Самоорганизация древесных ценозов
Н. А. Василенко © 2012

Наталья Александровна Василенко
Кандидат биологических наук, участница и организатор многих ботанических экспедиций по Дальнему Востоку и Забайкалью, зав. лабораторией экспериментальной фитоценологии Ботанического сада-института ДВО РАН. Область научных интересов: экспериментальная фитоценология, лесоведение, пространственная структура древостоев, зеленое строительство и ландшафтная архитектура, экологическое образование.
botantehnol@yandex.ru |
"Прежде всего нет смысла говорить о противопоставлении часть - целое. С целым как таковым мы не имеем дело. Какой бы детальный архетип мы не приняли, он не будет состалять целого, но лишь какую-то его часть (аспект). В этом смысле мы не можем оперировать с целым, которое остается в области трансцендентного и доступно лишь созерцанию, интуитивному восприятию, откровению"
С. В. Мейн
Василенко Н.А. Самоорганизация древесных ценозов. Владивосток: Дальнаука, 2008. 171 с.
В книге анализируется формирование и развитие в процессе динамики древостоя различных ценотических групп деревьев, отличающихся по диаметру. Выявлены особенности пространственного размещения деревьев в фитоценозах, в фитогенных полях соседей деревьев. Использованы оригинальные методы, позволяющие уловить пространственный ритм внутри каждого древостоя. Книга написана на материалах, собранных на постоянных пробных площадях в разных типах лесов российского Дальнего Востока.
Рассчитана на геоботаников, экологов, работников заповедников и специалистов лесного хозяйства. Ил. 120, библ. 171.
Vasilenko N.A. Self-organization of forest coenoses. - Vladivostok: Dalnauka, 2008. - 171 p.
The monograhpy is devoted to the issue about various tree groups fprmation and development as well as features of spatial distribution of trees in plant communities. The book is based on original materials of long-term reseach on the constant plots lokated in different nature reservs and various forest types. The constant plots are distribed and treated by inform technique that makes possible to compare the received results the issue of the stand functional elements formation. The numerous data schow that in various stands division of coenotic population of dominants and edificators on discrete groups by diameter can be assumed as a process of self-organizing of plant community as a whole. One of the chapters of the books studies peculiarities of spatial tree distribution in the forest. It is shown that within every stand, specific spatiel rhytm is maintaining together with the formation of the inheren order of functional part.
The book is intendid for geobotanists, ecologists, employees of nature resrves and forestry experts. Ill. 120, bibl. 171, suppl. 1 |
От ответственного редактора
Монография Н.А. Василенко посвящена сложной проблеме самоорганизации древесных фитоценозов. На огромном фактическом материале автор показала, что распределение особей деревьев разной толщины (а следовательно, и разной ценотической мощности) в пределах растительного сообщества относительно друг друга может быть неслучайным. Чаще всего такие неслучайности обнаруживаются в выработанных коренных фитоценозах, в то время как в ценозах серийных и пионерных такие закономерности отсутствуют или едва уловимы. Для доказательства этого факта автор использует серию оригинальных методов, в том числе и методы статистического анализа. Используемый ею метод накопления частот встречи условно подчиненных деревьев в фитогенных полях деревьев лидеров заслуживает большого внимания исследователей и дальнейшего совершенствования.
В конце ХХ столетия фитоценология, в том числе и лесоведение, зашли в теоретический и методический тупик. Идеи, заложенные в основу этих наук еще в конце ХIX в., к этому времени оказались исчерпаны, а новых не появилось. Надежды 50-60-х годов прошлого века на то, что толчок к развитию фитоценологии даст математическая статистика, не оправдались. С помощью точных методов удалось лишь подтвердить некоторые постулаты, которые были сформулированы фитоценологами 100 лет назад, не прибегая к статистике.
Пересмотр ряда основных теоретических положений фитоценологии и всей геоботаники начался в конце 80-х годов ХХ в. Отдельные исследователи усомнились в правильности ряда аксиом геоботаники и начали «штурм ортодоксальной цитадели». Помню, как сложно было опубликовать тогда в журнале статью с выводами, противоречащими положениям «классической» геоботаники. Но тем не менее, такие статьи иногда в журналах и сборниках выходили, факты накапливались, и в 1990-е годы стали появляться работы с попытками сформулировать принципы новой теории растительного покрова. Эти принципы отвергало старшее поколение геоботаников, но приходила молодежь и подхватывала эти идеи.
В начале нового века ушли из жизни геоботаники – строители ортодоксальной теории растительности, постарели и те исследователи, которые начинали раскачивать основы ортодоксальной геоботаники. Появились их ученики, подхватившие новые идеи и развивающие их дальше. Монография Н.А. Василенко является доказательством этого. В этой монографии положения не просто постулируются (это свойственно любой ортодоксальной теории), а тщательно проверяются на фактическом материале. Автор не вступает в бессмысленные схоластические полемики с классиками, а идет от его величества факта. А фактов в книге очень много. Прослежена динамика древостоев на десятках постоянных пробных площадей, заложенных в разных типах леса и в разных регионах Дальнего Востока и Восточной Сибири. Умелое использование возможностей ПК, оригинальные программы анализа конкретных статистических распределений особей в ценопопуляциях многих видов в разных фитоценозах позволили установить, что система фитоценоз обладает свойством самоорганизации. Одним из условий самоорганизации является нахождение системы в точке бифуркации, когда появляется множество возможных выходов системы из неравновесного состояния.
К сожалению, из монографии остается неясно, почему система из множества возможных выходов в точке бифуркации выбирает самый оптимальный, приводящий к повышению ее (системы) негэнтропии. Но это не беда автора монографии, ответ на этот вопрос наукой пока не найден. Возможно, Наталья Александровна сможет найти на него ответ в своих последующих исследованиях и расскажет об этом в статьях и монографиях.
Монография Натальи Александровны будет интересна всем, кто пытается строить и развивать новую теорию растительного покрова: геоботаникам, фитоценологам, экологам, лесоведам. Много полезного в ней найдут преподаватели геоботаники и экологии в вузах, а также любознательные студенты.
Издание монографии интернете, в отличие от бумажного издания, делает ее доступной для неограниченного количества читателей. Хочется надеяться, что все молодые исследователи последуют примеру Н.А. Василенко и станут помещать свои работы (статьи и монографии) в сети интернет. К сожелению, Академия наук все еще тормозит этот процесс, считая публикации в интернет третьестепенными по сравнению с бумажными изданиями. "Бухгалтерский учет" научных публикаций делает свое черное дело – тормозит развитие науки, как только может. А истинная наука – это не учет статей и монографий чиновниками от науки, а обмен мнениями, фактами, теориями и гипотезами самих ученых.
А. В. Галанин
ВВЕДЕНИЕ
Самоорганизация – это наиболее очевидная особенность биологических систем и заключается она в том, что системы способны к образованию и развитию сложных упорядоченных структур. Любой эволюционный процесс выражен чередой смен оппозиционных качеств - условных состояний порядка и хаоса в системе, которые соединены фазами перехода к хаосу (гибели структуры) и выхода из хаоса (самоорганизации). Развитие системы происходит за счет внутренних механизмов, в результате процессов самоорганизации и (или) за счет внешних управляющих воздействий (Аршинов и др., 1994; Буданов, Мелехова, 1999).
Изучение процессов самоорганизации растительных сообществ как части более сложных систем – биоценозов и биогеоценозов - является одним из основных вопросов фитоценологии. Для растительного сообщества, так же как и для всех открытых нелинейных систем, характерны устойчивость относительно внешних воздействий, самообновляемость, возможность к самоусложнению, росту, развитию, согласованность всех составных частей. Изучая сложные самоорганизующиеся системы, необходимо делать акцент на внутренние свойства как на источник саморазвития и познавать функционирования элементов. Например, благодаря дискретности жизни и эколого-биологической неравноценности особей растений, даже на совершенно однородном участке формируется неоднородная система, микроструктура которой обеспечивает более оптимальный ход биогеохимического процесса экосистемы в целом (Галанин, 1989).
Устойчивость является одним из ключевых свойств сообществ. Это состояние в существовании сообщества достигается, с одной стороны, разнообразием входящих в его состав членов, с другой, - наибольшей их приспособленностью как друг к другу, так и к условиям существования. Видовое и ценотическое разнообразие приводят к более устойчивому положению растительного сообщества, позволяя ему существовать длительное время в состоянии равновесия. Сообщество сохраняет свою уникальность, то есть чем-то отличается от других таких сообществ, и сохраняет эти отличия во времени. Поддержанием такого состояния природа демонстрирует нам не просто более высокую степень, а совсем другой уровень сложности. Решения, которые нашла природа за миллионы лет, являются оптимальными, а попытки перекраивания природы в угоду человеку приводят к появлению систем, энергетическая эффективность которых в конечном счете ниже природной (Баранцев, 2003).
Одним из элементов, составляющих растительное сообщество, является древостой, структура которого в основном определят всю структуру лесного фитоценоза. В естественных древостоях наблюдается более широкая амплитуда различий растений по генетическим и фенотипическим признакам, что проявляется в разной индивидуальной силе роста и дифференциации деревьев, сопровождающейся отпадом слабых экземпляров и являющейся основой устойчивости всего фитоценоза. В результате дифференциации особей древостоя в ценопопуляции возникают разные совокупности (группы) деревьев по толщине. Динамика лесного фитоценоза и процесс дифференциации его элементов определяются также и характером их размещения на площади относительно друг друга. Анализ структурных образований коренных и производных насаждений дает большую информацию о природе изучаемого леса и ее подлинном разнообразии. Использованный нами метод достаточно полно отражает ценотическую структуру сообщества.
Целью нашей работы было изучить закономерности формирования и развития различных ценотических групп деревьев, отличающихся по морфологическим показателям, а также особенности их пространственного размещения и динамику, что позволило бы выявить общие закономерности процесса самоорганизации древостоя как основного процесса лесной экосистемы.
Материалом для написания этой монографии послужили исследования, проведенные автором в процессе экспедиционных работ в период с 1997 по 2005гг. С 1997 по 2005гг. исследования проводились на территории Сихотэ-Алинского биосферного заповедника совместно с А.В. Галаниным, А.В. Беликович, Е.А. Пименовой; при участии Г.П. Аверковой, А.И. Недолужко, А.В. Недолужко, Т.А. Кузнецовой, А.Г. Киселевой. Исследования проводились при поддержке гранта Комитета по экологии и природным ресурсам, в рамках раздела № 9 Госпрограммы по биоразнообразию в 1997-1999гг.; гранта ДВО РАН «Выявление упорядоченности в пространственной структуре древостоев в лесах юга Дальнего Востока» в 2002-2003гг. и гранта ДВО РАН «Моделирование пространственной структуры смешанных широколиственно-хвойных лесов юга Дальнего Востока» в 2004-2005гг.
На территории заповедника «Бастак» исследования проводились в 2002г., совместно с П.В. Крестовым и А.П. Добрыниным. На территории Лазовского заповедника исследования проводились в 2005г., при участии И.А. Галаниной и Л.С. Яковченко. Исследования проводились при поддержке гранта ДВО РАН «Моделирование пространственной структуры смешанных широколиственно-хвойных лесов юга Дальнего Востока».
На территории Сохондинского биосферного заповедника исследования проводились в 2005г., при участии Л.С. Яковченко, при поддержке Программы комплексных исследований бассейна реки Амур (Программа ДВО РАН).
На острове Сахалин исследования выполнены в 2005г. совместно с А.В. Галаниным; при участии В.А. Галанина и А.В. Недолужко. Исследования проводились при поддержке гранта ДВО РАН «Экспедиция ботанического сада-института ДВО РАН на Сахалин».
Изучение динамики процесса формирования и развития ценотических групп деревьев по толщине было бы невозможно без наличия систем постоянных пробных площадей, которые существуют на территории Сихотэ-Алинского и Сохондинского биосферных заповедников. Существование систем постоянных пробных площадей стало возможным благодаря работам И.А. Флягиной в Сихотэ-Алинском биосферном заповеднике и А.В. Галанина и А.В. Беликович в Сохондинском биосферном заповеднике. Это огромный массив первичных данных, благодаря которому можно проследить судьбу отдельных фитоценозов, ценопопуляций и отдельных особей деревьев в течение нескольких десятилетий, в отдельных случаях начиная с 1953г.
Компьютерная программа TreeSta 1.1, для выявления пространственного ритма в размещении ценотических групп деревьев была написана И.В. Недолужко (лаб. спутникового мониторинга ИАПУ ДВО РАН). Автор выражает огромную благодарность всем, кто оказывал помощь и содействие в проведении исследований и сборе материалов, а также работникам заповедников, благодаря труду которых сохранились массивы леса, в которых заложены постоянные пробные площади.
Глава 1
РАСТИТЕЛЬНЫЙ ПОКРОВ В РАМКАХ СИНЕРГЕТИЧЕСКОЙ ПАРАДИГМЫ
Смена парадигм в любой науке происходит благодаря обновлению взглядов и методов, которые позволяют получать результаты, ведущие к новым теориям. Появление новых идей и новое понимание традиционных проблем в рамках той или иной науки происходит благодаря накопляющимся годами новым знаниям, не укладывающимся в рамки старых концепций. Многие научные мысли были не закончены из-за недостаточности накопленных знаний или несвоевременности их высказывания.
Грядущий век – это век междисциплинарных исследований. Переход к эволюционной парадигме сейчас связывают с новым осознанием роли человечества в мире в целом. Потребительское отношение человека к окружающему миру, желание потреблять и управлять процессами, происходящими в природе, сменяются желанием созерцать, понимать и создавать. Если дальнейшее видение места человека в окружающем мире будет развиваться в плане наблюдения и гармонии с природой, то вероятно, многие цивилизационные и экологические кризисы можно будет избежать.
Синергетика занимается изучением систем различной природы, состоящих из большого числа частей, компонент или подсистем, т.е. деталей, сложным образом взаимодействующих между собой тем или иным способом. Синергетическая парадигма исходит из идей единства и системной организации всего мира, а также наличия общих законов развития всех материальных объектов и систем, нелинейности и необратимости процессов, протекающих на всех уровнях бытия. Синергетика объясняет, почему в процессах самоорганизации образуются структуры (и именно такие). Кроме того, она раскрывает тот факт, что все структуры имеют эволюционный характер, т.е. представляют собой определенные стадии развертывания процессов (Буданов, 1999). Изучением открытых систем занимаются многие науки, имеющие свои методы, в том числе и фитоценология, объектом изучения которой является растительный покров (Миркин, Розенберг, 1978; Ипатов, Кирикова, 1999).
В каждой науке существует несколько подходов к изучению систем. В одних случаях мнение о структуре в целом составляют через изучение отдельных частей. Другие изучают систему как единое целое, указывая на невозможность существования частей отдельно друг от друга без их взаимодействия. Но можно предположить, что оба подхода имеют право на жизнь. Иногда детерминистский подход бывает просто необходим, например, при изучении функциональных частей системы, каждая из которых несет свои определенные функции в общей работе системы.
Растительный покров - это один из компонентов в структуре биологической системы более высокого уровня - биосферы. Биосфера как глобальная экосистема является обычной незамкнутой термодинамической системой. Основная цель биосферы суши, по всей вероятности, состоит в том, чтобы обеспечить как можно большую степень замкнутости биологического круговорота, захватив в него как можно больше подвижных химических элементов, которым в противном случае угрожает вынос за пределы конкретной экосистемы (Галанин, 1982).
Одна из характеристик биосферы – негэнтропия. Поступая из внешней среды, она является мерой разнообразия и упорядоченности, которые образуется на планете в результате эндогенных процессов. Благодаря негэнтропии каждая экосистема характеризуется своим термодинамическим потенциалом, определяемым ее неоднородностью и неравновесием ее компонентов относительно внешней среды. Понятие «термодинамический потенциал» в экологии пока используется очень редко, чаще используется понятие «эдафический потенциал». Эдафический потенциал экосистемы определяется характером почвы, составом подстилающих почву материнских пород, положением в макро- и мезорельефе, характером микрорельефа и, в связи с этим, характером фито- и зооценоза.
Важной характеристикой растительного покрова, так же как и систем любой природы, является нелинейность происходящих в нем взаимодействий. Она выражается в том, что будущая структура этой системы не предопределена; в ходе развития, находясь в неустойчивом состоянии, она выбирает наиболее оптимальный путь и оптимальную форму самоорганизации, характеризующуюся определенной структурой и порядком элементов. Для растительного покрова одной из таких структур является распределение видов конкретной флоры по конкретным местообитаниям на ландшафтно-экологическом уровне, а также возникновение определенного набора функциональных частей внутри каждого растительного сообщества.
Что касается элементов, то в процессе эволюции каждый вид растения выработал свою «стратегию жизни». Это выражается в способности входить в состав то большего, то меньшего числа типов фитоценозов, занимая в них различное положение. Возникли виды, способные в благоприятных для них условиях доминировать и оказывать большое средообразующее влияние, а также виды, способные доминировать, не влияя существенно на среду, и виды, не способные доминировать, занимающие второстепенное положение в фитоценозах (Работнов, 1992). Эти типы стратегий и приспособлений дают возможность одним растениям разных видов обитать совместно друг с другом и с организмами животных, грибов и микроорганизмов, а другим растениям нет. Все это необходимо для достижения равновесия растительности и среды, но равновесие в реальном растительном покрове никогда не бывает полным, так как экологическая система ландшафтного района постоянно выводится из равновесия разными факторами и разными способами: изменением климата, деятельностью человека, заносом новых видов растений и животных и т.д. Такое состояние неравновесности в системе в синергетике принято называть точкой бифуркации. Из этого состояния неустойчивости система выбирает оптимальный для нее путь развития. Для растительного сообщества стремление к наиболее устойчивому состоянию иногда достигается путем изменения его исходной структуры.
Выбор стратегии поведения предполагает существование целей, которые стремится достичь система. «Аттрактором», или целью для системы, является весь возможный набор элементов и относительно устойчивых состояний, которые могут реализоваться в зависимости от тех или иных начальных условий (Князева, Курдюмов, 1994). Считается, что стратегической целью биологических систем является стремление к выживанию самим или способствование выживанию систем вышестоящего уровня (Миркин, Розенберг, 1978). Это достигается путем подбора подходящих компонентов экосистем, например, для фитоценозов – это, в первую очередь, отбор видов из как можно большего числа потенциальных претендентов, а не только видов, способных сформировать на данном участке биогеохимически оптимальный фитоценоз. Благодаря этому при изменении условий местообитания видовой состав фитоценоза может быстро изменяться. Это важно при любом изменении термодинамического потенциала местообитания.
Определенный доступный сообществу видовой состав для каждой конкретной экосистемы является «аттрактором» или экологическим потенциалом конкретной флоры. Если фитоценоз выведен из равновесного состояния, то он становится открытым относительно этого потенциала. Такой подбор видов в фитоценозы происходит поэтапно в ходе сукцессии и является эколого-ценотическим процессом, процессом биогеохимической настройки экосистем, их биогеохимической оптимизации (Сукачев, 1928; Галанин, 1982; Селедец, Пробатова, 2007). Самыми оптимальными с точки зрения устойчивости являются многовидовые сообщества. В отличие от одновидовых у них существует больше путей решений; будучи выведены из состояния равновесия, они быстрее перестраивают свою биогеохимическую работу и возвращаются в состояние, близкое к исходному, либо полностью меняют свою структуру для достижения нового равновесного состояния. Такие многокомпонентные системы отличаются еще и наличием большего числа ценоэлементов во внутренней структуре сообщества, но это уже относится не к таксономическому, а к ценотическому разнообразию системы.
Чем больше у системы степеней свободы, тем более она способна к «самоподтягиванию» и самоусложнению, повышению уровня упорядоченности. Неравновесность и нестабильность системы, наличие в ней множества точек бифуркаций далеко не всегда ведут к ее разрушению. С точки зрения синергетики состояние хаоса не является чем-то вредным и абсолютно разрушительным, так как в это время система выбирает различные варианты самоорганизации и останавливается на более оптимальном. Иногда отношению понятий “хаос” и “порядок” придается самодостаточный смысл, причем не только в синергетике. Однако в природе нет абсолютного хаоса и абсолютного порядка. Мера упорядоченности (или хаотичности) может возрастать по какому-либо показателю за счет снижения меры упорядоченности (или хаотичности) по другому показателю (Князева, Туробов, 2002).
«Порядком» для экосистемы является поддержание биологического круговорота в таком состоянии, чтобы вещество возвращалось обратно к растениям почти полностью и почти полностью включалось ими в новый цикл круговорота, а не рассеивалось за пределами экосистемы (May, 1976; Breymeyer, 1979; Галанин, 1989; Работнов, 1998). Упорядоченность в растительном покрове носит сложный многоуровневый характер и реализуется как в пространстве, так и во времени в виде определенного набора состояний и существовании различных элементов во внутренней структуре.
Неустойчивые динамические системы являются весьма податливыми и чрезвычайно чувствительными к внешним воздействиям. Благодаря этому развивающаяся структура характеризуется возможностью перейти в одно из очень большого числа допустимых равноправных состояний (Лоскутов, 1990). Если имеют место малые возмущения начальных условий, то поведение такой системы иногда можно предсказать, зная начальное распределение элементов. В противном случае процесс может быть описан только вероятностным образом. Это правило применимо и для биологических систем, в том числе и для растительного покрова (Сукачев, 1954; Миркин, Розенберг, 1978).
Растительный покров является скорее процессом, нежели объектом, и самое интересное в нем – его динамика (Галанин, Беликович, 2004). Любые нарушения, вызванные внешними факторами или изменениями внутренней среды, приводят к закономерной динамике структуры сообщества. Динамика структуры или сукцессия есть ряд смен отдельных стадий развития сообщества. Смена ассоциаций закономерна и фиксирована для данного типа сообщества и характера нарушения. Именно в процессе такой динамики идет выработка упорядоченности в структуре сообществ, и выявляются закономерности его самоорганизации.
Каждая система в процессе динамики выбирает для себя оптимальную в данных условиях форму самоорганизации и «сбрасывает» в окружающее пространство лишнюю энергию, не нужную для образования и функционирования новой структуры. Этот процесс называют диссипативным. Синергетика исходит из того, что общее направление дальнейшего развития системы при некоторых условиях может определить случайность. Иначе говоря, когда система находится в состоянии неустойчивости (в точке бифуркации), какое-либо необязательное (однозначно не предопределенное) внешнее событие (воздействие) может столкнуть систему в состояние «творческого хаоса» и определить ту структурно-организационную форму, которую она далее примет (Лоскутов, 1990; Князева, Туробов, 2000).
Глава 2
ЦЕНОТИЧЕСКИЕ ГРУППЫ ДЕРЕВЬЕВ И ИХ РОЛЬ В ПРОЦЕССЕ САМООРГАНИЗАЦИИ
Главной компонентой биогеоценозов (экосистем) являются сообщества растений, они являются основными поставщиками органического вещества и определяют общий облик биогеоценозов. При применении детерминистического подхода к изучению растительного покрова выделяют функциональные структурные части растительных сообществ – ярусы, ценоячейки, синузии, которые возникают в результате дифференциации растительного покрова на ценотическом уровне организации (Шенников, 1934; Ниценко, 1971 , Норин, 1980; Ипатов, Кирикова, 1999). Образование таких внутренних элементов в системе есть результат ценофункциональных связей, а наличие таких элементов отражает ценотическую организацию растительного покрова. Такая внутренняя структура с ее элементами и связями стремится к самосохранению в некоторых границах допустимых возмущений (Галанин, 1982).
Ценотическая организация растительного покрова является результатом взаимодействия особей путем обмена веществом и энергией с внешней средой и через нее друг с другом. Биоценотические отношения связывают между собой все организмы биоценоза, ставят эти организмы в значительной степени в определенные отношения относительно друг друга, т.е. делают биоценоз и фитоценоз упорядоченными системами, развивающимися во времени (Сукачев 1956, 1964).
Определенное сочетание ценоэлементов приводит к образованию различных форм ценотической организации растительного покрова. В растительном покрове имеются ведущие ценоэлементы и подчиненные, играющие в экосистеме второстепенную роль. Качество и степень воздействия в основном зависят от эдификаторного вида, образующего ведущие ценоэлементы, которые в свою очередь определяют облик подчиненных ценоэлементов.
Одним из элементов, составляющих растительное сообщество, является древостой, структура которого в основном определяет всю ценотическую структуру фитоценоза.
Как уже отмечалось выше, в природе встречается большое разнообразие условий произрастания и связанных с ними растительных сообществ, в связи с этим следует ожидать разнообразия функциональных элементов, в том числе и многообразия строения древостоев (Колесников, 1966; Макаренко, 1970).
В начале формирования древостоя имеют значение наследственные различия растений по скорости роста, различия в размере семян, из которых они возникли, а также и различия в микроусловиях произрастания отдельных экземпляров и времени их появления. Но наиболее мощным на протяжении всего периода существования деревьев, хотя и затухающим после периода большого роста, остается фактор взаимодействия.
Дифференциация древостоев по диаметру деревьев - вопрос сложный из-за наличия целого комплекса факторов, влияющих на распределение числа деревьев в зависимости от толщины ствола. Среди этих факторов - условия местопроизрастания, возраст, высота, породный состав, густота (полнота), хозяйственная деятельность человека в лесу, загрязнение окружающей среды, стихийные бедствия и др. (Розенберг, Манько, 1959; Никитин, 1966; Лебков, 1967; Мошкалев, Книзе и др., 1973; Макаренко, 1975; Калинин, 1978, и др.). Изначальная неодинаковость часто связана с различными наследственными свойствами особей и, как следствие, с разными требованиями их к условиям внешней среды (Марков, 1975; Кузьмичев и др., 1989).
Годичные кольца – приросты древесины - рассматриваются в качестве своеобразной летописи смены условий, сопутствующих дереву в течение его жизни. Диапазон факторов, которые могут повлиять на годичный прирост, очень широк, поэтому невозможно выявить наличие прямой зависимости от одного фактора в отдельности. Ширина годичных колец древесины зависит от вида древесной породы, возраста и происхождения дерева, а также от внешних условий и других факторов (Лобжанидзе, 1961; Таранков и др., 2004).
На характере распределений деревьев по толщине также отражаются общественные отношения деревьев (Кищенко, 1926; Третьяков, 1927; Ипатов 1967, 1968, 1969, 1971). Н.В. Третьяков (1927) пришел к выводу о том, что вся внутренняя гармоничная структура древостоя характеризуется рангами деревьев по толщине ствола. Для выяснения закономерностей распределения деревьев в насаждениях, для учета влияния среды и возраста на их распределение, развитие и размеры, для установления показателей роста деревьев разные исследователи предлагали различные классификации деревьев в лесу. В первую очередь, это традиционная классификация Крафта (Воронов, 1973), различные ее доработки (Нестеров, 1954; Высоцкий, 1962; Тарасов, 1968; Ипатов, 1968, 1969; Пугачевский, 1983; Бебия, 2000; Ипатов, Кирикова, 2001). В.С. Ипатов смог статистически доказать наличие трех дискретных совокупностей деревьев в древостое. Математический анализ кривых распределения деревьев по размерам показал, что объективно в древостое в пределах фитоценоза формируются разные ценотические классы: угнетенные, индетерминантные и господствующие деревья (Ипатов, 1967, 1968, 1969, 1970).
В.Е. Барнацкий (1968) установил, что при пропитке живых деревьев бука водным раствором сернокислого железа и меди происходит перераспределение растворов, при этом обнаруживаются деревья-поставщики (лишающиеся этих растворов), деревья-потребители и нейтральные деревья. Видимо, первые – угнетенные, вторые – господствующие, третьи – индетерминантные (т.е. с равными потребностями).
В системе отношений особей деревьев можно обнаружить несколько типов взаимодействий, как-то: 1 – «лидер – лидер», 2 – «лидер – отстающий», 3 – «отстающий – отстающий», 4 - «отстающий – аутсайдер», 5 – «лидер – аутсайдер», 3 – «перспективный аутсайдер - безнадежный аутсайдер». Следует заметить, что каждая группа особей деревьев в фитоценозе является естественной и занимает свою эколого-ценотическую нишу. Разнообразие потребностей позволяет растениям по-разному и благодаря этому более продуктивно для растительного сообщества использовать определенный экотоп (Плотников, 1974; Костенчук, 1985). Так, например, в годы с благоприятными для роста растений условиями наибольший вклад в работу экосистемы вносят мощные деревья лидеры. При этом они становятся ценотически очень активными, быстро растут и сильно угнетают особей-аутсайдеров. В годы с неблагоприятными для роста этих растений погодными условиями, напротив, быстрее растут особи-аутсайдеры, так как имеющихся в экосистеме ресурсов для успешного функционирования особей лидеров не хватает, но их достаточно для роста и развития особей-аутсайдеров (Галанин и др., 2000). По всей вероятности, разделение ценопопуляции доминанта и эдификатора на дискретные группы по эколого-ценотической мощности можно рассматривать как процесс самоорганизации фитоценоза. В этом случае увеличивается биоразнообразие сообщества, а каждая группа деревьев «уходит» в свою экологическую нишу (Галанин, Беликович, 2002; Галанин, 2003; Галанин, Беликович, 2004).
Исследования некоторых авторов свидетельствуют, что «индивидуальная сила» деревьев определяется уже в фазе первичного роста лесных культур (когда конкуренция еще слаба), т.е. уже в это время формируется социальная структура будущего древостоя (Сукачев, 1953; Мослаков и др., 2001). Эти авторы считают, что фенотипическое и классовое разнообразие популяции деревьев, образующих древостой, есть не столько продукт конкурентных отношений между ними, сколько продукт генетического потенциала и уже существующего первичного эндогенетического разнообразия и расслоения всей популяции (Марков, 1989; Мослаков, 1996).
Несмотря на разнообразие факторов, влияющих на образование ценотических групп деревьев по толщине, а также на разнообразие строения древостоев по этому признаку, при анализе рядов распределения деревьев по толщине можно обнаружить общие закономерности в формировании и существовании данных функциональных образований во внутренней структуре древостоев. Эти сходные тенденции обусловлены взаимовлиянием в конкретный промежуток времени таких структуроформирующих процессов леса, как физический рост деревьев, их отпад и интенсивность внедрения в полог молодых особей из подроста (Рыжков, 2000).
Для изучения закономерностей распределения деревьев по диаметру мы использовали традиционные 4-х сантиметровые ступени толщины, принятые в лесоведении (Загреев, Вагин, 1975).
2.1. Социальная структура древостоев с малым числом элементов и малой долей участия
Как показывают многочисленные исследования, изначально древостой однороден (т.е. по толщине деревьев не дискретен!) (Поликарпов, 1962; Калинин, 1962,1978; Макаренко, 1963, 1970а, б; Иванюта, 1964; Рубцов, 1964; Смирнов, 1965, 1970, 1972; Захаров, 1967; Григалюнас, 1967а, 1968; Старостенко, 1968; Моисеев, 1971; Ипатов, 1971; Гнилякович, 1971; Багинский, 1972; Успенский, Попов, 1974, и др.). Такое строение достигается за счет высокой численности деревьев сходных размеров, и в древостое преобладает группа тонкомера. Такую картину мы наблюдали на месте сплошной вырубки, где идет формирование лиственнично-кедрового леса (рис. 1), и древесный полог находится на стадии формирования (Галанин, Беликович, 2004). Та же картина наблюдалась в дубняках, которые носят отчетливые следы повреждения пожарами (рис. 2) - порослевой характер стволов, пожарные подсушины, участие в составе пионерных видов, таких как лиственница (Флягина, 1993; Галанин и др., 2000), либо произрастающих в суровых экологических условиях (сильные ветра и близость моря) (рис. 3). Аналогичная картина наблюдалась на участке пихтового леса, где в результате распада древостоя из кедра и ели (Галанин и др., 2000) активизировалась и на какой-то период времени заняла ведущую позицию ценопопуляция пихты белокорой (рис. 4). Однородную структуру можно также наблюдать в отдельно взятых ценопопуляциях, например, ценопопуляции пихты белокорой при внедрении ее в основной древостой в ходе восстановительной динамики на месте кедровника (рис. 5), а также при увеличении численности ценопопуляции этого же вида в результате распада основного яруса кедра в кедрово-пихтовом лесу (рис. 6).
Все эти примеры состояний древостоев или отдельных их частей можно отнести к нахождению этих фитоценозов в точке бифуркации. Это и есть то состояние хаоса, в котором находятся все эти системы и подсистемы, и дальнейший путь к той или иной структуре у каждой из них индивидуален. Факторами, которые были причинами, подтолкнувшими рассмотренные нами древостои и отдельные ценопопуляции в состояние «творческого» хаоса, были неблагоприятные экологические условия и деятельность человека, а также естественный распад древостоя. Некоторые древостои могут существовать в таком положении длительное время, иногда даже постоянно на протяжении жизни всего древостоя, например, дубняки порослевого происхождения (по причине повторяющихся пожаров) или дубняки, произрастающие на морских террасах, в экстремальных условиях.

Фото 1. Кедрово-лиственничное возобновление после выборочной рубки и пожара (пп 7-2000) в Сохондинском биосферном заповеднике
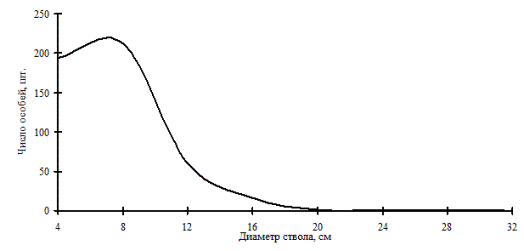
Рис. 1. Общее частотное распределение формирующегося древостоя в 2005 г. на месте сплошной вырубки (пп 7-2000) на территории Сохондинского биосферного заповедника |
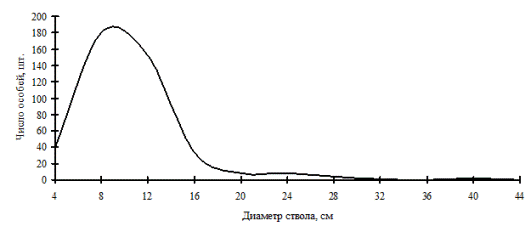
Рис. 2. Общее частотное распределение древостоя в дубняке рододендроновом (пп Б-4) в 1998 г. на территории Сихотэ-Алинского биосферного заповедника |
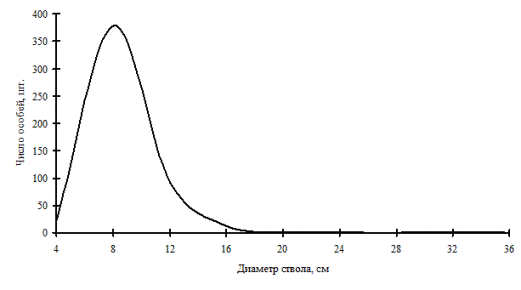
Рис. 3. Общее частотное распределение древостоя в дубовом криволесье (пп А-1) в 2000г. на территории Сихотэ-Алинского биосферного заповедника |
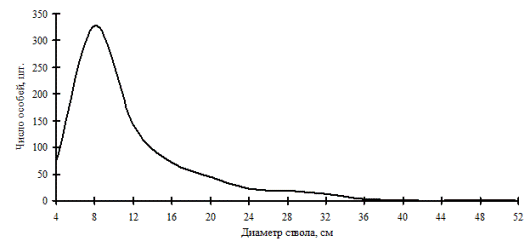
Рис. 4. Частотное распределение пихты белокорой в пихтовом лесу (пп Ф-2) на территории Сихотэ-Алинского биосферного заповедника в 2005г. |
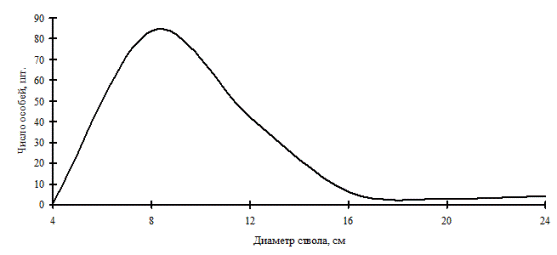
Рис. 5. Частотное распределение пихты белокорой в березняке (пп М-1) на территории Сихотэ-Алинского биосферного заповедника в 1999г. |
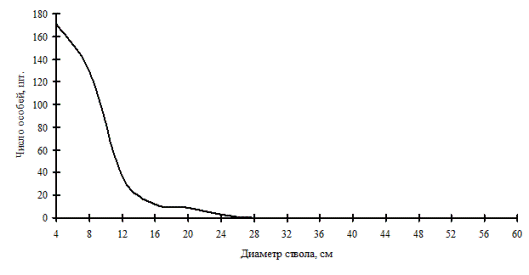
Рис. 6. Частотное распределение пихты белокорой в кедрово-пихтовом лесу (пп З-4) на территории Сихотэ-Алинского биосферного заповедника в 1998г. |
Нахождение в точке бифуркации дает шанс системе перейти к той или иной новой структуре. Один из вариантов перехода древостоев на другой уровень в строении по толщине - это переход к так называемому «нормальному распределению». Но переход системы к такому состоянию не выводит ее из состояния хаоса, в данном случае из состояния недискретности изменчивости древостоя по толщине ствола древесной популяции. По нашим данным, такую картину, близкую к «нормальному распределению» древостоя, мы наблюдали в дубняке, произрастающем на второй морской террасе Сихотэ-Алинского заповедника (рис. 7). Очень часто такая структура древостоя наблюдается в ценопопуляциях пионерных видов деревьев, образующих промежуточные сообщества в сукцессионных рядах. Примером этому могут служить ценопопуляции березы плосколистной, произрастающие на месте кедровых лесов, уничтоженных пожарами (рис. 8) и вырубками (рис. 9 и рис. 10) в Сихотэ-Алинском заповеднике. Такой тип строения древостоя наблюдается в сообществах, где деревья по мере их роста переходят в среднюю по толщине ценотическую группу, при этом присоединение новых элементов в виде тонкомера отсутствует. Это происходит за счет неблагоприятного влияния материнского (условно коренного) древостоя на подрост (как в случае с ценопопуляциями дуба и березы) и неблагоприятного влияние взрослых деревьев других видов. Такой тип неоднородности говорит о неперспективности изменчивости деревьев в ценопопуляции или о деградации либо всего древостоя, либо отдельных ценопопуляций видов, входящих в его состав (Галанин и др., 2000; Василенко, 2001; Галанин, Беликович, 2004).
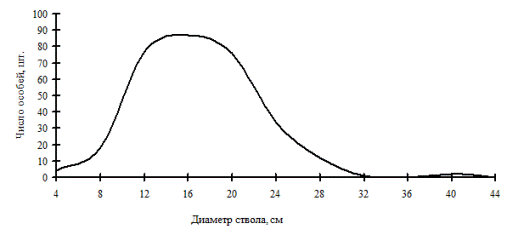
Рис. 7. Общее частотное распределение особей (дуб монгольский) в дубняке (пп Б-1) в 1998 г. на территории Сихотэ-Алинского биосферного заповедника |
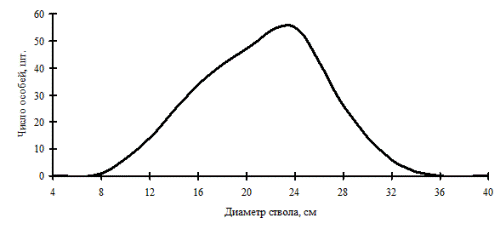
Рис. 8. Частотное распределение березы плосколистной в березняке (пп М-1) на территории Сихотэ-Алинского биосферного заповедника в 1987 г. |
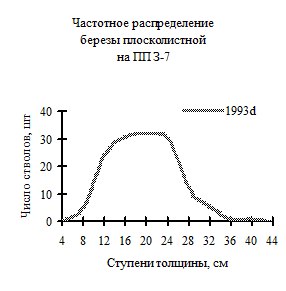
Рис. 9. Частотное распределение березы плосколистной в березняке (пп З-7) на территории Сихотэ-Алинского биосферного заповедника в 1993 г. |
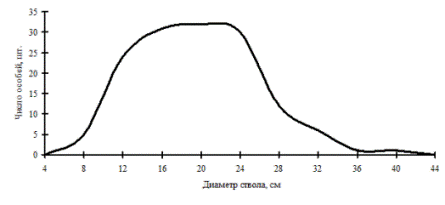
Рис. 10. Частотное распределение березы плосколистной в кедрово-пихтовом лесу (пп В-6) на территории Сихотэ-Алинского биосферного заповедника в 2000 г.
|
Рассмотренные выше примеры неоднородности в строении древостоев по диаметру характерны для систем с малым числом видов либо с малой долей участия произрастающих в фитоценозе видов. От этого также зависит разнообразие ценотических классов и, в соответствии с этим, перспективность данного фитоценоза, т.е. его устойчивость. В сообществах, где доминирует один вид, например, дуб монгольский в нижнем лесном поясе (рис. 11 и рис. 12) и пихта белокорая в поясе елово-пихтовых лесов (рис. 13) Сихотэ-Алинского заповедника, доминирующий вид полностью задает картину распределения деревьев по толщине, остальные породы деревьев присутствуют в древостое небольшим числом и представляют недискретные неоднородные совокупности деревьев по толщине.
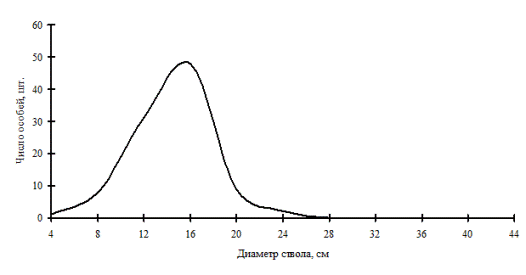
Рис. 11. Частотные распределения всего древостоя дубового леса (пп Б-3) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 1998 г. |
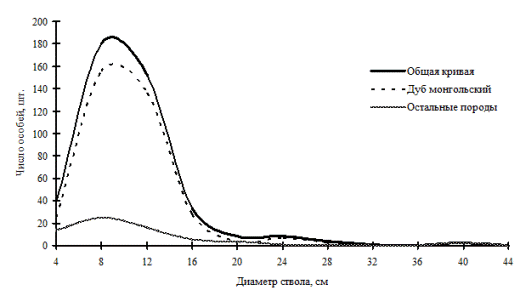
Рис. 12. Частотные распределения всех особей древостоя дубового криволесья (пп А-1) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 2000 г. |

Фото 2. Кедрово-пихтовый лес с рододендроном Фори в Сихотэ-Алинском заповеднике
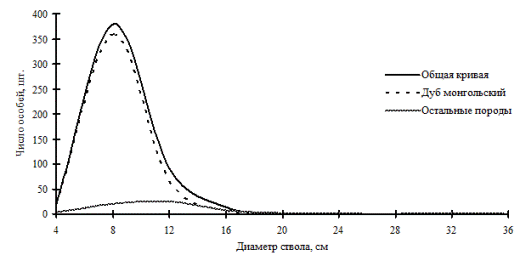
Рис. 13. Частотные распределения всего древостоя дубового леса (пп Б-2) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 2005г. |
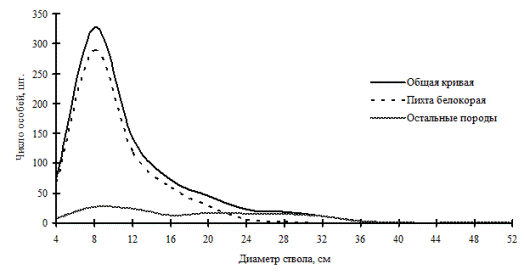
Рис. 13а. Частотные распределения всего древостоя пихтового леса (пп Ф-12) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 2005г. |

|
В монодоминантных лесных фитоценозах встречаются и древостои, неоднородные по толщине, в основном это древостои семенного происхождения. Примерами таких ценозов могут служить дубняки из дуба монгольского на территории Сихотэ-Алинского биосферного заповедника (рис. 14; фото 5) и из дуба зубчатого на территории Лазовского заповедника (рис. 15; фото 3). Эти примеры показывают, что в древостоях с доминированием одной древесной породы можно также наблюдать присутствие всех функциональных частей в виде разных дискретных групп деревьев по толщине. Но, к сожалению, дальнейшее поддержание и самоусложнение такого разнообразия внутренней структуры древостоев невозможно при отсутствии поступления новых элементов в виде подроста в ценопопуляции этих видов дуба.
Фото 3. Дубняк из дуба зубчатого (пп 2-2005) в Лазовском заповеднике |
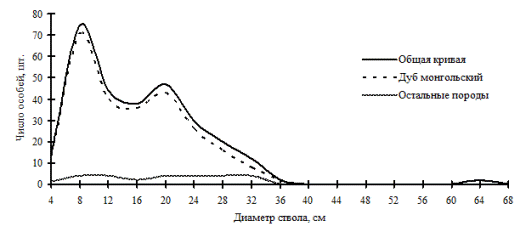
Рис. 14. Частотные распределения всего древостоя дубового леса (пп Б-5) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 1998 г. |
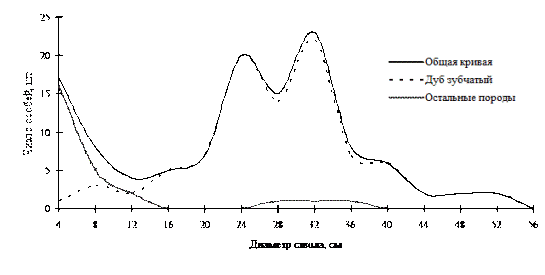
Рис. 15. Частотные распределения всего древостоя дубового леса (пп 2-2005) и отдельных видов, входящих в его состав, на территории Лазовского заповедника в 2005г. |
2.2. Социальная структура многокомпонентных и разновозрастных древостоев
В отличие от монодоминантных сообществ, растительные сообщества, состоящие из нескольких видов деревьев, имеют сложную внутреннюю структуру, в том числе по количеству и составу функциональных элементов. Чем больше видов представлено в древостое, тем сложнее его внутренняя структура и набор функциональных элементов (Высоцкий, 1962; Иванюта, 1964; Рыжков, 2000). Процесс дифференциации деревьев даже для одной и той же породы протекает по-разному в чистых и смешанных древостоях (Логвинов, 1971; Валяев, 1972; Чуенков, 1972). Как уже говорилось выше, многовидовые объединения особей более полно используют эдафические и аэрологические ресурсы местообитания, чем одновидовые. Многокомпонентность сообществ особенно важна в условиях суточных, сезонных и разногодичных колебаний погодных условий и в соответствии с ними колебаний величины термодинамического потенциала местообитаний (Дылис, Уткин, 1968; Галанин, 1989). Многокомпонентность лесных сообществ проявляется не только в совместном произрастании нескольких видов, но и в наличие различных ценотических групп деревьев по толщине. Чем ярче выражены такие временные элементы, тем в более равновесном состоянии находится система. Если принять во внимание тот факт, что каждый из этих элементов выполняет свою роль в биогеохимической структуре сообщества, можно говорить о более высокой ее организованности и эффективности за счет выполнения каждой группой деревьев своей функции в общей работе системы.
Для каждого древесного ценоза картина распределения особей деревьев по толщине ствола будет индивидуальной, но некоторые общие черты можно проследить. Общая картина распределения особей в древостоях, образованных несколькими видами деревьев, может резко отличаться от распределения отдельных ее частей. Неоднородные по толщине деревьев ценопопуляции могут образовывать более или менее непрерывный по однородности древостой, и наоборот, вся совокупность деревьев может быть неоднородна и дискретна.
Одной из общих тенденций в строении некоторых древостоев, состоящих из нескольких видов, является образование отдельными породами разных ценотических классов. Такую картину распределения мы наблюдали в лиственничнике бамбуковом (рис. 16), произрастающим на острове Сахалин, где лиственница была посажена на месте сплошных рубок. Ценопопуляция лиственницы имеет деформацию в среднем по толщине классе деревьев, в группе тонких деревьев деформация отсутствует. Как уже отмечалось выше, такое строение древостоя по толщине ствола деревьев говорит о неустойчивости ценопопуляции лиственницы в рассматриваемом сообществе.
стальные породы этого ценоза, составляющие класс угнетенных деревьев - это береза шерстистая, пихта сахалинская и сосна обыкновенная, при этом особи сосны были посажены в то же время, что и лиственницы, но не выдержали конкуренции с ними и с бамбуком курильским. Дальнейшая судьба этого сообщества может развиваться по нескольким сценариям, однако без вмешательства внешних факторов в любом случае на смену лиственнице придут такие виды, как пихта и ель.
Еще одним примером аналогичного типа строения древостоя по толщине стволов является чозениевый лес, расположенный на первой надпойменной террасе в Сихотэ-Алинском биосферном заповеднике (рис. 17; фото 4). Разделяя общую совокупность деревьев на ценопопуляции, мы видим, что группу индетерминантных деревьев и группу лидеров здесь составляют особи одного вида - чозении крупночешучайтой, в подросте и возобновлении данный вид отсутствует. Группа угнетенных особей состоит из тополя Максимовича, ясеня маньчжурского, ольхи волосистой и других пород. Участвующие в примеси породы в дальнейшем могут придти на смену доминирующей ценопопуляции чозении, это может произойти в случае повышения уровня террасы в результате вреза русла реки. Если река замоет свое сегодняшнее русло, то на повторно сформированной пойменной террасе вновь может образоваться чозениевый лес. В связи с перемещением русла в поймах рек этот процесс повторяется тысячи и десятки тысяч раз (Галанин и др., 2000).
Березово-лиственничный лес, произрастающий на территории Сихотэ-Алинского биосферного заповедника, образовавшийся в ходе послепожарной сукцессии, можно тоже отнести к этому типу по строению древостоя (рис. 18; фото 6). Береза плосколистная здесь относится к неустойчивым ценопопуляциям, и распределение ее особей по толщине носит характер «нормального распределения». Она уже потеряла ведущую позицию в данном фитоценозе, и эта роль теперь принадлежит лиственнице, которая уже имеет две ценотические группы деревьев – группу угнетенных и группу индетерминантных особей.
Рассмотренные нами типы строения древостоев по толщине ствола находятся в неустойчивом положении, а точнее, в состоянии хаоса:в каждом из этих древесных ценозов доминирующий вид теряет свои позиции лидера. Главная причина – это внутренний фактор, отсутствие группы тонких деревьев в каждой из рассмотренных ценопопуляций. Без присоединения к ценотической системе новых элементов (в данном случае это особи подроста, пополняющие в процессе динамики группу тонких деревьев) ценопопуляция теряет ведущую позицию, а древостой меняет своего лидера.
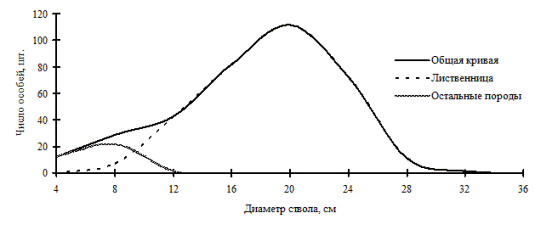
Рис. 16. Частотные распределения всего древостоя лиственничного леса (пп 1-2005) и отдельных видов, входящих в его состав, на острове Сахалин в 2005 г. |
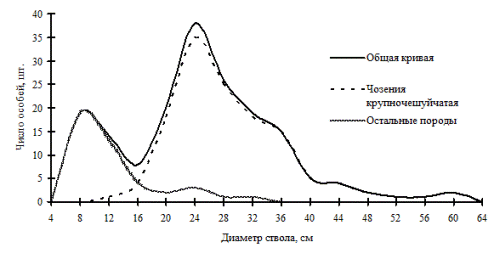
Рис. 17. Частотные распределения всего древостоя чозениевого леса (пп З-8) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 1999 г. |

Фото 4. Чозениевый высокотравно-кустарниковый лес (пп З-8) в Сихотэ-Алинском биосферном заповеднике
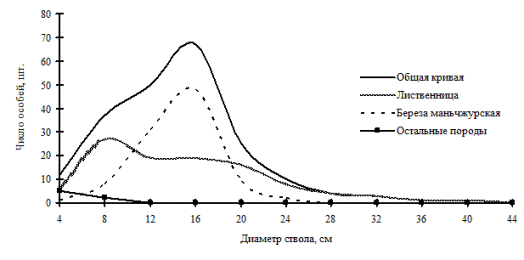
Рис. 18. Частотные распределения всего древостоя березово-лиственничного леса (пп В-4) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 2000 г. |
Состояние хаоса в многокомпонентных ценозах может проявляться в различной степени, но, как и в одновидовых ценозах, причины потери системой равновесия – это либо естественный распад основного яруса древостоя, либо воздействие антропогенных факторов. В таком состоянии система становится открытой и дает шанс другим видам занять лидирующее положение. Однако не все виды, входящие в состав древостоя, имеют шансы занять лидирующее положение в такой ситуации (Сукачев, 1950; Нечаева, 1979; Галанин, 1989).
Состояние хаоса в многовидовых древостоях, возникшее по причине отсутствия тонкомерной части ценопопуляции лидера, можно проиллюстрировать на примере двух ценопопуляций дуба монгольского в кедрово-дубовых лесах, расположенных в Сихотэ-Алинском заповеднике (рис. 19 и рис. 20). Здесь ведущие позиции, которые некогда принадлежали дубу монгольскому, занимают более молодые ценопопуляции кедра корейского. |

Фото 5. Дубняк разнотравно-осочковый (пп Б-12) в Сихотэ-Алинском биосферном заповеднике |

Фото 6. Березово-лиственничный лес (пп В-4) в Сихотэ-Алинском биосферном заповеднике |
Обе ценопопуляции кедра являются прогрессирующими, и за счет наличия всех ценотических групп они обеспечивают себе устойчивую позицию, в отличие от дуба монгольского, ценопопуляции которого представлены группой индетерминантных особей и особей лидеров. В результате развития растительности в этих сообществах происходит усложнение общей ценотической структуры древостоя и повышение эколого-ценотического разнообразия. Дуб на данных участках леса постепенно переходит под полог кедра, и в дальнейшем должен довольно быстро выпасть из состава древостоя. Прервать этот естественный процесс может только лесной пожар. Тогда восстановление леса снова начнется с дубово-березовой стадии.
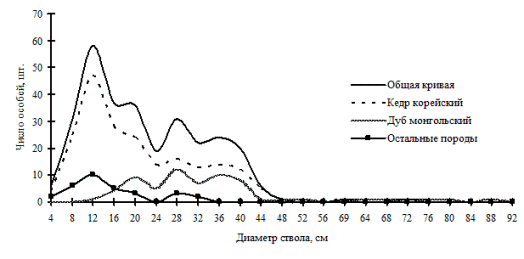
Рис. 19. Частотные распределения всего древостоя кедрово-дубового леса (пп З-1) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 1998 г. |
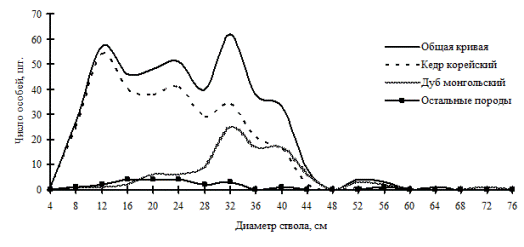
Рис. 20. Частотные распределения всего древостоя кедрово-дубового леса (пп З-2) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 1998 г. |

Фото 7. Леса в заповеднике "Бастак" в Еврейской Автономной области. Фото с сайта: http://www.vitim.zapoved.ru/ca… |
Еще большую степень хаоса можно наблюдать в системах, нарушенных человеком, об этом свидетельствует увеличение численности группы тонкомера. Резкое увеличение численности тонкомера, превышающее допустимое экотопом данного местообитания, может служить показателем неравновесности системы в целом. При этом в результате антропогенного фактора происходит возрастание не только числа элементов, которые могут заменить основного лидера, но и пород, сопутствующих основному эдификатору. По нашим данным, такая картина наблюдается в древостоях, где были проведены частичные вырубки, в кедрово-пихтовом лесу в Сихотэ-Алинском заповеднике (рис. 21) и в елово-кедровом лесу в заповеднике «Бастак» (рис. 22; фото 7).
Таким образом, для достижения древостоем либо ценопопуляцией лидером динамического равновесия необходимо присутствие всех ценотических групп деревьев и, что особенно важно для поддержания такого устойчивого состояния, постоянное пополнение группы тонких деревьев из подроста. Как правило, вид-эдификатор в фитоценозах представлен серией индивидов, отличающихся друг от друга как по возрасту, так и по мощности своего развития. Только при полном разнообразии возрастной структуры и структуры по мощности особей древостоя, эдификатор обеспечивает себе господствующее положение в фитоценозе и устойчивость при воздействии самых разных неблагоприятных факторов (Шенников, 1944; Бузыкин, Гавриков и др., 1985). |
Эндогенное развитие ценопопуляций деревьев направлено в сторону абсолютной разновозрастности, в которой они пребывают при отсутствии нарушений фитоценоза неопределенно долгое время. В этом проявляется системное качество популяции – ее непрерывность, порожденная сменой поколений (Миркин, Розенберг, 1978; Куркин, Матвеев, 1982).
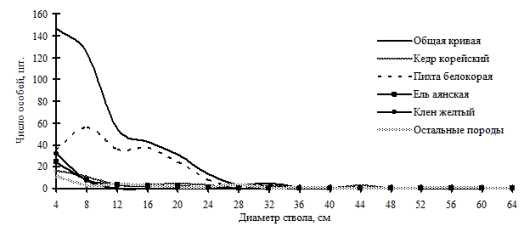
Рис. 21. Частотные распределения всего древостоя кедрово-пихтового леса (пп В-6) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 2000 г. |
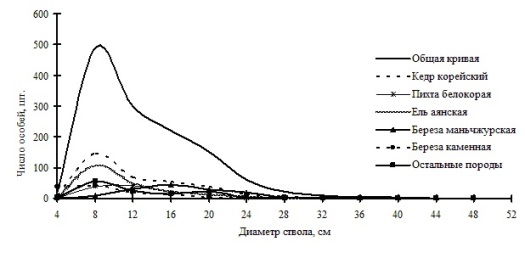
Рис. 22. Частотные распределения всего древостоя елово-кедрового леса (04-2002) и отдельных видов, входящих в его состав, на территории заповедника «Бастак» в 2002 г. |
Примеры древостоев, находящихся в состоянии динамического равновесия, можно привести из разных заповедников. Интересную картину в распределении деревьев по толщине можно наблюдать в приручьевом кедрово-лиственничном (рис. 23; фото 7) и лиственнично-кедровом лесах (рис. 24; фото 8, 9) в Сохондинском биосферном заповеднике (здесь приводятся результаты ревизии за 5-летний период). Результаты первого описания строения этих древостоев по диаметру опубликованы в монографии А.В. Галанина и А.В. Беликович (2004), и хотя отнесение деревьев по ступеням толщины не соответствует стандартным лесоведческим, которыми мы пользовались в этой работе, можно проследить нечто общее с нашими данными. Общие черты заключаются в полном или частичном несовпадении деформаций на кривых частотных распределений двух ценопопуляций в рассматриваемых сообществах. Такое несовпадение авторы связывают с наличием своеобразного «кедрово-лиственничного эколого-ценотического маятника», возникающего в связи с несовпадением во времени и цикличностью активизации возобновления кедра и лиственницы в фитоценозе. Деформации кривых частотных распределений особей этих видов появились за счет конкурентного исключения и расхождения особей по ярусам, а также за счет неравномерного во времени процесса возобновления этих пород деревьев (Галанина, Беликович, 2004). Доля участия каждой ценопопуляции в этих смешанных древостоях практически одинакова и, в отличие от предыдущих ценозов, этот можно отнести к более равновесным системам.
Подобную картину можно наблюдать и в хвойно-широколиственном лесу, произрастающем на территории заповедника «Бастак» (рис. 25): здесь модальные значения численности для групп угнетенных, индетерминантных особей и особей лидеров для ценопопуляций пихты белокорой и кедра корейского не совпадают. По-видимому, это также можно связать с неравномерностью пополнения групп угнетенных деревьев в обоих ценопопуляциях.

Фото 8. Кедрово-лиственничный лес (пп 5-2000) в Сохондинском биосферном заповеднике
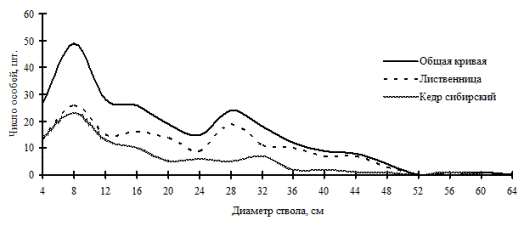
Рис. 23. Частотные распределения всего древостоя кедрово-лиственничного леса (пп 5-2000) и отдельных видов, входящих в его состав, на территории Сохондинского биосферного заповедника в 2005 г. |
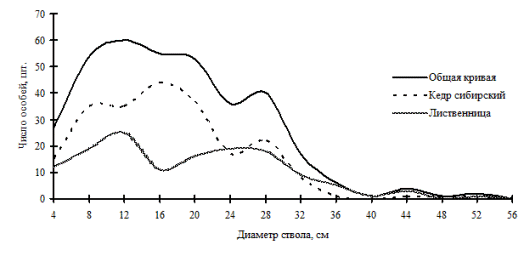
Рис. 24. Частотные распределения всего древостоя лиственнично-кедрового леса (пп 6-2000) и отдельных видов, входящих в его состав, на территории Сохондинского биосферного заповедника в 2005 г. |

Фото 9. Лиственнично-кедровый лес (пп 6-2000) в Сохондинском биосферном заповеднике
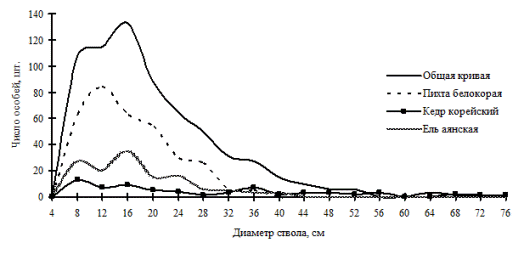
Рис. 25. Частотные распределения всего древостоя хвойно-широколиственного леса (05-2002) и отдельных видов, входящих в его состав, на территории заповедника «Бастак» в 2002 г. |
Другими примерами древесных ценозов, находящихся в состоянии динамического равновесия, но в отличие от предыдущих примеров характеризующихся совпадением модальных значений основных групп деревьев по толщине, являются кедрово-лиственничный (рис. 26); кедрово-пихтовый (рис. 27) и сложный многоярусный хвойно-широколиственный лес (рис. 28) в Сихотэ-Алинском биосферном заповеднике, а также кедровник с елью и липой (рис. 29) и пихтово-кедровый лес в заповеднике «Бастак» (рис. 30; фото 10). В сложных многовидовых древостоях, кроме основных доминирующих видов, произрастают виды сопутствующих пород, которые никогда не выходят в доминанты и представлены в основном деревьями нижних ярусов. Способны ли такие виды входить в ценотическую группу лидеров или, образуя нижние ярусы древесного полога, эти деревья и по толщине составляют группу подчиненных? По нашим данным, такие виды в древостоях действительно имеются и входят в основном в группу угнетенных и индетерминантных деревьев, в группе лидеров такие виды представлены единичными особями. |
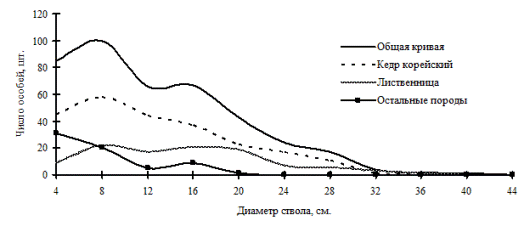
Рис. 26. Частотные распределения всего древостоя кедрово-лиственничного леса (пп З-3) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 1998 г. |
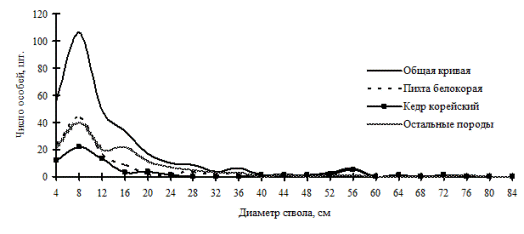
Рис. 27. Частотные распределения всего древостоя хвойно-широколиственного леса (пп З-6) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 1998 г. |
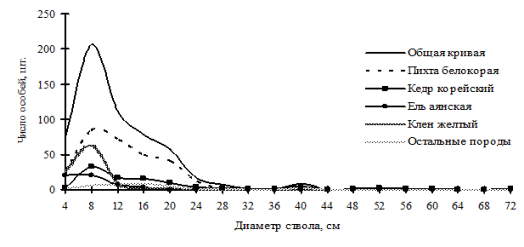
Рис. 28. Частотные распределения всего древостоя кедрово-пихтового леса (пп В-1) и отдельных видов, входящих в его состав, на территории Сихотэ-Алинского биосферного заповедника в 2000 г. |
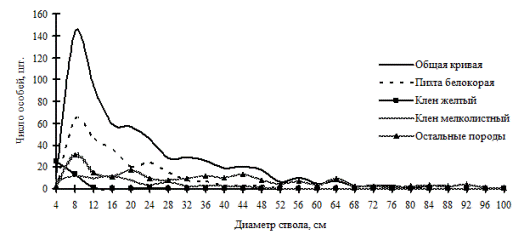
Рис. 29. Частотные распределения всего древостоя кедрового леса (01-2002) и отдельных видов, входящих в его состав, на территории заповедника «Бастак» в 2002г.
|
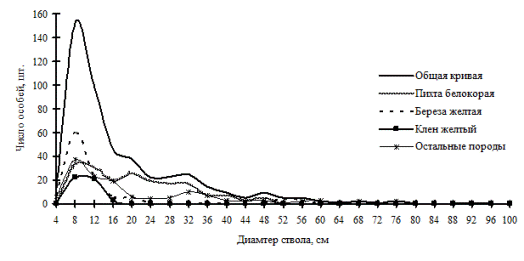
Рис. 30. Частотные распределения всего древостоя пихтово-кедрового леса (пп 02-2002) и отдельных видов, входящих в его состав, на территории заповедника «Бастак» в 2002г. |

Фото 10. Смешанные пихтово-кедровые леса на территории заповедника "Бастак". Фото с сайта: http://tigromania.ru/news/zapo… |
Продолжение следует
