Самоорганизация древесных ценозов
Н. А. Василенко © 2012

Наталья Александровна Василенко
Кандидат биологических наук, участница и организатор многих ботанических экспедиций по Дальнему Востоку и Забайкалью, зав. лабораторией экспериментальной фитоценологии Ботанического сада-института ДВО РАН. Область научных интересов: экспериментальная фитоценология, лесоведение, пространственная структура древостоев, зеленое строительство и ландшафтная архитектура, экологическое образование.
botantehnol@yandex.ru |
"Прежде всего нет смысла говорить о противопоставлении часть - целое. С целым как таковым мы не имеем дело. Какой бы детальный архетип мы не приняли, он не будет состалять целого, но лишь какую-то его часть (аспект). В этом смысле мы не можем оперировать с целым, которое остается в области трансцендентного и доступно лишь созерцанию, интуитивному восприятию, откровению"
С. В. Мейн
Василенко Н.А. Самоорганизация древесных ценозов. Владивосток: Дальнаука, 2008. 171 с.
В книге анализируется формирование и развитие в процессе динамики древостоя различных ценотических групп деревьев, отличающихся по диаметру. Выявлены особенности пространственного размещения деревьев в фитоценозах, в фитогенных полях соседей деревьев. Использованы оригинальные методы, позволяющие уловить пространственный ритм внутри каждого древостоя. Книга написана на материалах, собранных на постоянных пробных площадях в разных типах лесов российского Дальнего Востока.
Рассчитана на геоботаников, экологов, работников заповедников и специалистов лесного хозяйства. Ил. 120, библ. 171.
Vasilenko N.A. Self-organization of forest coenoses. - Vladivostok: Dalnauka, 2008. - 171 p.
The monograhpy is devoted to the issue about various tree groups fprmation and development as well as features of spatial distribution of trees in plant communities. The book is based on original materials of long-term reseach on the constant plots lokated in different nature reservs and various forest types. The constant plots are distribed and treated by inform technique that makes possible to compare the received results the issue of the stand functional elements formation. The numerous data schow that in various stands division of coenotic population of dominants and edificators on discrete groups by diameter can be assumed as a process of self-organizing of plant community as a whole. One of the chapters of the books studies peculiarities of spatial tree distribution in the forest. It is shown that within every stand, specific spatiel rhytm is maintaining together with the formation of the inheren order of functional part.
The book is intendid for geobotanists, ecologists, employees of nature resrves and forestry experts. Ill. 120, bibl. 171, suppl. 1 |
Глава 3
ФОРМИРОВАНИЕ И РАЗВИТИЕ ЦЕНОТИЧЕСКИХ ГРУПП ДЕРЕВЬЕВ, ОБРАЗУЮЩИХ ДРЕВОСТОЙ
При понимании процесса самоорганизации наряду с исследованием упорядоченных структур, образующихся в открытых системах в результате этого процесса, важно изучение порядка возникновения этих структур и последовательности их развития во времени. Для того чтобы начался процесс самоорганизации, необходимо, чтобы отбор происходил при определенных условиях, а именно: система должна быть далекой от равновесного состояния; и интенсивность роста числа элементов должна быть достаточной для того, чтобы вывести эту систему из устойчивого состояния. В любой системе со временем назревает необходимость качественной смены (перестройки) самой структуры (Шустер, 1987; Лоскутов, 1990; Аршинов, Буданов, 1994). Краткий момент неустойчивости, балансирования системы на острие выбора между будущими состояниями, когда судьба всей системы может зависеть от вторжения одной случайной флуктуации, и есть ее точка бифуркации (Князева, Туробов, 2003).
Растительные сообщества являются динамическими системами. Это означает, что с момента возникновения взаимодействия между компонентами сообщества постоянно изменяются во времени. В этом разделе мы рассматриваем лесные фитоценозы в сомкнутом растительном покрове. Как известно (Шенников, 1964; Работнов, 1978), динамика таких сообществ проходит намного сложнее. Каждый фитоценоз в сукцессионном ряду рассматривается как стадия сукцессии и возникает он не сразу, а имеет определенную "историю" развития. Разные исследователи развития растительных сообществ в ходе эндогенных сукцессий предлагали различные типы сукцессионных рядов и указывали на то, что стадии экзогенной и эндогенной смен имеют соответственно характер смены доминирования не отдельных видов, а целых групп видов (Сукачев, 1942; Шенников, 1964; Ниценко, 1973; Миркин, 1984). И в фитоценологии, и в синергетике можно найти один и тот же тезис - это тезис о дискретности возможных состояний, в которые может переходить система в процессе динамики, а также заданность, ограниченность их числа.
Иначе говоря, спектр возможных структур-аттракторов динамики не является сплошным. В ходе сукцессионных смен растительных сообществ возможен только определенный набор путей, ибо только этот набор соответствует внутренним свойствам рассматриваемой системы. Набор аттракторов – это все растительные сообщества, которые могут образоваться в конкретном ландшафтно-экологическом районе благодаря наличию экологического потенциала в виде конкретной флоры (Юрцев, 1982, 1987; Галанин, 1989; Князева, Туробов, 2003).
Как известно, внутри процесса самоорганизации лежит взаимодействие между элементами системы. Результат этого влияния различен для каждого из участников взаимодействия. При взаимодействии естественным образом происходит выявление отличий объектов, то есть их сравнение, и на этом основании неслучаен их отбор: менее устойчивое, более податливое вытесняется, трансформируется, заменяется более устойчивым (Саночкин, 2001). Это касается и выбора функциональных элементов в процессе динамики систем.
Дифференциация деревьев по толщине тесным образом связана с эндо- и экзогенными сукцессиями лесной растительности. Важным вопросом при этом является выяснение закономерностей образования ценотических групп деревьев в процессе динамики. Может ли в первоначально однородном древостое или отдельной ценопопуляции образоваться определенная структура в виде ценотических групп деревьев? Как ведут себя отдельные группы деревьев в процессе динамики? И может ли хаотичное состояние отдельных групп деревьев распространиться на всю структуру древостоя или отдельных ценопопуляций деревьев? В литературе можно встретить некоторые сведения о закономерностях динамики рядов распределения деревьев в различных древостоях (Макаренко, 1967, 1974; Моисеев, 1971; Дыренков, 1973; Свалов Н., Свалов С., 1973; Лебков, 1973; Мошкалев, 1974; Ястребов, 1989).
А.Б. Ястребов (1989: 53) предложил следующую схему хода дифференциации древостоя. «Первоначально древостой однороден. Впоследствии в результате смыкания ризосфер и крон развивается конкуренция между деревьями, и ценопопуляция распадается на два ценотических класса. Меньшая часть становится господствующими деревьями, а большая - угнетенными I порядка. В дальнейшем, в результате периодического смыкания внутри класса господствующих деревьев от него отделяются группы угнетенных II, III и т.д. порядков. На все это накладывается процесс перехода части деревьев из более высоких ценотических групп в более низкие». Из-за гибели большого числа особей в группе угнетенных деревьев, эта группа более рано отделяется хиатусом от группы господствующих, и, в конце концов, все угнетенные особи погибают. Эти процессы дифференциации приводят к тому, что в древостое одновременно присутствуют, как правило, три ценотических группы деревьев. На эти процессы накладывается еще процесс стохастического отпада из всех ценотических групп.
Роль стохастического отпада усиливается со временем. К концу жизни древостоя разрежение в нем достигает такой степени, что оставшиеся деревья почти не взаимодействуют, и постепенно возникает относительная однородность древостоя. Такая схема хода дифференциации, предложенная А.Б. Ястребовым, носит обобщенный характер. Динамика упорядочивания во внутренней структуре древостоев - процесс сложный и многогранный, причем и для древостоев в целом, и для составляющих ценоз отдельных ценопопуляций. Общей закономерностью процесса дифференциации деревьев по толщине является тенденция к самоусложнению ценотической структуры во времени. В процессе самоорганизации древесных ценозов лидирующее и подчиненные положения в ходе сукцессионных смен имеют шансы занять именно те ценопопуляции, которые могут образовывать устойчивые функциональные элементы в виде групп деревьев различного диаметра.
Так, занимая промежуточное положение на пути к равновесной структуре системы, ценопопуляции раннесукцессионных видов деревьев, по-видимому, не способны к образованию устойчивой структуры в виде длительно существующих ценотических групп. Это можно увидеть на примере ценопопуляции березы плосколистной и осины Давида, произрастающих в березняке разнотравно-кустарниковом (рис. 31-32), и ценопопуляции березы плосколистной в березняке разнотравно-осоково-папоротниковом (рис. 33) на территории Сихотэ-Алинского биосферного заповедника. Эти вторичные сообщества образовались в результате пожаров на месте кедровников. Динамика процесса дифференциации в каждой рассмотренной ценопопуляции идет индивидуально, общей направленностью служит переход особей по мере роста в толщину в группу деревьев с большим диаметром ствола, при этом в отдельные периоды наблюдений однородность в ценопопуляциях березы и осины сменяется состоянием неоднородности по толщине.
Но ценотические группы в таких ценопопуляциях являются неустойчивыми структурными образованиями, они не могут существовать длительное время и постоянно перестраиваются. По-видимому, это характерный признак для серийных видов, которые занимают промежуточное положение в сукцессионных рядах. Для трех рассмотренных нами ценопопуляций деревьев характерной чертой является отсутствие притока особей из подроста и быстрое разрушение существующей на данный момент структуры за счет потери старых элементов. Это является общим принципом для систем любого уровня и природы: если скорость роста числа элементов невелика и не превышает скорость отмирания старых, то это ведет к неизбежной гибели структуры (Лоскутов, 1990). В конечном итоге, без воздействия внешнего фактора (в данном случае очередного пожара) ценопопуляции рассмотренных видов деревьев как временные элементы сменяются в процессе развития системы новыми более устойчивыми ценопопуляциями.
Подобным образом может проходить динамика ценотических групп деревьев и для других видов, например, для дуба монгольского, который на территории Сихотэ-Алинского биосферного заповедника входит в состав широколиственно-кедровых лесов, а также образует устойчивые ценотические системы в нижнем лесном поясе. Рассматриваемые ценопопуляции дуба растут в сообществах, находящихся на разных стадиях развития - в березняке разнотравно-папоротниковом и кедрово-дубовом лесу. В поясе кедрово-широколиственных и кедрово-елово-широколиственных лесов дуб монгольский может входить в сообщества как на ранних стадиях сукцессий, так и в виде субдоминанта в коренные древесные ценозы (Флягина, 1982; 1990; 1993).
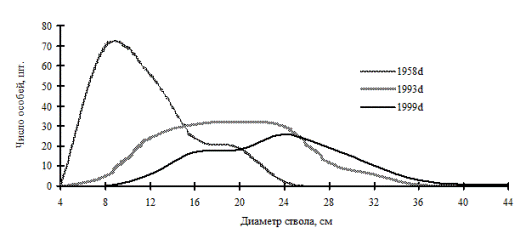
Рис. 31. Частотные распределения березы плосколистной в березняке разнотравно-кустарниковом (пп З-7) на территории Сихотэ-Алинского биосферного заповедника |
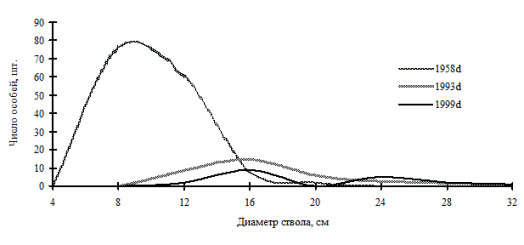
Рис. 32. Частотные распределения осины Давида в березняке разнотравно-кустарниковом (пп З-7) на территории Сихотэ-Алинского биосферного заповедника |
Ценопопуляция дуба в березняке разнотравно-осоково-папоротниковом во все периоды наблюдений постоянно сокращала свою численность, при этом кривая распределения особей по толщине постепенно приобретала правостороннюю асимметрию (рис. 34). Хотя в отдельные периоды ценопопуляция этого вида и распадалась на дискретные группы угнетенных, индетерминантных особей и особей лидеров, группы эти не были устойчивы и исчезали на момент следующих ревизий, так что совокупность деревьев снова приобретала однородное, недискретное строение. По-видимому, такое строение характеризует переломные моменты в динамике ценопопуляции, когда по мере роста деревьев в толщину происходит перестройка ценотических классов деревьев.
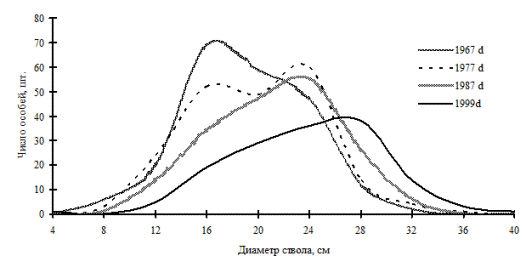
Рис. 33. Частотные распределения березы плосколистной в березняке разнотравно-осоково-папоротниковом (пп М-1) на территории Сихотэ-Алинского биосферного заповедника |
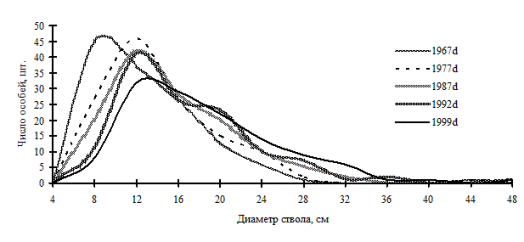
Рис. 34. Частотные распределения дуба монгольского в березняке разнотравно-осоково-папоротниковом (пп М-1) на территории Сихотэ-Алинского биосферного заповедника |
Ценопопуляция дуба из кедрово-дубового леса на момент первого описания имела сложное неоднородное строение по диаметру. Статистическое распределение при этом имело правостороннюю асимметрию (рис. 35). За время наблюдений эта ценопопуляция постоянно сокращала свою численность, но при этом оставалась сложно дифференцированной и лишь в момент ревизии в 1978 г. она приняла более однородную форму. Вероятно, этот переходный момент был связан с отпадом деревьев в группе лидеров, зафиксированным в 1968 г. Хотя дуб монгольский можно отнести к коренным породам деревьев, способным произрастать в устойчивых древесных ценозах без пополнения тонкомерной части древостоя, ценопопуляции этого вида постепенно могут терять лидирующую позицию, что приводит к потере устойчивости и разрушению структуры.
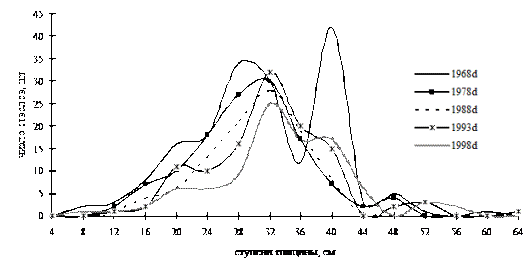
Рис. 35. Частотные распределения кедра корейского в кедрово-дубовом мелкотравно-мертвопокровном лесу (пп З-2) на территории Сихотэ-Алинского биосферного заповедника |
Помимо смешанных сообществ дуб монгольский в Сихотэ-Алинском биосферном заповеднике образует дубовые леса приморской полосы. Считается, что почти все они пирогенного происхождения. Но так как пожары случаются здесь регулярно, то эти вполне сложившиеся флористически и ценотически леса можно рассматривать как весьма устойчивые ценотические системы (Колесников, 1938, 1969; Куренцова, 1939; Васильев, 1948; Дылис, Виппер, 1953; Соловьев, 1958; Шеметова, 1975; Добрынин и др., 1992; Кудинов, 1993; 1994; Добрынин, 1999; Петропавловский и др., 1999, и др.). Дубовые леса порослевого происхождения образуют в основном однородные по толщине древостои и не отличаются ценотическим разнообразием. В отличие от них, дубняки семенного происхождения, которые отнесены И.А. Флягиной (1990) к условно коренным типам леса, могут образовывать устойчивые во времени ценотические группы деревьев. Ценопопуляция дуба монгольского в дубняке лещиновом разнотравно-папоротниковом на момент первого описания в 1986г. была разделена на два ценотических класса (рис. 36), которые сохранились и в момент следующей ревизии в 1998г. |
За этот период произошли лишь сокращение общей численности ценопопуляции дуба и смена максимума в группе лидеров за счет прироста деревьев этой группы по диаметру. Подобный характер динамики выработанной ценотической структуры можно наблюдать для ценопопуляции кедра корейского в кедрово-дубовом лесу (рис. 37). Во все периоды наблюдений эта ценопопуляция также сохраняла определенный характер распределения деревьев по толщине на всех этапах наблюдений. Деревья в разных группах прирастали равномерно, но при этом ценопопуляция кедра сокращала общую численность. Можно предположить, что такой характер существования ценотических групп говорит об упорядоченности функциональных элементов во внутренней структуре ценопопуляции. Очевидно, такая упорядоченность может существовать некоторое время без самоусложнения в состоянии динамического равновесия. Дальнейшая судьба обеих ценопопуляций зависит от того, смогут ли они со временем поддерживать такое состояние равновесия за счет присоединения тонких деревьев, в противном случае может произойти разрушение образовавшейся структуры, и это снова приведет систему в состояние хаоса.
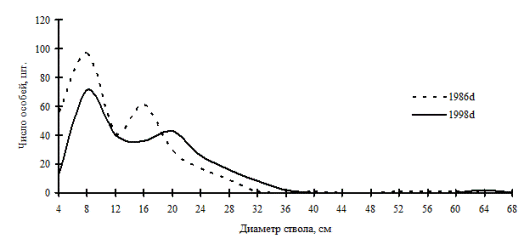
Рис. 36. Частотные распределения дуба монгольского в дубняке лещиновом папоротниково-разнотравном (пп Б-5) на территории Сихотэ-Алинского биосферного заповедника |

Рис. 37. Частотные распределения кедра корейского в кедрово-дубовом мелкотравно-мертвопокровном лесу (пп З-2) на территории Сихотэ-Алинского биосферного заповедника |
На основании всех рассмотренных выше примеров можно сделать предположение: для того, чтобы определенный вид мог занимать устойчивое положение в древостое, он должен не только выработать определенную устойчивую структуру своей ценопопуляции в виде совокупности разных ценотических групп деревьев, но и постоянно пополнять их, а это может происходить только за счет поступления новых элементов из подроста. В этом, по-видимому, и выражается гармония во внутреннем строении древостоя по диаметру. Подрост под пологом древостоя – это один из лабильных элементов структурной организации биогеоценоза. В конкретных типах леса он представляет собой тот относительно стабильный резерв, индивиды которого в процессе борьбы за существование постепенно или в короткий срок сменяют материнские поколения (Бузыкин и др., 1985).
В то же время, за счет присоединения новых элементов в самоорганизующемся древостое может наступить максимальный беспорядок, например, за счет резкого увеличения численности тонкомерной части ценопопуляций деревьев. Примером этого могут служить ценопопуляция лиственницы даурской в дубняке рододендроновом и пихты белокорой в кедрово-пихтовом лесу, произрастающих на территории Сихотэ-Алинского биосферного заповедника. В первом случае увеличение численности тонкомера в ценопопуляции лиственницы не дает ей возможности выйти из состояния однородности по толщине (рис. 38). Результаты первой ревизии 1992г. показывают, что частотное распределение особей данного вида уже начало менять свой характер по мере роста деревьев в толщину, а на момент последней ревизии 1998г., за счет резкого поступления тонких особей из подроста система вернулась к начальному характеру однородности. У ценопопуляции пихты белокорой наблюдается другая ситуация (рис. 39). На первых этапах наблюдений и ревизий, вплоть до 1993г., данная ценопопуляция имела однородный характер распределения и небольшую численность, а в 1993г. и вовсе была представлена единичными особями. Но за пять лет до момента последней ревизии в 1998г. ценопопуляция пихты увеличила свою численность за счет подроста, и это привело в состояние хаоса не только ценопопуляцию этой породы, но и весь древостой.
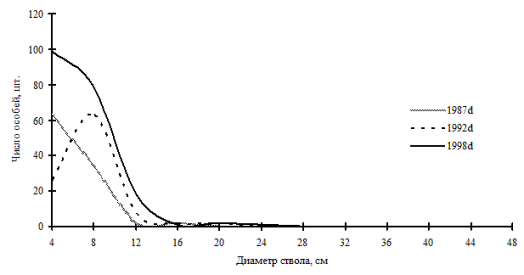
Рис. 38. Частотные распределения лиственницы даурской в дубняке рододендроновом (пп Б-4) на территории Сихотэ-Алинского биосферного заповедника |
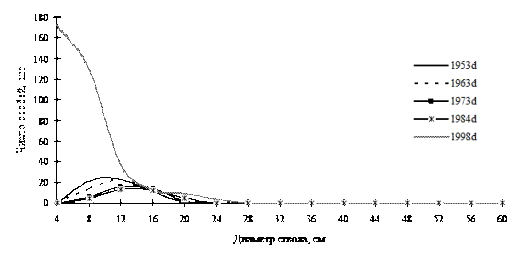
Рис. 39. Частотные распределения пихты белокорой в кедрово-пихтовом с елью и дубом мелкотравном лесу (пп З-4) на территории Сихотэ-Алинского биосферного заповедника |
Как известно, жизнь течет неравномерно, спокойные периоды сменяются напряженными состояниями – состояниями, когда делается выбор, каким будет дальнейший путь. В такие моменты большее влияние на будущее системы оказывает не порядок в системе, а хаос. И без этой неупорядоченной, неконтролируемой, случайной компоненты в динамике системы были бы невозможны ее качественные изменения. Выходит, что при всем стремлении к упорядочению какая-то доля хаоса для эволюции жизни просто необходима (Баранцев, 2003).
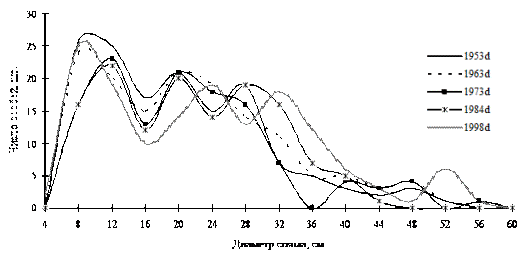
Рис. 40. Частотные распределения кедра корейского в кедрово-пихтовом с елью и дубом мелкотравном лесу (пп З-4) на территории Сихотэ-Алинского биосферного заповедника |
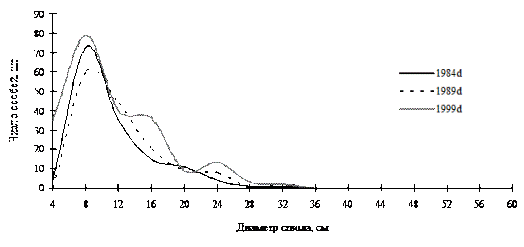
Рис. 41. Частотные распределения пихты белокорой в пихтово-кедровом мохово-мертвопокровном мелкотравно-папоротниковом лесу (пп Ф-1) на территории Сихотэ-Алинского биосферного заповедника |
В наших примерах хаос служит на благо рассмотренным ценопопуляциям деревьев: в благоприятных для себя условиях они смогут усилить свои позиции внутри фитоценоза и прийти к состоянию устойчивости за счет образования ценотических групп деревьев. Однако некоторые ценопопуляции деревьев, по-видимому, могут пребывать в состоянии неустойчивости, недискретности длительное время, на протяжении всей жизни древостоя. Примером этого могут служить дубняки порослевого происхождения или дубняки, произрастающие в экстремальных условиях на морских террасах в Сихотэ-Алинском биосферном заповеднике. Как показывают результаты наших исследований, процесс дифференциации особей в сообществах с однородной структурой проходит по одному сценарию, а в сообществах неоднородных - по другому, более сложному сценарию.
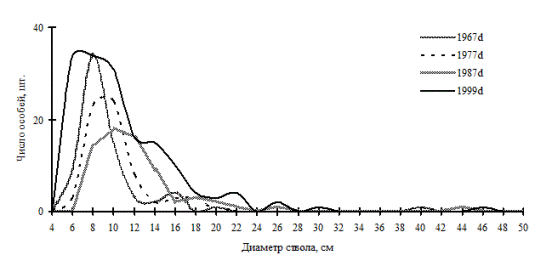
Рис. 42. Частотные распределения кедра корейского в березняке разнотравно-осоково-папоротниковом (пп М-1) на территории Сихотэ-Алинского биосферного заповедника |
Если скорость роста числа новых элементов невелика, то независимо от начальной численности через определенное время установится стационарное состояние (Лоскутов, 1990). Это означает, что присоединение новых элементов для сохранения существующей структуры не должно превышать определенного предела. В нашем случае этот предел должен соответствовать ресурсному потенциалу местообитания. Вероятно, находясь в состоянии, близком к динамическому равновесию, ценопопуляции деревьев сохраняют свой внутренний порядок в распределении по толщине при постепенном увеличении численности, что способствует гармонии во внутренней структуре в виде постоянного существования трех ценотических классов. При этом происходит постепенное усложнение этой структуры.
Примерами таких ценопопуляций могут служить кедр корейский в кедрово-пихтовом лесу; пихта белокорая в пихтово-кедровом лесу, а также кедр корейский в березняке разнотравном в Сихотэ-Алинском биосферном заповеднике. Все эти ценопопуляции находятся в состоянии динамического равновесия: динамика их ценотической структуры проходит по типу постепенного присоединения новых элементов в количестве, необходимом для поддержания и усложнения существующей неоднородности во внутренней структуре (рис. 40-42). |
3.1. Закономерности отпада деревьев
в разных ценотических классах
Динамика древостоя – процесс ветвящийся, дихотомический, поскольку каждое дерево в каждый момент роста может или продолжать расти, или отмирать. Структура древостоя в целом представлена двумя наборами чисел. Первый из них показывает, сколько деревьев относятся к i-му классу толщины, второй – сколько в этом классе толщины деревьев погибло за некоторый промежуток времени. Малое число повторных наблюдений не позволяет выяснить внутренние закономерности динамики древостоев, связанные с экологическим временем. Однако появляется возможность сравнить интенсивность потоков в разных направлениях по дифференциальным характеристикам, найти функциональные зависимости темпов отмирания деревьев от темпов их роста в каждом классе толщины (Черкашин, 1986).
Как известно, по мере роста особей деревьев их потребность в ресурсах увеличивается, и в связи с этим конкуренция между ними ужесточается. Это приводит к тому, что часть особей на площади фитоценоза становится лишней, так как местообитание может обеспечить рост и развитие уже только части из них. Максимальная густота – это критическое состояние ценоза, дальнейший рост которого включает механизм регулирования численности деревьев в сторону ее снижения. Естественные механизмы регуляции приводят численность ценоза в соответствие с экологическими условиями и ресурсами среды (Эйтинген, 1949, 1950; Проскуряков, 1971; Бузыкин и др., 1985).
В результате конкуренции за ресурсы внешней среды изначальная неодинаковость деревьев усиливается, часть особей ценопопуляции гибнет, остальные же растут и дифференцируются дальше. По мере роста оставшихся в живых особей снова наступает перенаселение, и снова часть более слабых из них выпадает, давая возможность расти и развиваться оставшимся. Очень важно проследить, как идет отбор в ценотических группах по мере существования древостоя. К сожалению, на тех участках леса, где нами проводилась ревизия, изначально не были учтены сухие деревья, поэтому мы не смогли проследить динамику отпада деревьев.
Лесообразовательный процесс представляет собой динамическое единство поселения и естественного отпада растений. Если бы вероятность элиминации особей не зависела от размера площадей питания и взаимодействия деревьев в ценозе, т.е. естественное изреживание носило стохастический характер, то эффект изреживания сводился бы к перемещению кривых распределения с возрастом вправо при сохранении их подобия. Стохастическая форма отпада в той или иной мере имеет место в любом ценозе и характерна для разреженных группировок древесных растений. Поскольку кривые распределения не только смещаются, но и деформируются с возрастом, то это свидетельствует о существовании в общем процессе естественного изреживания помимо стохастической еще и дискриминационной формы элиминирования угнетенных деревьев. В этом случае в ценотической группе деревьев элиминируют преимущественно особи с малыми площадями питания. Только изменяя площадь питания особей в загущении вырубкой отдельных деревьев, можно увеличить выживаемость остающихся особей (Эйтинген, 1949).
Как известно, на протяжении всей жизни древостоя существует как дискриминационный, так и стохастический типы отпада. Дискриминационный тип присутствует на протяжении всего существования древостоя, но к концу жизни преобладает стохастический тип отпада (Проскуряков, 1971; Дыренков, 1984; Бузыкин и др., 1985). Как известно из литературных данных, стохастический отпад деревьев происходит во всех классах толщины. В связи с этим может возникнуть вопрос - какие закономерности дискриминационного отпада можно наблюдать в системах, находящихся в состоянии хаоса, и в системах, находящихся в состоянии динамического равновесия? Происходит ли он непосредственно внутри ценотических классов, или конкуренция идет между особями разных классов?
Наши данные показывают, что существует несколько форм дискриминационного отпада. Их можно встретить в древостоях, находящихся как в состоянии динамического равновесия, так и в разной степени хаотичности. Общими закономерностями для всех таких рассмотренных нами древостоев является максимальный избирательный отпад в группе угнетенных и преобладание стохастического отпада в группе лидеров, независимо от степени устойчивости и развития древостоя.
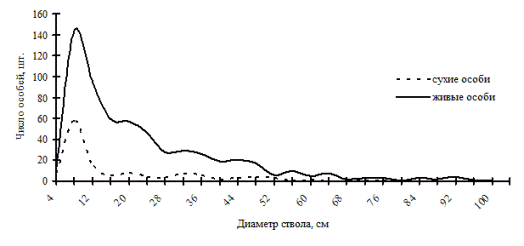
Рис. 43. Частотные распределения древостоя в кедровнике с елью и липой разнотравном (01-2002) на территории заповедника «Бастак» в 2002г. |
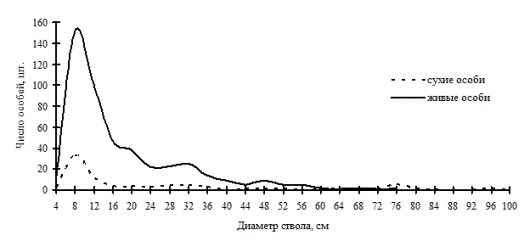
Рис. 44. Частотные распределения древостоя пихтово-кедрового леса (02-2002) на территории заповедника «Бастак» в 2002г. |
В высокодифференцированных древостоях, поддерживающих состояние упорядоченности в строении по толщине, избирательная форма отпада может проходить внутри образованных групп, но основной процесс такого типа отпада происходит в группах угнетенных деревьев и в группе индетерминантных особей. В группе лидеров преобладающей формой отпада является стохастическая. По-видимому, совпадение пиков на обоих кривых может говорить о некоторой стабильности существования групп внутри древостоя: отпад идет строго внутри групп. Примерами таких древостоев могут служить древостои в кедровнике с елью и липой и в пихтово-кедровом лесу (рис. 43-44), наблюдаемые в заповеднике «Бастак», а также древостои в пихтово-кедровом и кедрово-пихтовом лесу (рис. 45-46) в Сихотэ-Алинском биосферном заповеднике. Все эти древостои можно отнести к системам, поддерживающим порядок в строении функциональных элементов древостоя в некоторых пределах возмущений. В таких древесных ценозах, по видимому, поддержание состояния равновесия происходит за счет постепенного пополнения в процессе динамики группы тонких деревьев и путем перераспределения особей внутри существующих ценотических групп. Это мы и наблюдали выше при анализе процесса динамики ценотических групп деревьев во времени.
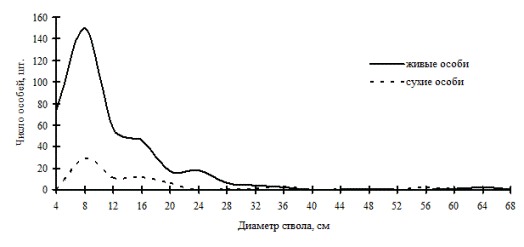
Рис. 45. Частотные распределения древостоя в пихтово-кедровом мохово-мертвопокровном мелкотравно-папоротниковом лесу (пп Ф-1) на территории Сихотэ-Алинского биосферного заповедника |
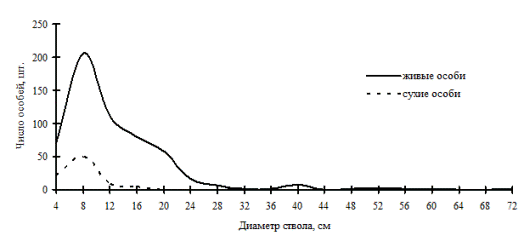
Рис. 46. Частотные распределения древостоя кедрово-пихтово разнотравно-папоротникового леса (пп В-1) на территории Сихотэ-Алинского биосферного заповедника в 2000г. |
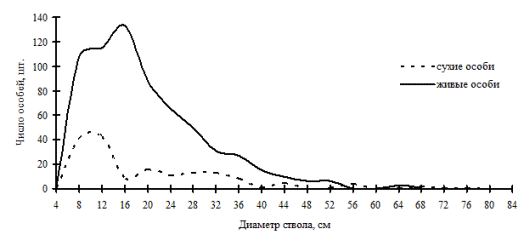
Рис. 47. Частотные распределения древостоя хвойно-широколиственного леса (05-2002) на территории заповедника «Бастак» в 2002г. |
Пример иного типа дискриминационного отпада мы наблюдали в хвойно-широколиственном лесу в заповеднике «Бастак». Этот древесный ценоз относится к сложным многокомпонентным системам, состоящим из нескольких ведущих ценопопуляций разных видов. В этом древостое по числу стволов преобладают пихта белокорая, ель аянская, кедр корейский и липа амурская. Несмотря на многочисленность компонентов, этот ценоз можно охарактеризовать как устойчивую систему, сохраняющую упорядоченность в виде наличия всех ценотических групп во внутренней структуре. Как показывает рис. 47, пики и провалы в частотных распределениях живых и сухих деревьев в правой части кривых не совпадают. Возможно, это и есть тот тип отпада, который происходит при перегруппировке, когда деревья переходят из одной группы в другую, либо когда некоторая часть общей совокупности деревьев распадается на группы по толщине. |
В древостоях, находящихся в неустойчивом состоянии, где доля хаоса имеет различную степень, дискриминационный отпад может идти по-разному, и направлен он большей частью на группу угнетенных деревьев. В древостоях с невыработанной ценотической структурой можно встретить дискриминационный тип отпада, который идет непосредственно по группам, если таковые уже образовались внутри ценоза, либо внутри однородной совокупности, если древостой однороден по толщине. Такую картину отпада мы наблюдали в дубняке разнотравном в Сихотэ-Алинском биосферном заповеднике. Как видно из частотных распределений (рис. 48), кривая сухих деревьев практически повторяет характер кривой для живых особей. В кедрово-пихтовом лесу, пройденном рубками, где древостой неоднороден по толщине и разделен на две группы, отпад непосредственно идет внутри образованных групп (рис. 49). В данном сообществе древостой и микрорельеф сильно нарушены в результате рубки и трелевки вырубленных деревьев. Однако нами учитывались только сухие деревья, стоящие на корню, поэтому отпад оказался немногочисленным.
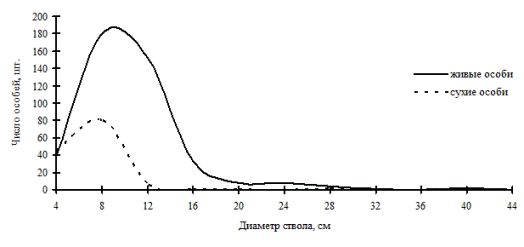
Рис. 48. Частотные распределения древостоя в дубняке рододендроновом разнотравно-осочковом (пп Б-3) на территории Сихотэ-Алинского биосферного заповедника в 1998г. |
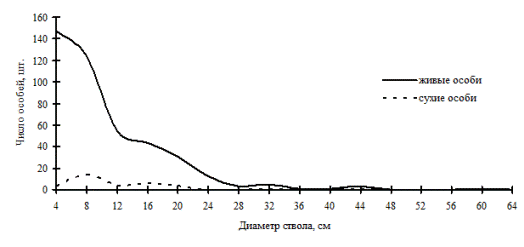
Рис. 49. Частотные распределения древостоя кедрово-пихтового леса (пп В-6) на территории Сихотэ-Алинского биосферного заповедника в 2000г. |
В елово-кедрово-зеленомошном лесу, пройденном рубками (рис. 50), в заповеднике «Бастак» и в березово-лиственничном лесу на месте пожара (рис. 51) в Сихотэ-Алинском заповеднике можно наблюдать еще одну форму избирательного отпада. При общей неоднородности этих древесных ценозов распределение сухих деревьев имеет одно модальное значение в группах тонких деревьев. Обратная картина отпада наблюдается в пихтовом лесу (рис. 52): частотное распределение живых особей имеет один отчетливый максимум в группе угнетенных, максимумы в группе индетерминантных и лидеров только намечаются, а вот кривая сухих деревьев более деформирована. Приведенные примеры показывают разную долю и степень хаоса, а также разные причины, приведшие эти системы в состояние неравновесности. В первых двух случаях это антропогенный фактор, а для пихтового леса - это естественный распад основного древесного яруса. По-видимому, и дискриминационный отпад направлен в таких системах на разные элементы древостоя. В пихтовом лесу такая форма отпада, очевидно, говорит о начале процесса образования ценотических групп, т.е. о процессе самоорганизации древостоя.
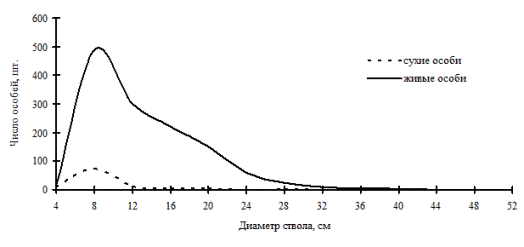
Рис. 50. Частотные распределения древостоя елово-кедрового леса (04-2002) на территории заповедника «Бастак» в 2002г. |
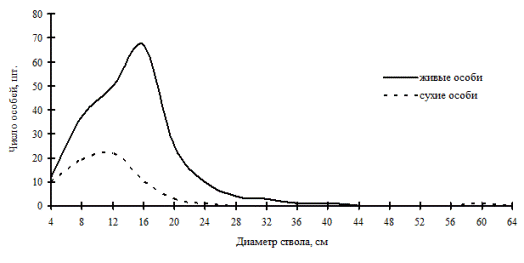
Рис. 51. Частотные распределения древостоя березово-лиственничного леса (пп В-4) на территории Сихотэ-Алинского биосферного заповедника в 2000г. |
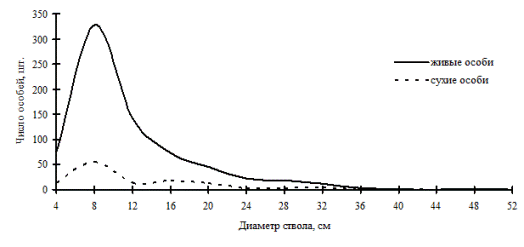
Рис. 52. Частотные распределения древостоя в пихтовом с березой каменной рододендроново-кустарничково-зеленомошном лесу (пп Ф-2) на территории Сихотэ-Алинского биосферного заповедника в 2005г. |
В наших данных можно наблюдать еще несколько примеров избирательного отпада, идущего в неравновесных древостоях, по которым можно судить о переломном моменте в динамике систем. Так, например, в дубняке разнотравно-папоротниковом (рис. 53), который является однородным по толщине и имеет распределение деревьев по толщине, близкое к нормальному, можно увидеть несовпадение кривых распределения. Это произошло по причине наступившего переломного момента, когда деревья из группы угнетенных в процессе роста перешли в группу средних деревьев, а в результате жесткой борьбы выпали в основном тонкие деревья с диаметром ствола от 6,1 до 10см. В отличие от предыдущего ценоза, дубняк лещиновый (рис. 54) показывает более разнообразное ценотическое строение живых деревьев, но при этом здесь мы видим одновершинное распределение сухих особей непосредственно в группе тонких деревьев. Выше мы рассматривали динамику ценотической структуры этого древостоя и видели, что характер распределения деревьев по группам не изменился, т.е. разнообразие функциональных элементов не увеличилось, лишь группа индетерминантных деревьев сместила свой максимум в ступени большего диаметра, а вот основной отпад пришелся на группу тонкомера. Из смешанных древостоев, находящихся в состоянии неравновесия, можно назвать кедровник с дубом (рис. 55), где дуб монгольский теряет свои позиции лидера. |
Теперь в этом ценозе идет смена лидера, сопровождающаяся перестройкой ценотической структуры. Об этом говорит частотное распределение сухих и живых деревьев в этом фитоценозе. Здесь модальные значения кривых мертвых и живых особей не совпадают.
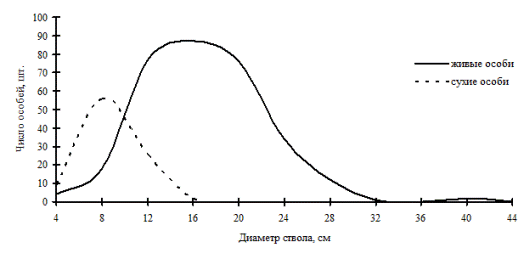
Рис. 53. Общее частотное распределение древостоя в дубняке разнотравно-папоротниковом (пп Б-1) в 1998г. на территории Сихотэ-Алинского биосферного заповедника |
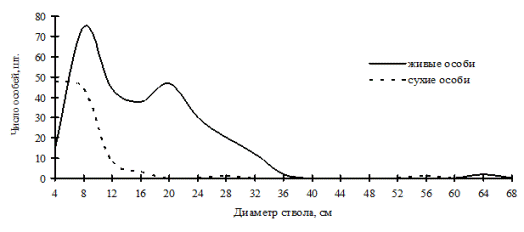
Рис. 54. Частотные распределения древостоя в дубняке лещиновом папоротниково-разнотравном (пп Б-5) на территории Сихотэ-Алинского биосферного заповедника |
Приведем еще один процесс отпада, который идет в результате усыхание дуба монгольского в дубняке разнотравно-осочковом с примесью берез плосколистной и даурской в Сихотэ-Алинском заповеднике. Участок леса, на котором произошло усыхание дуба, расположен рядом с трассой. По всей вероятности, этот процесс как-то связан с изменением гидрологического режима в связи со строительством шоссе, отсыпкой насыпи и расширением дороги. Выросшие в благоприятных гидрологических условиях довольно крупные деревья дуба монгольского в результате ухудшения условий минерального и водного питания оказались не способны жить дальше в новых условиях и стали слабеть. На соседнем участке леса, а также в той части пробной площади, которая примыкает к этому участку, усыхание дуба уже идет независимо от размеров особей. В общей картине элиминации особей дуба здесь наблюдается преобладание отпада в группе угнетенных деревьев (рис. 56), которая, по-видимому, является самым слабым звеном в этой системе особей. Угнетенные особи первыми реагируют на ухудшение условий в данном сообществе.
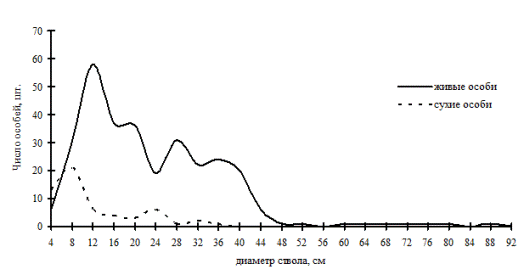
Рис. 55. Частотные распределения древостоя в кедрово-дубовом мелкотравно-мертвопокровном лесу (пп З-1) на территории Сихотэ-Алинского биосферного заповедника в 1998г. |
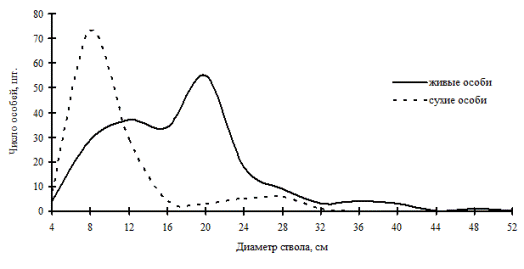
Рис. 56. Частотные распределения древостоя в дубняке разнотравно-осочковом (пп Б-12) на территории Сихотэ-Алинского биосферного заповедника в 1998г. |
Таким образом, в результате антропогенной и естественной динамки во всех рассмотренных нами фитоценозах идет постепенное увеличение ценотического разнообразия древостоев. В процессе самоорганизации древесных ценозов именно те ценопопуляции, которые могут образовывать устойчивые функциональные элементы в виде групп деревьев различного диаметра, имеют шансы занять лидирующее положение в древостое. Однако для того, чтобы древостой мог поддерживать динамическое равновесие, необходимо, чтобы скорость роста числа элементов превышала скорость отмирания старых. По всей вероятности, можно рассматривать и оценивать не только термодинамическое равновесие растительности ландшафтного района в целом, термодинамическое равновесие отдельного контурфитоценоза, но даже равновесие функциональной части древостоя и отдельного ценоэлемента (Галанин, 1989).
