на материк их мощность ослабевает.
Тихоокеанский муссон действует на протяжении длительного геологического времени, поэтому хорологическое подразделение растительного покрова, обусловленное этим фактором, заслуживает высокого ранга ботанико-географической области.
Колебания климата в направлении "гумидность-аридность" вызывали миграции видов в сторону то материка (на запад), то в сторону океана (на восток). Изменения климата в направлении потепления-похолодания приводили к миграциям видов и целых сообществ растений с севера на юг и обратно. В соответствии с этими изменениями климата изменялся и вектор естественного отбора.
Проводниками северных видов на юг служили горные цепи, а южных на север - долины рек и южные склоны гор.
Если внимательно присмотреться к современным ландшафтам, то в них - в их компонентах - можно увидеть следы прошлых климатических эпох: реликтовую вечную мерзлоту в степях Даурии, реликтовые песчаные барханы на севере Читинской области и даже в Якутии. |
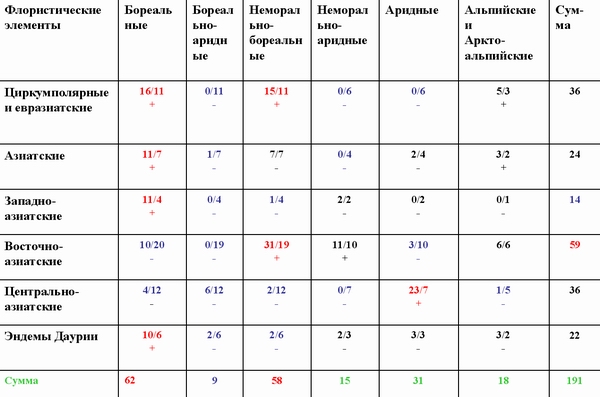
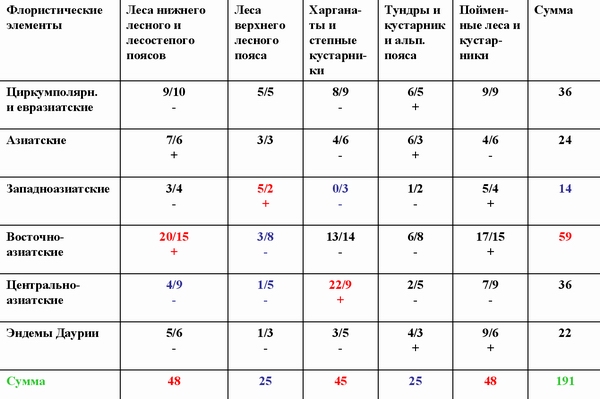
Сопряженный анализ структуры дендрофлоры Даурской подобласти Восточноазиатской ботанико-географической области. В числителе - наблюдаемое число видов в элементе пересечения, в знаменателе - рассчитанное теоретически при условии, что сопряженность между структурами отсутствует.
|
При климатических изменениях одни виды растений были более подвижны, они мигрировали быстрее (виды ив, тополей, лиственниц, берез, кленов) другие- медленнее (кедр сибирский, кедровый стланик, дуб монгольский, орех маньчжурский), что связано с особенностями распространения семян и плодов у этих видов.
Западная граница Восточно-Азиатской ботанико-географической области совпадает с довольно резким флористическим градиентом, хорошо выраженным в ареалогической структуре конкретных флор (Беликович, Галанин, 2005, Галанин и др.2005; Галанин, Беликович, 2006; Наумов, 2009).
Многие виды с восточноазиатским типом ареала выклиниваются на долготе Байкала, и, как было нами показано на примере дендрофлоры Даурии, структура флор Южной Сибири существенно отличается от таковой Даурии.
Сопряженный анализ распределения видов дендрофлоры Даурии по долготным и широтным ареалогическим элементам показал, что между долготной и широтной протяженностью географического ареала видов в дендрофлоре Даурии существует сопряженность. Положительно сопряжены: циркумполярные и евразиатские виды с бореальным и неморально-бореальными широтными поясами растительности. Азиатские и западноазиатские виды - с бореальным поясом. Восточноазиатские - с неморально-бореальным. Центральноазиатские виды - с аридным поясом растительности. А вот эндемы Даурии положительно сопряжены с бореальной растительностью. Выявленные положительные и отрицательные сопряженности говорят о направлениях миграций видов и условиях, в которых происходили эти миграция. В холодные влажные эпохи активизировались циркумполярные, Азиатские и западноазиатские виды (миграции с запада и с севера). В сухие теплые эпохи активизировались аридные центральноазиатские виды (миграции с юга и востока). В теплые влажные эпохи активизировались неморально-бореальные виды (миграции с востока).
Те же закономерности выявляются и при анализе сопряженности ареалогической структуры флоры с ландшафтно-экологической структурой. Западноазиатские виды положительно сопряжены с лесами верхнего лесного пояса, восточноазиатские - с лесами нижнего лесного пояса, а центральноазиатские - со степными сообществами и степными кустарниками - харганатами. Этот анализ показывает структуру миграционных потоков видов и их приуроченность к разным фазам климатического цикла (Галанин и др., 2005).
|
Мы не претендуем на точность проведения границ Восточно-Азиатской неморально-бореально-аридной ботанико-географической области и ее подобластей. Дальнейшие исследования несомненно внесут корректировку в их положение, особенно на территории Монголии, Китая и Кореи.
Разделение подобластей на провинции – это следующий этап ботанико-географического районирования. Пока что мы смогли разработать такое районирование только для Даурской подобласти, растительный покров которой исследовали в течение 25 лет.
Мы надеемся, что предлагаемая схема поможет более оптимально планировать ботанические исследования в регионе и проводить их в русле динамической и филогенетической парадигм.
Небесполезной эта схема будет и при долгосрочном планировании ресурсопользования и охраны природы в этом регионе особенно там, где это требует международного сотрудничества.
Использованная литература

