Мониторинг и биоразнообразие экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2012. – 140 с.
Список авторов публикаций. С. 139. Предисловие C.6-8.
Галанин А.В., Галанина И.А. Динамическая парадигма в геоботанике и ботанической географии. С. 9-24.
Урусов В.М., Петропавловский Б.С., Варченко Л.И. К корректировке ботанико-географического и флористического районирования Дальнего Востока России С. 25-45.
Сухомлинов Н.Р. Мониторинг пирогенной трансформации экосистем: проблемы, методы, подходы. С. 46-52.
Игуменова О.П. Экологический мониторинг юго-западной территории республики Башкортостан. С. 53-60.
Киреева Н.А., Якупова А.Б., Григориади А.С. Интегральная оценка устойчивости почвы к нефтяному загрязнению. С. 61-66.
Сухомлинова В.В. Изменение межвидовых стратегических взаимодействий при пирогенной трансформации экосистем. С. 67-74.
Галанин А.В., Долгалева Л.М. К вопросу о применении методики ранжированных невзвешенных вариационных рядов для анализа древостоев на постоянных пробных площадях в Сохондинском биосферном заповеднике. С. 75-92.
Галанин А.В., Галанина И.А. Корейско-Хасанская ботанико-географическая подобласть (очерк растительности). С. 93-114.
Миронова Л.Н., Реут А.А. Итоги репродукции редких и исчезающих видов декоративных травянистых растений в республике Башкортостан. С. 114-118.
Энхтайван Н. Устойчивое лесоуправление в северной Монголии. С. 119-123.
Урусов В.Н., Варченко Л.И. Врищ Д.Л., Петропавловский Б.С. Пути улучшения озеленения Владивостока. С. 123-138.
|

|
УДК 58.00 ББК 72 я 43 М77
Рецензент Христофорова Н.К., д.б.н., проф. ДВФУ
Мониторинг и биоразнообразие экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2012. – 140 с. ISBN 978-5-89694-136-1
Редакционная коллегия: Долгалева Л.М., к.б.н.; Беликович А.В., д.б.н.; Галанин А.В., д.б.н., проф; Петропавловский Б.С., д.б.н.
В сборнике представлены результаты научно-исследовательской и практической работы ученых, преподавателей, аспирантов Дальневосточного региона, в которых отражены глобальные изменения климата и трансформация экосистем, флора, биология и экология растений Сибири и Дальнего Востока, экология природных сообществ, информационные технологии и системы в исследовании экосистем, экологические проблемы сохранения биоразнообразия природных систем Сибири и Дальнего Востока, проблемы охраны и рационального использования животного мира Сибири и Дальнего Востока.
Предназначен для студентов, аспирантов, ученых, специалистов и профессорско-преподавательского состава колледжей и университетов.
© НОУ ВПО «Институт технологии и бизнеса», 2012
|
УДК 574.42
К ВОПРОСУ О ПРИМЕНЕНИИ МЕТОДИКИ РАНЖИРОВАННЫХ НЕВЗВЕШЕННЫХ ВАРИАЦИОННЫХ РЯДОВ ДЛЯ АНАЛИЗА ДРЕВОСТОЕВ НА ПОСТОЯННЫХ ПРОБНЫХ ПЛОЩАДЯХ В СОХОНДИНСКОМ БИОСФЕРНОМ ЗАПОВЕДНИКЕ
А.В. Галанин, Л.М. Долгалева
А.V. Galanin, L.М. Dolgalyeva. ON THE QUESTION OF APPLICATION OF THE METHOD OF RANGED UNWEIGHTED VARIATIONAL SERIES TO ANALYSE FOREST STANDS ON CONSTANT SAMPLE PLOTS IN SOKHONDO BIOSPHERE RESERVE.
The paper includes results of forest stand analysis on constant sample plots selected in the southern Transbaikalia by the method of variational series. The authors proved efficiency of the method in monitoring vegetation.Кeywords: geobotany, silvics, research methods, forest stand, vegetation monitoring.
Большинство геоботаников использует понятие о площади выявления ассоциации (Ярошенко, 1953; Марков, 1962; Сукачев, 1961, 1972; Воронов, 1973; Анучин, 1982). При этом достаточно большой по площади фитоценоз рассматривается как выразитель основных признаков ассоциации. Однако если рассматривать растительную ассоциацию как совокупность не совсем одинаковых фитоценозов, то при характеристике и описании растительного покрова правильнее говорить еще и о площади выявления фитоценоза (Раменский, 1925; Быков, 1953; Шенников, 1964; Грейг-Смит, 1967; Василевич 1969). Для характеристики ассоциации таким образом необходима достаточно большая выборка пробных площадей из разных фитоценозов этой ассоциации, а размер каждой пробной площади должен быть не меньше площади выявления фитоценоза (Василевич, 1969).




|
Совокупность деревьев на пробной площади, заложенной в некотором массиве леса, может рассматриваться как репрезентативная выборка особей из ценопопуляций видов, растущих в данном сообществе (фитоценозе) в том случае, если в ней представлены все возрастные группы и все ярусы древостоя. Вопрос о размерах пробных площадей обсуждался в геоботанике и лесоведении более 100 лет, были проведены исследования по определению оптимального размера и конфигурации пробных площадей.
Установлено, что размер пробной площади зависит от типа растительности. Для луговой и степной растительности достаточна площадка 10х10 м, для лесной – 100х100 м, или, в крайнем случае, 50х50 м, для характеристики напочвенного покрова в лесу используют площадки 2х2 м и даже 0,5х0,5 м, а лишайниковую синузию можно изучать и с помощью площадок размером 15х20 см (Галанина,2008).
При определении размеров площади, с помощью которой мы будем получать характеристику ценопопуляции, необходимо в первую очередь учитывать принцип репрезентативности – соответствия структуры выборки структуре генеральной совокупности, которую с помощью выборки мы изучаем.
При этом одно из правил математической статистики гласит, что даже для характеристики однородной генеральной совокупности элементов объем выборки не должен быть меньше 30. Если же эта совокупность неоднородна, то объем выборки должен быть таков, чтобы каждый вариант (класс элементов) генеральной совокупности в среднем был представлен в выборке не менее чем 10 элементами.
При анализе размерной структуры древостоев (ценопопуляций древесных растений) чаще всего используют частотные распределения деревьев в зависимости от толщины ствола (Ипатов, 1969). Такие анализы можно встретить в большинстве публикаций по лесоведению. С их помощью были изучены многие аспекты структуры и динамики лесов в ходе сукцессий, эти методы в той или иной модификации известны хорошо геоботаникам и лесоведам (Галанин, Беликович, 2004; Василенко, 2008).
Мы же в этой статье рассмотрим метод статистического анализа, который пока не нашел широкого применения, и мало известен специалистам, так как он, хотя и очень прост, но не традиционен. Использовать этот метод до создания персональных ЭВМ было невозможно из-за его чрезмерной трудоемкости, но в настоящее время с использованием программы EXEL анализ этим методом занимает совсем немного времени. Этот метод мы называем методом невзвешенных ранжированных вариационных рядов. Впервые для анализа ценопопуляций лишайников эпилитов его применил А.А. Галанин (1995).
Понятие вариации – это ключевое понятие математической статистики, отражающее различие значений признака внутри исследуемой совокупности. Вариационный ряд – это ряд значений (характеристик) параметра на множестве элементов выборки из генеральной совокупности. В геоботанике часто используют ранжированный взвешенный вариационный ряд, или гистограмму, построенную на основе сгруппированных в классы измерений диаметра ствола, размера годового кольца, прироста побега и т.д. Каждая группа чисел (вариант), на которые разбивается совокупность, или выборка, называется классом. В лесной таксации такие классы называют ступенями толщины. В некоторых случаях совокупность градируется на классы по качественным признакам, имеющим неопределенные количественные характеристики.
В основу моделирования нами положен вариационный ряд окружности стволов деревьев на уровне 1,3 м, упорядоченный по возрастанию, его называют ранжированным невзвешенным вариационным рядом (Общая теория статистики, 1995). Эти параметры скоррелированны с ярусом, в котором растут особи в ценопопуляции и ролью дерева в ассимиляционных процессах фитоценоза. Толщина дерева (размер окружности ствола) – косвенная характеристика его доминирования в фитоценозе. Основными количественными характеристиками таких вариационных рядов являются средняя арифметическая, среднее квадратическое отклонение, дисперсия, размах вариации.
Если невзвешенный ранжированный вариационный ряд аппроксимировать непрерывной кривой, то можно получить некоторую модель генеральной совокупности, если только наша выборка была репрезентативна (А.А. Галанин, 1995, 2002). Длина вариационного ряда, степень его кривизны или вогнутости, разница между максимальными и минимальными значениями измеренного параметра несут информацию о генеральной совокупности.
Целью данной статьи является демонстрация результатов использования метода ранжированных невзвешенных вариационных рядов для анализа структуры древостоев.
Объекты и методы исследования
Объектом исследования послужили постоянные пробные площади (ПП), заложенные в типичных участках лесной растительности в двух высотных поясах в Сохондинском государственном биосферном заповеднике и в окрестностях с. Кыра Кыринского района Забайкальского края в период с 2004 по 2007 гг. Краткая характеристика ПП представлена в таблице 1.
Закладка и описания ПП проводилось по стандартной методике (Галанин, Беликович, 2004). Расположение их в Сохондинском заповеднике отмечено на рисунке 1.
Мониторинг растительного покрова в заповеднике и в пограничных территориях представляет собой комплексную систему наблюдений, оценки и прогноза изменений среды, структуры и биоразнообразия экосистем, позволяющую оценить изменения их состояния и происходящие процессы в естественных условиях и под влиянием антропогенной деятельности.
Мониторинг растительного покрова осуществляется для решения двух задач: анализа стабильности условий обитания и целостности экосистем особо охраняемой природной территории и для оценки скорости и характера динамики растительного покрова, для разработки прогнозов, рекомендаций и выбора вариантов управленческих решений. Поскольку сообщества растений лежат в основе круговорота вещества экосистем, то ключевым компонентом мониторинга окружающей среды является мониторинг состояния фитоценозов.
Каждые 5–10 лет на ПП делается оценка состояния компонентов фитоценоза и прогноз изменений в ценотических компонентах, вызванных факторами естественного и антропогенного происхождения. Учитывая, что фитоценозы – многокомпонетные системы, их количественная оценка должна проводиться на основе математических моделей.
Математические модели – это неполное абстрактное выражение систем реального мира, отражающих ряд математических зависимостей и описывающих какие-либо физические или биологические функции, закономерности структуры и динамики этих систем. В нашем случае вариационные ряды деревьев на ПП являются графическими моделями ценопопуляций видов деревьев и достаточно полно отражают структуру древостоев.
|
Вариационная кривая (рис. 2-7) – это наглядное изображение совокупности (выборки) особей по длине окружности стволов, показывающая как диапазон вариации свойства, так и частоту отдельных вариант.
Нами предложено использовать метод невзвешенных вариационных рядов, представляющих кривые модификации признаков, подчиняющиеся логарифмическому закону. Предложенный метод основан как на сравнении вариационной кривой с апроксимирующей ее логарифмической кривой, так и на оценке основных статистических величин, характеризующих ценопопуляцию (выборку деревьев). Несмотря на известность этого метода, для оценки структуры лесных фитоценозов другими исследователями он пока использовался очень мало (Н.А. Василенко, Т.А. Кузнецова-Парилова, И.В. Козырь). К достоинствам метода относится возможность работы с малыми выборками, наглядность, сравнимость по количественным характеристикам, достоверность, простота.
В левой части наших графиков-диаграмм показаны наиболее толстые особи, в правой части – более тонкие, в конце - тонкомер, подрост. Для характеристики популяции учитываются как длина вариационного ряда (от числа особей в ценозе зависит степень его развития), так и конфигурация кривой - ее отклонение от апроксимирующей логарифмической кривой.
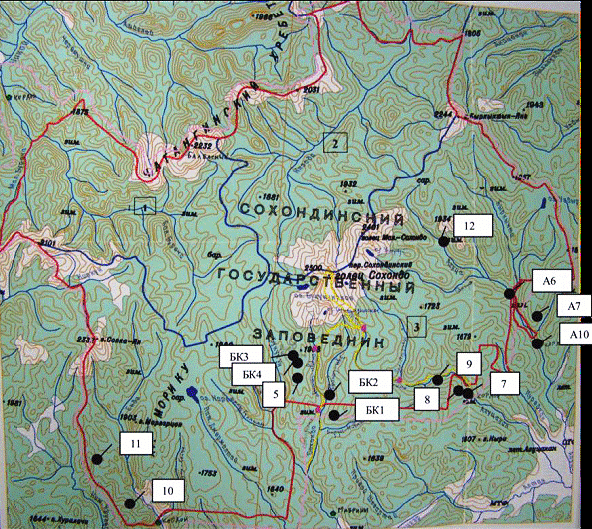
Рис. 1. Карта Сохондинского государственного заповедника и размещение постоянных лесных пробных площадей, заложенных авторами в 2000–2007 гг. |
Наиболее оптимальным, по мнению многих авторов, является выборки, насчитывающие не менее 200 деревьев на ПП, в нашем случае этому правилу обычно соответствует пробная площадь размером 0,25 га (Методы изучения лесных сообществ, 2002). Такая ПП позволяет получить достоверную
информацию, оцениваемую по количественным и качественным критериям.
В экологии используется широкий круг характеристик параметров, вычисленных в виде средних величин их значений. Вообще надо помнить, что вычисление среднего арифметического – это один из распространенных приемов обобщения материала. Средний показатель отражает общие, характерные признаки генеральной совокупности, игнорируя различия отдельных ее единиц, которые могут отклоняться от средней в ту, или иную сторону. По закону больших чисел случайности оклонений членов вариационного ряда от среднего уравновешиваются, поэтому и можно абстрагироваться от количественных значений отдельностей, ограничившись в ряде случаев только средними значениями признаков (параметров).
Динамика средних значений признаков генеральной совокупности во времени – это обобщающая характеристика процесса в тех конкретных условиях, в которых он протекает. При этом величины средней арифметической оцениваются совместно с показателями модального значения, среднеквадратического отклонения, коэффициента вариации. Таким образом статистическую выборку и генеральную совокупность характеризуют не только по средней величине признака, но и по степени вариабильности и степени асимметрии вариационного ряда.
Помимо этого статистические невзвешенные распределения в виде кривых можно сравнивать с их логарифмическими и иными моделями.
|
По форме мы различаем невзвешенные вариационные кривые вогнутые, выпрямленные, выпуклые, сложные. Первые часто характерны для устойчивых и агрессивных ценопопуляций. Вариационные ряды выпрямленные характерны для ценопопуляций, выходящих в 1 ярус и занимающих доминирующую позицию в фитоценозе, но динамическое равновесие которых находится под угрозой. Выпуклые вариационные кривые (как правило, в своей средней части) показывают деградацию ценопопуляции за счет сокращения количества возобновления и тонкомера и выпадения перестойных деревьев.
Более сложная форма кривых невзвешенных распределений показывает гетерогенность изучаемой ценопопуляции, ее непростые конкурентные отношения с другими деревьями фитоценоза - видами, возрастными группами. Ломанные линии вариационных рядов указывают на катастрофические изменения, повлиявшие на ценопопуляцию на одном из этапов ее развития. По длине вариационного ряда мы выделяем нормальные выборки (с размером выборки 100–200 особей), дефицитные (менее 100 особей) и избыточные (более 200–250 особей).
Многие процессы роста и развития живых систем подчиняются логарифмическому закону, когда некоторый измеряемый параметр изменяется со скоростью, пропорциональной имеющемуся в данный момент значению. Логарифмическая функция выражает то же самое, что и формула степенная. Логарифмическая функция называется обратной к степенной функции. Известно, что логарифмическая функция моделирует самые разные природные явления. По логарифмическим спиралям, например, выстраиваются цветки в соцветиях подсолнечника, закручиваются раковины моллюска Nautilus и рога горного барана. Все эти природные формы могут служить примерами кривой, известной под названием логарифмической спирали. Такую кривую описывает движущаяся точка, расстояние которой от начала движения растет в геометрической прогрессии, а угол, описываемый ее радиусом-вектором, – в арифметической.

Рис. 2 Вариационный ряд тополя душистого (живых) на ПП-1 в 2004 г. в пойме р. Кыра |
Впервые логарифмическую функцию анализа размерной структуры популяций накипных лишайников использовал А.А. Галанин (1995, 2002). Он показал, что весьма надежным индексом для определения возраста ценопопуляции может служить теоретический максимальный диаметр, пред-сказанный логарифмическим трендом, убывающего вариационного ряда,
представляющего выборку диаметров слоевищ особей лишайников.
Эмпирические вариационные ряды особей лишайников описываются уравнениями регрессии типа y = a ln x + b, где y – диаметр слоевища, х – его порядковый номер в вариационном ряду, a и b – коэффициенты.
Мы начали использовать этот метод для анализа ценопопуляций деревьев на пробных площадях в Сихотэ-Алинскогм заповеднике в 1999 г. (Галанин и др., 1999) и Сохондинском заповеднике в 2000 г. (Галанин, Беликович, 2004). Выяснилось, что логарифмические модели вариационных рядов особей деревьев в ценопопуляциях являются весьма информативными количественными характеристиками этих ценопопуляций, что очень важно при их длительном мониторинге.
Латинские названия растений в этой статье приводятся по «Флоре центральной Сибири» (1979). |
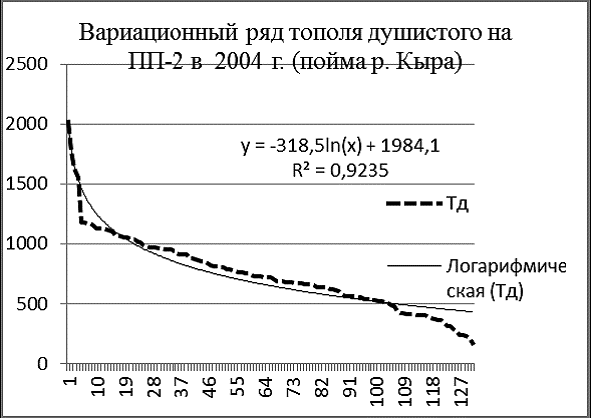
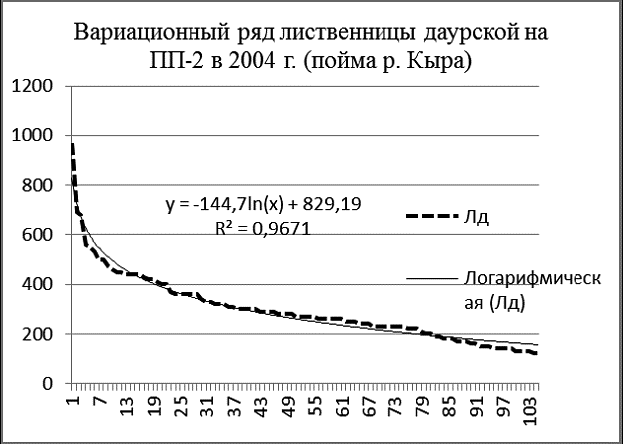
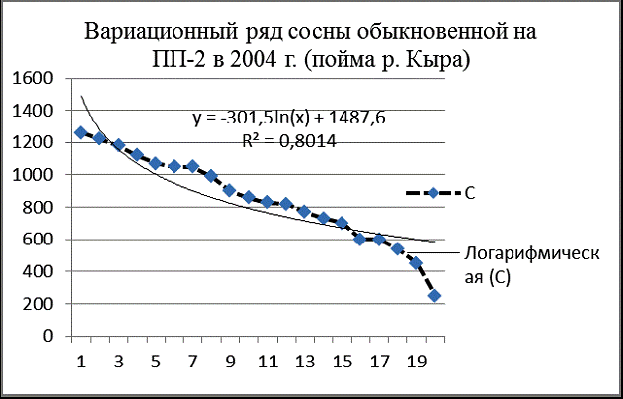
Рис. 3. Вариационные ряды деревьев и апроксимирующие их логарифмические кривые на ПП-2 в 2004 г. в пойме р. Кыра |
Результаты и их обсуждение
Ниже даны краткие геоботанические описания ПП и анализ ранжированных невзвешенных вариационных рядов деревьев на этих ПП.
На ПП-1 в пойме р. Кыра растет перестойный тополевый лес. Всего на площади зарегистрировано 62 особи тополя душистого (Populus suaveolens).
Вариационный ряд – расположение особей в порядке убывания их толщины – представлен на рис. 2. Дефицитная выборка говорит о деградации древостоя. Приспущенные ветви кривой указывают на дефицит как тонкомера, так и модельных деревьев. Флуктуации, графически отображенные на кривой как микроволны, показывают неустойчивость ценоза, конкурентную борьбу особей тополя за экологические ресурсы, которые часто оказываются в дефиците.
Этот ряд аппроксимируется волнистой линией, форма которой не вполне соответствует кривой логарифмического роста (сплошная тонкая линия). Отклонение от логарифмического ряда наблюдается в отрицательную сторону (-) в области толстых деревьев и в области тонкомера. Если исходить из логарифмического закона роста, то очевидно, что в данной ценопопуляции деревья не достигают предельной толщины, возможной для этого вида в оптимальных условиях.
Предельная длина окружности ствола, рассчитанная по закону логарифмического роста, здесь у тополя должна быть 2650 мм. Неблагоприятные для тополя экологические условия (низкий уровень грунтовых вод, остепнение поймы, повышение ее уровня за счет аллювия) ограничивали рост деревьев. Об этом свидетельствует и низкий бонитет древостоя.
Скорее всего, такие теоретически предсказанные на основе логарифмического закона особи в этой ценопопуляции были, но они уже выпали.
Недостаточное количество тонкомера, рассчитанного по модели логарифмического роста, говорит о том, что возобновление тополя в этом сообществе затруднено в связи с понижением уровня грунтовых вод и переходом террасы из пойменного режима в надпойменный. Отчасти это вызвано и антропогенным фактором (выпас скота, сбор сухостоя на дрова фермерами).
Повышено против логарифмической модели роста в данной
ценопопуляции количество особей средней толщины. Это следствие выпада толстых особей, недостаточное возобновление и недостаточного пополнения ценопопуляции тонкомером. Таким образом, на данной пробной площади происходит деградация ценопопуляции тополя. На месте тополевого леса постепенно формируется закустаренный ксероморфный разнотравный луг с элементами остепнения.
Провал кривой распределения тополя на ПП-2 (рис. 3) в области толстомера вызван недавним пожаром и рубкой сухостоя, а также внедрением в древостой лиственницы даурской и сосны обыкновенной. На площади ПП-2 зарегистирировано 131 особь тополя душистого, 105 особей лиственницы даурской и 20 – сосны обыкновенной (исключая сухостойные деревья).
Вариационные ряды лиственницы и тополя почти нормальные, сосны – дефицитный. Приспущенные ветви кривых указывают на недостаток подроста, вызванный как затенением, так и флювиальными процессами реки. Вариационный ряд тополя представляет собой ломаную кривую, которая резко отличается от его логарифмической модели. Похоже, что самые толстые деревья тополя вообще не относятся к данной ценопопуляции.
|
Они сохраняются здесь с того времени, когда пойма была занята чистым тополевником. Современная же ценопопуляция тополя на этой ПП находится в более пессимальных условиях, чем прежняя, и ее особи никогда не смогут достичь размеров немногих старых лидеров, выросших в прошлой экосистеме в более благоприятных для тополя условиях. Признаком деградации данной ценопопуляции является слабое возобновление тополя и ее пополнение.
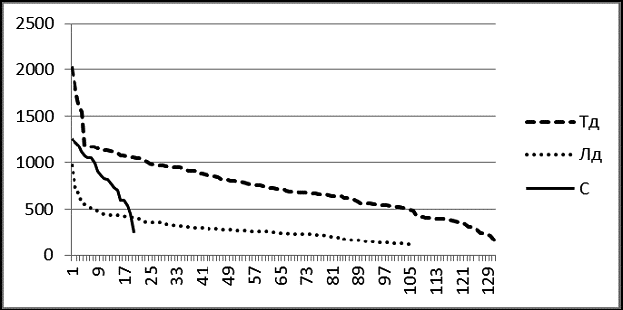
Рис. 4 Соотношение вариационных рядов живых тополя душистого (Тд), лиственницы даурской Лд), сосны обыкновенной (С) на ПП-2 в 2004 г. в пойме р.
Кыра |
Изображенные в одном масштабе на одной диаграмме вариационные ряды тополя, лиственницы и сосны на ПП-2 (рис. 4) наглядно характеризуют ценотическую роль этих видов в древостое в 2004 г. Лиственница даурская внедряется в фитоценоз, ее вариационный ряд вогнут и почти совпадает со своей логарифмической моделью. Вариационный ряд тополя говорит о размерной гетерогенности его ценопопуляции и о том, что по толщине и количеству особей тополь на ПП преобладает над лиственницей. Похоже, что пока тополь и лиственница в ценозе занимают разные экологические ниши, их распределения на диаграмме не пересекаются. Сосна по сравнению с лиственницей мало устойчива к пожарам, плохо растет на близко залегающей вечной мерзлоте, но она более вынослива к засухе. Сосна практически не встречается в сообществах поймы реки Кыры или присутствует там в качестве примеси. На диаграмме видно, что пополнение ценопопуляции сосны на ПП-2 затруднено, тонкомера явно недостаточно.
|
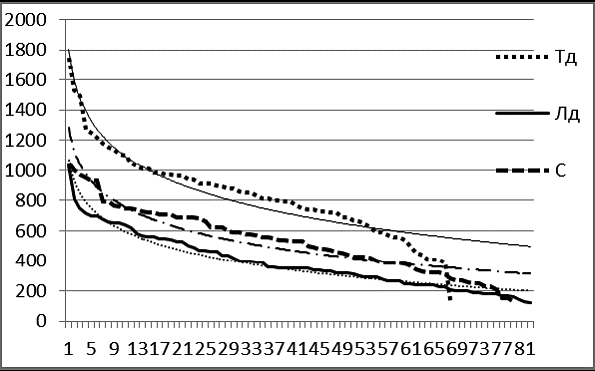
Рис. 5 Сводные вариационные ряды живых тополя душистого (Тд), лиственницы даурской (Лд), сосны обыкновенной (С) на ПП-3 в 2004 г. в пойме р. Кыра.
Тонкими линиями обозначены логарифмические кривые |
В сходном пойменном ценозе на ПП-3 (рис. 5) наблюдается начало деградации ценопопуляции тополя, о чем свидетельствует выпуклость вариационной кривой в области средних величин окружностей деревьев и резко спущенная правая ветвь кривой в области тонкомера.
Ценопопуляция сосны здесь также не в оптимуме. Сосна здесь не выдерживает конкуренции со стороны тополя и лиственницы. Ее особи не достигают максимальной теоретически расчитанной величины. Пополнение ценопопуляции из тонкомера затруднено, так как наблюдается его дефицит. Но ценопопуляция тополя на ПП-3 испытывает в большей степени дефицит поплнения, чем популяция сосны.
В пойме Кыры наносные плодородные почвы, высокая влажность почвы, аэрация благоприятны для тополя, который доживает здесь до 250 лет. Выход особей в лидеры у тополя, как и у лиственницы происходит здесь в соответствии с логарифмической моделью. Но особи лиственницы пока уступают тополю по толщине.
|
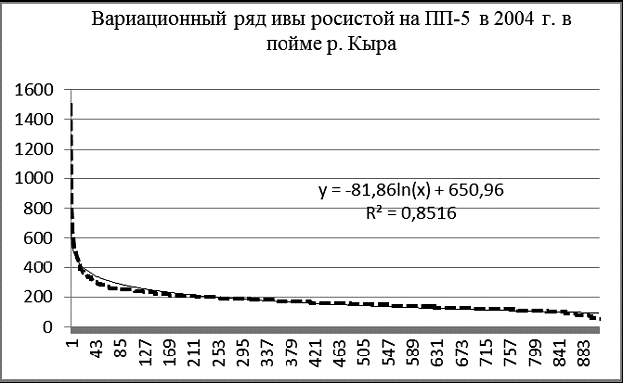
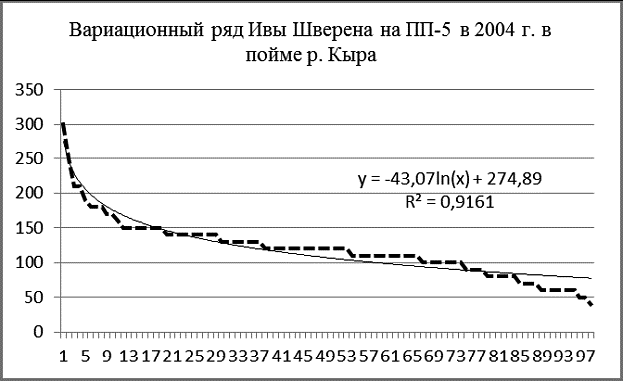
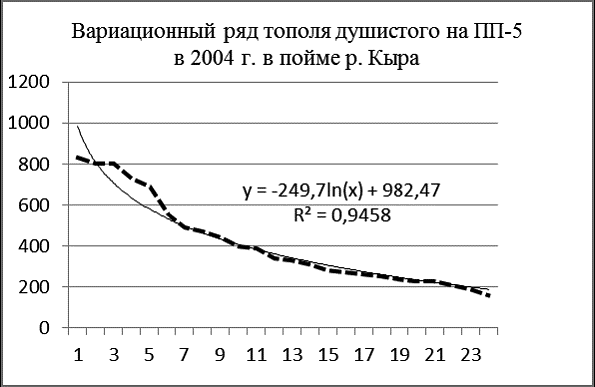
Рис. 6 Вариационные ряды живых ивы росистой (А), ивы Шверина (Б), тополя душистого (В) на ПП-5 в 2004 г. в пойме р. Кыра |
Между тем, высокая фаутность тополя (64%) говорит о уже идущих в этой экосистеме изменениях, вызванных понижением уровня грунтовых вод и снижением доступности растворенных солей корням деревьев тополя. Это и привело к сокращению подроста и пока что частичному усыханию деревьев первого яруса.
Судя по характеру вариационного ряда лиственницы, почти совпадающему со своей логарифмической моделью, можно утверждать, что будущее на ПП-3 за лиственницей. Фитоценотическая система на ПП-3 развивается в направлении выхода в абсолютные лидеры лиственницы при сохранении в качестве примеси сосны и полной деградации ценопопуляции тополя.
Идеальны по своим графическим параметрам древостои с высоким содержанием возобновления и подроста. К таким в пойме Кыры относятся ивовые ленточные леса, широко распространенные в лесо-степном поясе. На площади ПП-5 было зарегистрировано более 1100 особей двух видов ив и тополя душистого (рис. 6, 7). Здесь абсолютно преобладают ивы, численность тополя только 25 особей.
Судя по характеру вариационных рядов, численность особей ивы шверина и тополя душистого достигла предела, их возобновление затруднено. Ценопопуляция ивы росистой пока пополняется из подроста очень интенсивно.
В наибольшей степени логарифмической модели соответствует размерная структура ценопопуляции ивы росистой, в меньшей – ивы Шверина. Успешнее всех в межвидовой конкуренции ива росистая, которая по численности заметно превосходит иву Шверина – 889 особей против 97.
Ива Шверина (гигрофит) занимает экосистеме особую экологическую нишу, размещаясь по границам заплеска реки во время периодических паводков, поэтому она находится под контролем флювиальных процессов.
А вот ива росистая (гигромезофит) предпочитает террасы с умеренным увлажнением. На ПП-5 экологические условия больше соответствуют ей.
Тополь в таком фитоценозе находится в состоянии готовности к «ценопопуляционному рывку», его ценопопуляция накопила некоторое количество относительно толстых особей, но пока что они в конкурентной борьбе уступают ивам. Возможно в будущем ослабление пойменного режима ослабит позицию ив, и тогда тополь на площади ПП-5 сможет стать лидером.
Сводные вариационные ряды особей разных видов в фитоценозе показательны для оценки экологической роли каждого из них. Доминирование ивы росистой связано с тем, что она здесь в умеренно флювиальном местообитании находится в экологическом оптимуме для роста и возобновления. Роль тополя душистого на этом этапе развития экосистемы пока малозначительна. Ива Шверина занимаает в экосистеме свою экологическую нишу, ее вариационная кривая повторяет кривую ивы росистой, но на меньшем размерном уровне особей и при их меньшей численности.
|
Заключение
Рассмотренные нами примеры анализа размерной структуры древостоев на нескольких ПП с помощью ранжированных невзвешенных вариационных рядов позволяют
графически выявить тенденции изменений в ценопопуляциях видов деревьев. Метод вариационных рядов при анализе размерной структуры ценопопуляций деревьев
на постоянных пробных площадях позволяет получить информацию, необходимую для уточнения фитоценотического статуса ценопопуляций разных видов и
определения вектора динамики фитоценозов. Метод этот достаточно прост, благодаря использованию пакета программ Exel. Его использование возможно как при
очень большой численности особей на пробной площади, так и при их малой численности порядка 30 особей на ПП.
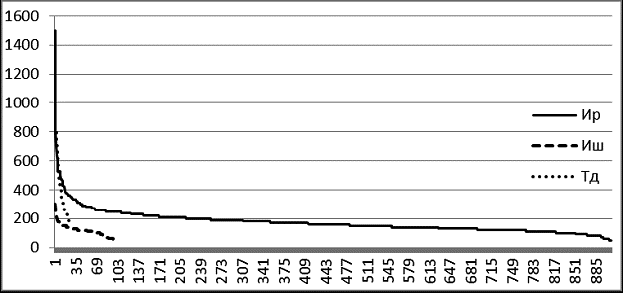
Рис. 7 Сводные вариационные ряды живых тополя душистого (Тд), ивы росистой (Ир), Ивы Шверина (Иш) на ПП5 в 2004 г. в пойме р. Кыра
|
Аппроксимация частотных распределений видов деревьев на пробных площадях в зависимости от их толщины логарифмическими линиями, по сути, является построением логарифмических моделей размерной структуры этих ценопопуляций. Логарифмическое уравнение y = - a ln(x) + b описывает идеальную равновесную структуру ценопопуляции. Здесь коэффициент b – это максимальный
размер особи в оптимальной ценопопуляции, a – напряженность конкуренции за ресурсы местообитания так, что чем больше а, тем интенсивнее конкуренция, тем сложнее особи данной ценопопуляции в конкретном фитоценозе выйти в лидеры, тем дольше ее особи задерживаются в состоянии тонкомера и в классе особей средней толщины.
Индех R2 показывает, насколько реальный вариационный ряд отклоняется от своей логарифмической модели, чем меньше значение этого индекса, тем больше размерная структура ценопопуляции соответствует этой модели.
|
Литература
1. Анучин Н.П. Лесная таксация. 5-е изд. – М.: Лесн. пром-сть, 1982. – 552 с.
2. Быков Б.А. Геоботаника. – Алма-Ата: Изд-во АН КазССР, 1953. – 457 с.
3. Василевич В.И. Статистические методы в геоботанике. – Л.: Наука, 1969. – 273 с.
4. Василенко Н.А. Самоорганизация древесных ценозов. – Владивосток: Дальнаука, 2008. – 171 с.
5. Воронов А.Г. Геоботаника. – М: Высшая школа, 1973. – 384 с.
6. Галанин А.А. Лихенометрические исследования на Северо-Востоке России. – Магадан: Изд-во СВНЦ ДВО РАН, 1995. – 51 с.
7. Галанин А.А. Лихенометрия: современное состояние и направления развития метода. – Магадан: Изд-во СВКНИИ ДВО РАН, 2002. – 74 с.
8. Галанин А.В., Беликович А.В. Постоянные пробные площади Сохондинского биосферного заповедника. – Чита: БСИ ДВО РАН, 2004. – 228 с.
9. Галанин А.В., Н.А. Василенко, А.А. Кузьмин. Эколого-ценотическое разнообразие хвойно-широколиственных лесов восточного макросклона Сихотэ-Алинского заповедника: оценка и анализ // Биологические исследования на Горнотаежной станции. – Владивосток: ДВО РАН, 1999. – С. 20–24.
10. Галанина, И.А. Синузии эпифитных лишайников в дубовых лесах юга Приморского края. – Владивосток: Дальнаука, 2008. – 238 с.
11. Грейг-Смит П. Количественная экология растений. – М.: Мир, 1967. – 358 с.
12. Ипатов В.С. Дифференциация древостоя. 2. Выявление деформаций кривых распределения деревьев по толщине // Вестн. ЛГУ. Биология. 1969. – №15. – вып. 3. – С. 43-53.
13. Марков М.В. О взаимоотношениях между растениями в растительных сообществах // Проблемы внутривидовых отношений организмов. – Томск: Изд-во Том. гос. ун-та, 1962. – С. 59–61
14. Общая теория статистики: учебник/ А.И. Харламов, О.Э Баширина, В.Т. Бабурин и др. – М.: Финансы и статистика, 1995. – 296 с.
15. Раменский Л.Г. Основные закономерности растительного покрова и их изучение // Вестн. опытного дела, 1925. – С. 5–32.
16. Сукачев, В.Н. Методические указания к изучению типов леса. – М.: Изд-во АН СССР, 1961. – 144 с.
17. Сукачев В.Н. Основы лесной типологии и биогеоценологии. – Л.: Наука, 1972. – 276 с.
18. Шенников А.П. Введение в геоботанику. – Л.: Изд-во ЛГУ, 1964. – 447 с.
19. Ярошенко П.Д. Основы учения о растительном покрове. – М.: Гео-графгиз, 1953. – 350 с.
|

