Мониторинг и биоразнообразие экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2012. – 140 с.
Список авторов публикаций. С. 139. Предисловие. C.6-8.
Галанин А.В., Галанина И.А. Динамическая парадигма в геоботанике и ботанической географии. С. 9-24.
Урусов В.М., Петропавловский Б.С., Варченко Л.И. К корректировке ботанико-географического и флористического районирования Дальнего Востока России С. 25-45.
Сухомлинов Н.Р. Мониторинг пирогенной трансформации экосистем: проблемы, методы, подходы. С. 46-52.
Игуменова О.П. Экологический мониторинг юго-западной территории республики Башкортостан. С. 53-60.
Киреева Н.А., Якупова А.Б., Григориади А.С. Интегральная оценка устойчивости почвы к нефтяному загрязнению. С. 61-66.
Сухомлинова В.В. Изменение межвидовых стратегических взаимодействий при пирогенной трансформации экосистем. С. 67-74.
Галанин А.В., Долгалева Л.М. К вопросу о применении методики ранжированных невзвешенных вариационных рядов для анализа древостоев на постоянных пробных площадях в Сохондинском биосферном заповеднике. С. 75-92.
Галанин А.В., Галанина И.А. Корейско-Хасанская ботанико-географическая подобласть (очерк растительности). С. 93-114.
Миронова Л.Н., Реут А.А. Итоги репродукции редких и исчезающих видов декоративных травянистых растений в республике Башкортостан. С. 114-118.
Энхтайван Н. Устойчивое лесоуправление в северной Монголии. С. 119-123.
Урусов В.Н., Варченко Л.И. Врищ Д.Л., Петропавловский Б.С. Пути улучшения озеленения Владивостока. С. 123-138.
|

|
УДК 58.00 ББК 72 я 43 М77
Рецензент Христофорова Н.К., д.б.н., проф. ДВФУ
Мониторинг и биоразнообразие экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2012. – 140 с. ISBN 978-5-89694-136-1
Редакционная коллегия: Долгалева Л.М., к.б.н.; Беликович А.В., д.б.н.; Галанин А.В., д.б.н., проф; Петропавловский Б.С., д.б.н.
В сборнике представлены результаты научно-исследовательской и практической работы ученых, преподавателей, аспирантов Дальневосточного региона, в которых отражены глобальные изменения климата и трансформация экосистем, флора, биология и экология растений Сибири и Дальнего Востока, экология природных сообществ, информационные технологии и системы в исследовании экосистем, экологические проблемы сохранения биоразнообразия природных систем Сибири и Дальнего Востока, проблемы охраны и рационального использования животного мира Сибири и Дальнего Востока.
Предназначен для студентов, аспирантов, ученых, специалистов и профессорско-преподавательского состава колледжей и университетов.
© НОУ ВПО «Институт технологии и бизнеса», 2012
|
УДК 581.527:551.432 (571.6)
К КОРРЕКТИРОВКЕ БОТАНИКО-ГЕОГРАФИЧЕСКОГО И ФЛОРИСТИЧЕСКОГО РАЙОНИРОВАНИЯ ДАЛЬНЕГО ВОСТОКА РОССИИ
В.М. Урусов, Б.С. Петропавловский, Л.И. Варченко
V.M. Urusov, B.S. Petropavlovsky, L.I. Varchenko. ON THE ADJUSTMENT OF BOTANICAL-GEOGRAPHIC AND FLORISTIC REGIONALIZATION OF THE RUSSIAN FAR EAST. The authors display conjugacy of the three-dimensional genetic units of relief, or megasmorphostructure of the central type (MСT) of different orders with the floral provinces and the geobotanical and phytogeographical regionalisation, as well as with ranges of endemic alpine prostrate shrubs and some spe-cies of angiosperms. It is shown that MСTs are the principal factors of isolation and divergence of biological systems, the highly-ordinal sections of the disloca-tion of species, coenoses and flora.
Il. 5. Bibl. 87.
Keywords: Floristical regionalization, Russian Far East, flora, central type megamorphostructure, adaptive evolution of plants.
Несмотря на полноту освещения ряда геологических и биологических аспектов природопользования российского Дальнего Востока (ДВ), рациональное природопользование требует дальнейшего углубления, обобщения, разработки и корректировки естественнонаучных представлений. При этом прежде всего необходимо уточнение природного районирования, в т.ч. и флористического, на основе знания определяющих его факторов неживой и живой природы. Решение данного круга вопросов является основой повышения результативности не только научных исследований, но и хозяйствования. В этой связи возникает необходимость выявить соответствие состава и структуры растительного покрова современному климату, а также сопряженность пространственно-генетических единиц рельефа, или мега-морфоструктур центрального типа (МЦТ) разных порядков с флористическими провинциями и геоботаническими районами, а также ареалами эндемичных высокогорных стланцев и некоторых видов покрытосеменных.
Синхронность морфотектонических и биоэволюционных процессов рассматривалась неоднократно (Красилов, 1977; Урусов, 1988, 1998; Урусов, Лобанова, Варченко, 2005), но проблемы флористического районирования и его уточнения оставались в тени или, по крайней мере, не были предметом специальных исследований, в т.ч. для ДВ, до нашей недавней публикации (Урусов, Варченко, Петропавловский, 2009). Необходимо развить и пояснить ряд ее положений. Имея определенный «задел» в этой области, считаем возможным уточнить нашу позицию по ДВ России. Она кратко такова: флористические области и провинции – достаточно древние общности растительности, чтобы быть связанными с чрезвычайно медленной пространственной дифференциацией рельефа, структуры которого на определенных этапах своей истории выступают как фактор изоляции, вычленения очагов эволюции биоты, по крайней мере, на уровне ее макрорефугиумов. Эволюция морфоструктур на позднейших этапах их развития и динамика климата обусловили некоторое несовпадение единиц рельефа и флористических общностей, но следы первоначальных ареалов последних на уровне скальных, подлесочных, пойменных видов и даже экстразональных формаций, как правило, остаются. Горные сооружения, маркируя границы МЦТ, обеспечивают достаточную однородность климата, его сходную континентальность или, наоборот, океаничность и, в итоге, единство флоры внутри МЦТ. В то же время унаследованный внутренний рельеф, например, хребты субмеридеонального простирания, делят флористические провинции на подпровинции или, по крайней мере, на округа. В данном сообщении мы не касаемся иерархии флористических отдельностей в гори-зонтальном и вертикальном пространстве – это специальная очень ответственная тема, видимо, требующая молодых сил.
Основная цель исследований – выявить сопряженность пространственно-генетических единиц рельефа, или МЦТ разных порядков с флористическими областями и провинциями, а также ареалами эндемичных высокогорных стланцев и некоторых видов покрытосеменных для задач возможной корректировки флористического районирования ДВ, а также для выявления наиболее характерных ботанико-географических соотношений, видообразования, миграций растений в ракурсе анализа становления и развития МЦТ разных порядков.
Материал и методика
Объектом исследований явились флора и растительный покров материковой части юга ДВ в пределах Сихотэ-Алиня, Восточно-Маньчжурских гор и Сахалинской области, Камчатского края и водосбора Охотского моря. В результате многолетних исследований изучены состояние субальпийских стланиковых группировок на верхнем пределе растительности, закономерности усложнения растительного покрова в процессе почвообразования в предгольцовой зоне, а также на скалах низкогорий и морского побережья и на песчаных и галечных береговых валах.
Смены доминантов и типов растительного покрова в пространстве – и отчасти во времени – для Приморья и Приамурья по геоботаническим и лесотаксационным описаниям и документам установлены с 1859 г. (Маак, 1861; Будищев, 1867: Пржевальский, 1870; Коржинский, 1899; и др.), для крайнего юга Приморья и сопредельных стран – с 1882 г. (Барабаш, 1883; Вебель, 1890; Стрельбицкий, 1987; и др.), а также по материалам ревизий лесоустройства - с 1913 г. и характеру синузий трав, что уже апробировано Г.Э. Куренцовой (1968а, б; и др.) и дает основание для реконструкции растительного покрова, по крайней мере, в период голоценового климатического оптимума.
Для голоцена и антропогена в целом растительный покров неоднократно реконструирован по спорово-пыльцевым спектрам и иногда остаткам растений (Урусов, 1988). Для заповедника «Кедровая Падь», Восточно-Маньчжурских гор, хребта Ливадийский, водосборов рек юга Приморья смены доминантов лесных экосистем прослеживаются по деревьям-долгожителям, например, Taxus cuspidata Siebold et Zucc., Betula schmidtii Regel и их долго, иногда полутысячелетия не разлагающимся стволам (Урусов и др., 2010). Ареалы сосудистых растений уточнялись по сводкам и флорам для бывшего СССР, ДВ и сопредельных регионов (Воробьев, 1963, 1968; Ohwi. 1965; Соколов, Связева, 1965; Положий, 1968; Соколов и др., 1977; Толмачев, 1974; Сосудистые растения советского Дальнего Во-стока, тт. 1-8, 1985-1996), по фототаблицам в палеоботанических сводках, которые отчасти позволили определить геологический возраст ряда видов (Фотьянова, 1988; Аблаев и др., 1994; Тащи и др., 1996). Соотношение спорово-пыльцевых спектров с синхронной им растительностью для ДВ подробно рассмотрено В.М. Урусовым (1988). Шесть ритмов развития плейстоцен-голоценовой растительности ДВ приняты по А.М. Короткому с соавторами (1980), отнесшим ближайшие к нашему времени ледниковые эпохи (их 4) к позднему плейстоцену.
Периодизация голоцена принята по Н.А. Хотинскому (1977), возраст флорогенетических элементов – по А.Н. Криштофовичу (1936), В.Б. Сочаве (1945), В.Н. Васильеву (1958) с уточнениями в соответствии с характером ареалов, биологией и динамикой конкретных формаций в неогене-антропогене.
Флористическое районирование А.Л. Тахтаджяна (1970, 1978) дополнено геоботаническим Г.В. Крылова (1962) и Б.П. Колесникова (1969), а
также отдельными разработками для Сахалина (Толмачев, 1955а) и Курильских островов (Воробьев, 1963).
В качестве низшей классификационной единицы растительных сообществ вслед за Г.Э. Куренцовой (1967; и др.) принята ассоциация, являющаяся совокупностью участков определенного флористического состава в определенных условиях существования, в том объеме, как ее рассматривают В.Н. Сукачев (1915, 1935), В.В. Алехин (1935, 1938), Б.П. Колесников (1938, 1956), П.Д. Ярошенко (1953, 1961, 1969), Б.А. Быков (1957) и др. Высшие единицы классификации лесных ассоциаций приняты по Б.П. Колесникову (1956) и сводятся к группам типов леса и их геоморфологическим комплексам, которые выделены на основе сходства в положении на склоне, его экспозиции и крутизне.
Материалы по пространственно-генетическим единицам рельефа, или МЦТ разных порядков привлечены из монографий и статей геоморфологов школы Тихоокеанского института географии ДВО РАН (Кулаков, 1980 а, б, 1984, 1986; Худяков, 1968, 1988; Худяков, Кулаков, Тащи, 1980; Худяков, Денисов, Короткий и др., 1972; и др.), причем для данной работы особенно важны морфоструктуры 3-го порядка диаметром около 1 тыс. км, имеющие на краю Азии обширные равнины или моря в центральной зоне. Эти МЦТ и определяют географию провинциальных флор и отграничены горными хребтами преимущественно мелового возраста. Более древними хребтами маркируются кольцевые структуры рельефа в разы большего диаметра, с которыми в значительной мере совпали простирания флористических областей А.Л. Тахтаджяна (1978) и флористические построения М.Г. Попова (1969, 1983 и др.).
Результаты и их обсуждение
Сопоставление пространственно-генетических единиц рельефа, или МЦТ разных порядков, понимаемых в динамике тектонических процессов и как географическая определенность (Худяков и др., 1980) с флористическими провинциями (Тахтаджян, 1978), подпровинциями и геоботаническими районами, ареалами эндемичных высокогорных стланцев, прежде всего хвойных, например, сабиновых можжевельников (у них листья и элементы шишкоягоды соединены в мутовки попарно), но также и некоторых покрытосеменных, уточненными ареалами доминирующих хвойных и других видов-эдификаторов и индикаторов ландшафтов вскрывает несомненную связь между этими пространственными системами.
Суть ее не только в том, что рельеф определяет (или в своей зоне, по крайней мере, перераспределяет) климатические условия и, в итоге, особенности распределения растительных зон и формаций, а также и флоры (Сочава, 1986). Гораздо важней, что именно МЦТ являются главнейшими факторами изоляции и дивергенции биологических систем, высокопорядковыми участками дислокации видов, ценозов и флор, по которым осуществляется географическая и экологическая дифференциация, идет выравнивание состава популяций и сообществ. На уровне соседних МЦТ третьего порядка изоляцией на протяжении геологических веков формируются виды, сходные как звенья одного географического или эколого-географического видового ряда.
МЦТ (их обоснование и ранжирование мы считаем одним из важнейших достижений Тихоокеанского института географии ДВО РАН, которое фито- и биогеографам и биологам-эволюционистам еще предстоит осмыслить) – динамическое явление, развивающееся как: 1) поднятие гигантского свода со сложным рельефом, 2) его стабилизация и конструктивное развитие, 3) общее тектоническое погружение и формирование впадин окраинных морей. Возраст фаз жизни МЦТ соответственно более 0,5 млрд лет, более 230 млн. лет, примерно от 200 до 1–2 млрд лет (Худяков и др., 1980; Кулаков, 1986).
Таким образом, факторы изменения среды и изоляции действуют громадные промежутки времени, достаточные для микро- и макроэволюционных процессов, идущих в целом за геологическими событиями (Васильев, 1958). Вполне вероятно, что фаза сводового поднятия в этот период пространственно сжатых гигантских МЦТ является временем ускоренного видообразования и формирования центра видового разнообразия сосудистых растений и не только их. Потому что при подъеме горного узла или отдельных горных систем осуществляется одно из самых важных условий эволюции – обособление мутаций от родительской популяции. Расселение из этого центра начинается уже в данной фазе и продолжается в фазы расширения и стабилизации и, в гораздо меньшей мере, тектонического погружения. Вот так расселялись древние – на уровне видовых рядов – и третичные субальпийские стланики.
Вскрытая П.Г. Горовым с соавторами (1979) самостоятельность и особая архаичность Bupleurum komarovianum Lincz. и B. scorzonerifolium Willd. как раз связана с их формированием, соответственно, в Японской и Амурской МЦТ и расселением преимущественно на северо-запад и запад. Приведенный карио-морфологический материал доказывает в целом восточноазиатское происхождение рода Bupleurum (Горовой и др., 1979) и, вероятно, целого ряда других родов, включая Lilium, Betula, Lonicera.
Специфика, особенности современной флоры и растительности во многом определяются изоляцией видов и формаций в замкнутых прогибах, в том числе и на берегах формирующихся окраинных морей. Чем значительней эта изоляция, чем полней сохраняются автохтонные виды, тем существенней своеобразие современной флоры, прослеживающееся, кстати, и на крайнем северозападе Амурской морфоструктуры – на юге Бурятии. Здесь от Малханского хребта на восток и юго-восток нарастает участие характерных Маньчжурской флористической провинции видов разных жизненных форм, что хорошо освещено в литературе (Коржинский, 1899;
Комаров, 1901; Пешкова, 1968; Коропачинский, 1980, и др.; Малышев, Пешкова, 1984; Положий, Крапивина, 1985) и наблюдалось нами в природе.
То же самое относится и к крайнему северо-востоку Амурской морфоструктуры, где еще произрастают виды зимнеголых лесов (Воробьев, 1937; Соколов, Связева, 1965; Чащин, 1966; Соколов и др., 1977) и даже растения, типичные для берегов пресных водоемов в зоне ультранеморальной растительности (Пехтерев, 1963; и др.).
Если в фазах воздымания и стабилизации МЦТ характерны условия, способствующие видообразованию на основе ядра местных видов, например, лесообразователей неморальной и бореальной вертикальных зон, ландшафтообразующих стланцев субальп, а эдафические условия таковы, что расселение растений идет преимущественно к периферии структур и за их пределы [по течению рек, перестройка которого отчасти все еще маркируется в т.ч. расселением древнейших северопацифических псаммофитов из родов Leimus, Papaver, Ligusticum, Glehnia и др. (Урусов, 1998)] , что обеспечивает, с одной стороны, изоляцию местной флоры, не принимающей или почти не принимающей внедренцев, с другой, распространение новообразований в другие регионы, то на фазу тектонического прогиба приходится период вбирания аллохтонных видов и формаций, особенно на этапах разрушения кольцевых горных сооружений окраин МЦТ.
Общеизвестным условием образования зон гибридизации у прежде изолированных таксонов является именно разрушение физических географических барьеров, что мы полностью принимаем. Поэтому инвазия видов и их гибридизация – самые молодые явления во флоре МЦТ. Вот так сформировались гибриды елей, берез и таволг в среднегорьях Сихотэ-Алиня, лиственниц, ольхи, ясеней, роз в низкогорьях. Формирование окраинных морей ДВ обнажает новое пространство ресурсов, например, в береговой зоне, ширина которой в системе стадиал-межстадиал смещается в пределах от первых десятков до 150 км в зоне Японского моря, где, во-первых, под действием ряда факторов, воздуха и почвы, включая химизм, растет на порядки частота мутаций, во-вторых, становятся обычными полиплоиды разного геологического возраста (Пробатова и др.,1984), в-третьих, значителен четвертичный эндемизм. Именно здесь сформированы молодые виды и подвиды хвойных (Урусов, Лобанова, Варченко, 2007; Урусов и др., 2010).
Пожалуй, стоит упомянуть вот о чем: северное формирование и смещение на юг внетропических флор, обоснованное М.Г. Поповым (1969, 1983) является частным, хотя и ярчайшим примером возникновения видов и флор на уровне Арктической МЦТ второго порядка, открытой Г.И. Худяковым (1988) к северу от 62-64° с.ш. Тогда данная МЦТ обеспечила как минимум формирование циркумполярной Арктической флористической провинции и еще ряда североевразиатских и североамериканских провинций флоры. Разумеется, в холодные эпохи их границы сдвинулись к югу.
Енисейско-Курильская МЦТ 2-го порядка (Ежов, Андреев, 1989) обеспечила формирование по крайней мере основных флористических провинций Восточноазиатской флористической области с ее сабиновыми можжевельниками Juniperus (= Sabina) davurica Pall., J. sargentii (A. Henry) Takeda ex Koidz., аянскими и корейскими ельниками, смешанными и зимнеголыми лесами. Уже это – наличие ядра видообразования и ценогенеза в средних широтах, сопоставимого с центром арктического видообразования М.Г. Попова, – позволяет скорректировать, если не отвергнуть идею монофилетического происхождения внетропичексих флор Северного полушария Земли.
Итак, максимальное проявление в пространстве и, наконец, энтропия МЦТ обусловливают дивергенцию видов, сообществ и флор и их последующие регресс, консолидацию, гибридизацию и замену. МЦТ третьего порядка (в т.ч. Охотской, Японской, Корейской с их в каждом случае более чем тысячекилометровым диаметром), как правило, соответствует один вид видового ряда или секции. Ареалы близких видов перекрываются в зонах сближения или наложения окраин морфоструктур, что подтверждает существенность роли МЦТ при похолоданиях антропогена в качестве макрорефугиумов. Эти зоны сближения являются естественными районами смешения и гибридизации близких видов, наложения флор, где любое районирование затруднено. Яно-Колымской, Охотской, Амурской, Японской (Япономорской), Восточно-Китайской мегаморфоструктурам 3-го порядка (Кулаков, 1980 а, б, 1984, 1986; Худяков, Кулаков, Тащи, 1980) соответствуют Северо-Восточносибирская, Охотско-Камчатская, Маньчжурская, Японо-Корейская, Северо-Китайская флористические провинции А.Л. Тахтаджяна (1970, 1978). Сахалино-Хоккайдcкой провинции А.Л. Тахтаджяна соответствует лишь зона сближения кольцевых морфоструктур (рис. 1).
Японо-Корейская флора достаточно неоднородна, чтобы быть разделенной на Японскую и Корейскую провинции с изначально субтропическим в пределах собственно Японской МЦТ и зимнеголым ядром экосистем и видов (Корейская МЦТ А.П. Кулакова, вмещающая как корейские, так и китайские виды и формации, присутствующие и на японском о-ве Кюсю, например, сосна Арманда Pinus armandii Franch).
 |
В этом причина близости флоры юга Сахалина и Курил охотско-камчатской флоре с одной стороны, с другой – маньчжурской и японо-корейской. Переходный характер Сахалино-Хоккайдcкой флористической провинции как раз и обусловлен отсутствием собственной мегаморфоструктуры центрального типа. Уточнение флористического районирования, например, юго-восточной части Хабаровского края, предопределено характером Охотской МЦТ гораздо основательней вдающейся в Сихотэ-Алинь, чем Охотско-Камчатская флористическая провинция на схеме А.Л. Тахтаджяна.
Находка пихты сахалинской Abies x sachalinensis Fr. Schmidt с ее характерными подлесочными видами в Советскогаванском и Ванинском районах Хабаровского края позволяет несколько расширить границы Сахалино-Хоккайдской флористической провинции в сторону материкового побережья Татарского пролива (Урусов, 1988; Урусов, Чипизубова, 2000; Уру-сов, Лобанова, Варченко, 2007). В то же время следует уточнить ее предел в зоне Средних Курил, где по о-в Кетой, как минимум, господствует хоккайдская флора (Воробьев, 1963; Урусов, Лобанова, Варченко, 2007).
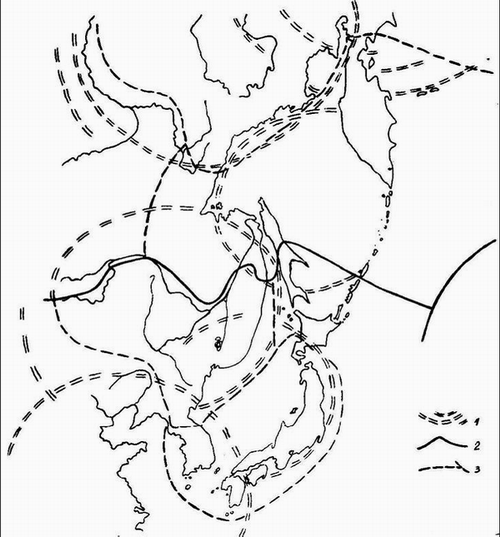
Рис. 1. Границы флористических провинций на востоке Азии в значительной мере определяются мегаморфоструктурами центрального типа (МЦТ). Выраженность МЦТ в пространстве и зоны их наложения сами по себе могут служить ориентиром для уточнения флористического районирования.
1 – контуры МЦТ 3-го порядка (Кулаков, 1986), 2 – границы флористических областей, 3 – границы провинций (Тахтаджян, 1970, 1978), 4 –контуры Арктической Г.И. Худякова (1988 и др.) и Енисейско-Курильской МЦТ Б.В. Ежова и В.Л. Андреева (1989). По Худякову и Ежову приводим МЦТ 2-го порядка.
|
Для ландшафтных формаций МЦТ характерными сообществами и видами являются:
- Яно-Колымской МЦТ: формация кедровостланиковая Pinus pumila (Pall.) Regel, виды: кедровый стланик и лиственница Каяндера (Larix cajanderi Mayr);
- Алданской МЦТ – лиственница Гмелина (L. gmelinii (Rupr.) Rupr).;
- Охотской МЦТ – лиственница камчатская (L. kamtschatica (Rupr.) Carr.) и ель аянская (Picea ajanensis (Lindl. et Gord.) Fisch. ex Carr.), березы: Эрмана (Betula ermanii Cham). и Тауша (B. tauschii (Regel) Koidz.), рябина бузинолистная (Sorbus sambucifolia Cham. et Schlecht.), курильский бамбук (Sasa kurilensis (Rupr.) Makino et Shibata) и тисовый стланик, или тис карликовый (Taxus nana (Rehd.) Urussov.) Тис карликовый – особый – даже не восточно-азиатский, а по своему генезису, скорее, западносевероамериканскиий вид, монтанную западнопацифическую природу которого предполагал еще В.Л. Комаров (1901), что и подтвердилось;
- Амурской МЦТ – сосна кедровая корейская (кедр корейский) (Pinus koraiensis Siebold et Zucc.), ели: мелкосеменная (Picea microsperma Carr.) и корейская (P. koraiensis Nakai), пихта белокорая (Abies nephrolepis (Trautv.) Maxim.), лиственницы: ольгинская (Larix olgensis A. Henry) (в настоящее время единственная эндемичная лиственница этой МЦТ), лиственница Гмелина (L. gmelinii (Rupr.) Rupr) и ряд гибридных: лиственница амурская (L.x amurensis Kolesn.), лиственница охотская (L. x ochotensis Kolesn.), лиственница приморская (L. x maritima Sukacz.), лиственница Комарова (L. x komarovii Kolesn.), лиственница Любарского (L. x lubarskii Sukacz.); дуб монгольский (Quercus mongolica Fisch. ex Ledeb.); березы: маньчжурская (Betula mandshurica (Regel) Nakai), даурская (B. davurica Pall.) и ребристая (B. costata Trautv.), а также «свита» япономорских видов (в Амурской структуре они являются доминантами коренных ландшафтов юго-восточного сектора, но унаследованы от Японской – Япономорской – МЦТ): сосны: густоцветковая (Pinus densiflora Siebold et Zucc.), Тунберга (P. thunbergii Parl.), мелкоцветковая (P. parviflora Siebold et Zucc.), густоцветковая – Тунберга (P. x densi-thunbergii Uyeki) (гибрид); можжевельник Саржента (Sabina (= Juniperus) sargentii Nakai.), дуб курчавый (курчавенький) (Quercus mongolica Fish. ex Ledeb. subsp. crispula (Blume) Menitz.) и т.п.;
- Корейской МЦТ – пихта цельнолистная (Abies holophylla Maxim.), сосна китайская (Pinus tabulaeformis Hort. ex Carr.), сабиновый можжевельник китайский (Sabina chinensis (L.)Ant.), ряд видов дубов и берез.
 |
Фазам развития морфоструктур должны соответствовать этапы видообразования, так как при сводовом воздымании создаются условия для вычленения стланцев, которые из-за особенностей ортоселекции и распространения могут быть локализованы в определенных горных системах или в ряде горных систем, теперь отчасти или в очень значительной мере ушедших под уровень моря (рис. 2).
Микробиота перекрестнопарная (Microbiota decussata Kom.) происходящая из позднего мела или олигоцена, формирует верхнюю границу растительности только в Сихотэ-Алине, в отличие от сабины Саржента, практически обрамляющей в высокогорьях водосбор Японского моря, не обязательно моложе этой сабины. Напомним, что у микробиоты нет распространителей. Ее семена осваивают новые территории только в процессе роста ее ветвей (Урусов, 1979; Урусов, Лобанова, Варченко, 2007). Следовательно, орнитохория всегда способствовала ускоренному освоению подходящих экотопов как в пределах генетических общностей, так и вне их. Виды с выраженной орнитохорией еще и сегодня позволяют вскрыть характер палеоландшафтов внутри МЦТ 3-го порядка.
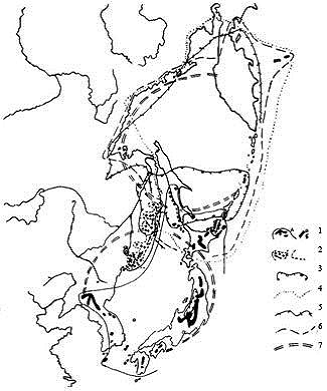
Рис. 2. Некоторые автохтонные субальпийские и ультрабореальные виды растений, ареалы которых маркируют зоны древних высокогорий. В пределах одной МЦТ выделяются участки с эндемами разного ранга, с особым генезисом флоры.
1 – Sabina (Juniperus) sargentii (А – распространение в горах и граница ареала, Б – популяции, смещенные на прибрежную полосу), 2 – Microbiota decussata (основные массивы формации в высокогорьях Сихотэ-Алиня, вероятные пути расселения вида и граница ареала); границы ареалов: 3 – кустарникового вида тиса Taxus nana, 4 – Betula ermanii, 5 – Lonicera chamissoi, 6 – Sorbus sambucifolia по уточненным данным, 7 – границы Охотской и Японской МЦТ, 8 – граница Арктической МЦТ. |
 |
В то же время представляется древним возраст каменных берез и их дивергенция на березу Эрмана и березу шерстистую. По крайней мере, ареал Betula ermanii (нам удалось установить наличие ее береговых популяций к северу от пос. Де-Кастри в Хабаровском крае в 2001 г.) очень близок ареалу Sorbus sambucifolia, что, видимо, подтверждает их происхождение в горах на месте современного Охотского моря.
Становление современных настоящих и кедровых сосен и дубов родства дуба монгольского на ДВ связано, по-видимому, с формированием прогибов в центре МЦТ третьего порядка и изоляцией внутри морфоструктур, а также с процессами, включающими по длительности значительную часть времени существования гигантских МЦТ (рис. 3, 4).
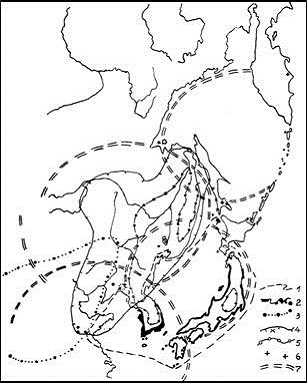
Рис. 3. Ареалы сосен и некоторых других растений на востоке Азиатского материка довольно четко вписываются в МЦТ, которые являются (или являлись) в дочетвертичное время - областями видообразования, а в стадиалы – макрорефугиумами.
Ареалы: 1 – Pinus densiflora, 2 – P. thunbergiana с местонахождениями ее гибридов в ареалах викарных видов, 3 – P. tabulaeformis, 4 – Quercus mongolica и его гибридов, 5 – P. koraiensis, 6 – известные популяции Abies gracilis, 7 – границы Охотской, Амурской, Японской, Корейской мегаморфоструктур центрального типа по А.П. Кулакову (1986), 8 – граница Арктической МЦТ Г.И. Худякова (1988) |
 |
Зарождение дивергенции дубов может быть обеспечено разными климатическими режимами внешних приокеанических и внутренних континентальных структур, в основном существующих как поднятия. Крайне маловероятен раннеплейстоценовый возраст Pinus pumila. Его обособление от Pinus sibirica Du Tour во второй половине раннего плейстоцена И.Ф. Удра (1978) обосновывает исходя из возраста ископаемой пыльцы сосны кедровой сибирской на Колымском нагорье и присутствия кедровниковой популяции на о-ве Карагинский: кедровый стланик должен был образоваться и расселиться на данную территорию до ее отделения от по-ва Камчатка.
Рис. 4. Наполнение флороценотипами «кедровников» ДВ Амурской МЦТ свидетельствует об их ландшафтной роли на ее севере не позже начала квартера.
1 – границы РФ и ДВ, 2 – контур Амурской МЦТ, 3 – генерализованные ареалы Pinus koraiensis, 4 – Euonymus sacresancta Koidz., 5 – Eleuterococcus senticosus (Rupr. et Maxim.) Maxim.
|
Но, во-первых, кедровый стланик не принадлежит к эколого-географическому надряду «кедров» корейского и сибирского, т.к. существенно различается с ними по строению женских шишек. Он ближе к североамериканской Pinus albicaulis Engelm. и вместе с ней выделен Е.Г. Бобровым в особый ряд (Бобров, 1978: 118; Урусов, Гущин, 1985).
Во-вторых, присутствие кедрового стланика по недавнее время на о-ве Медный, Командоры, где Российско-Американская компания интродуценты не вводила (Васильев, 1957; Кабанов, 1977; Бобров, 1978; Русская Америка…, 1979), снимает необходимость определять возраст вида отделением о-ва Карагинский (Камчатская область). В то же время гипотеза возникновения кедрового стланика на Колымском нагорье и расселение его на Камчатку и Чукотку уже после распада Берингии (Удра, 1978) вполне вероятна, как вероятен и поздний приход вида на о-в Сахалин, юг Курил и в Японию. По данным палеогеографов, ТИГ ДВО РАН это произошло поэтапно около 100 и 65-20 тыс. лет назад. Изучение генэкологии Pinus pumila поможет уяснить его миграционные пути и, возможно, более сложную историю и географию расселения. По крайней мере, часть курильских популяций кедровника по набору морфологических форм не имеет аналогов на о-ве Сахалин.
МЦТ более низкого ранга могут соответствовать молодые эндемичные виды, например, тимьяны или подвиды и расы. Локализация в их пределах древних хвойных может быть и вторичной. В данном случае само понятие молодости таксона должно рассматриваться на фоне процессов, охватывающих сотни и многие млн лет. Сам по себе ареал может свидетельствовать о возрасте таксона, если возможна достаточно точная интерпретация конкретного ареала. Во всяком случае более широкие ареалы можжевельников ряда обыкновенных, елей с четырехгранной хвоей являются достоверным признаком их большей древности. Сабиновые можжевельники имеют более локальные ареалы еще и потому, что они моложе.
Активные вертикальные перемещения поверхности Земли разного знака, генеральный морфоструктурный перекос в сторону океана, формирование окраинных морей увязаны с последним этапом развития окраинноматериковых МЦТ, этапом тектонического погружения (Худяков, Кулаков, Тащи, 1980; Кулаков, 1986). Воздымание Больших Курил и юга Сахалина при наличии лестницы береговых террас на суше по крайней мере до абсолютной высоты до 250 м и под водой до глубины около 1 тыс. м (Кулаков, 1973; и др.) свидетельствует о динамизме и данного этапа, возможности ротации, видимо, в рамках более общих процессов пульсации Земли и единого галактического ритма, биогеографический смысл которых рассматривается, например, А.В. Галаниным (1999).
Развитием окраинно-материковых МЦТ вопрос о том, на сколько же метров опустился край Азии, решается дифференцированно. Как уже говорилось, дифференцированный ответ дает и положение субальпийцев, в
особенности способных существовать и под пологом леса. Положение микробиотовой формации Microbiota decussata Kom. или формаций сабины Саржента и кустарникового тиса Taxus nana показывает, как конкретно и насколько неравномерно с перекосом к морям и югу, к 43–45° с.ш. на берегах Приморья и на 46–47° с.ш. на Сахалине шло погружение Сихотэ-Алиня или Сахалина (Толмачев, 1956; Урусов, 1988). Биология видов и реконструкция палеоклимата позволяют примерно определить высоту горных сооружений в геологическом прошлом, косвенно устанавливая величину еще более древних поднятий поверхности Земли. Становится понятным сниженное положение физиологически (биологически) высокогорных видов в интразональных сообществах или в качестве экстразональной растительности.
Современные флористические провинции ДВ первоначально формировались в пределах гигантских МЦТ третьего порядка за геологическое время, превышающее возраст их характерных видов, который может составлять в т.ч. десятки млн лет (Урусов, Лобанова, Варченко, 2007). Провинциальные флоры типично выходили за пределы своих МЦТ. Но глобальное похолодание, унаследованное с миоцена, привело к их территориальному сжатию и отступлению на юго-восток и юг с одновременным уцелеванием в основном подлесочных и лесостепных видов в оптимальных по климату урочищах или на инсолируемых склонах и известняковых скалах в пределах древней границы провинции.
 |
Ротационное изменение климата в системе стадиал-межстадиал, когда за 90–100 (110) тыс. лет на юге ДВ среднегодовые температуры менялись от 0–1°С до +7, +8° и обратно, а суммы активных температур менялись от 1600 до 3100° (Величко, 1973) обусловливало смещение провинциальных флор к югу. Этому способствовало в ледниковье закрытие Корейского пролива Японского моря, субконтинентализация Японии и Сахалина, ликвидация теплого течения в Японском море.
Эти процессы все еще сказываются: минимизируется территория неморальных и квазибореальных флор, возросла вероятность обнаружения экстразанальных сообществ Abies x sachalinensis, A. mayriana, Pinus koraiensis, Quercus monglica, Q. aliena Blume, Q wutaishanica H. Mayr, Alangium platanifolium (Siebold et Zucc.) Harms в зонах со смягченными зимами и более равномерным увлажнением. Причем падение континентальности климата как раз гарантировало уцелевание повышенного биологического разнообразия в некоторых урочищах ДВ, включая возвышенности в низкогорьях, в т.ч. в ледниковое время.
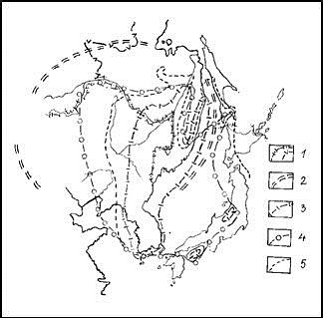
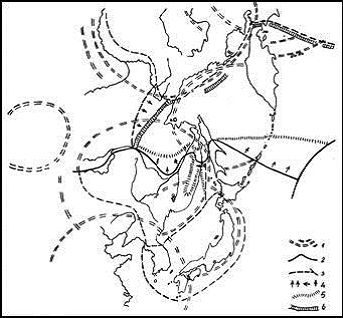
Рис. 5. Желательные изменения флористического районирования в пределах ДВ в связи с уточнениями ареалов видов и ценозов после 1980 г.
1 – контуры гигантских структур рельефа – МЦТ, 2 и 3 – границы флористических областей и провинций на схеме акад. А.Л. Тахтаджяна (1978 и др.), 4 – наши предложения по смещению границ областных и провинциальных флор, 5 – уточненные флористические границы, 6 – северный предел Восточноазиатской флористической области в нашем понимании |
В связи с этим можно предвидеть сдвиг на север общей границы Восточноазиатской флористической области при одновременном исключении из Циркумбореальной области и включении в Восточноазиатскую флористическую область всей Охотско-Камчатской провинции и выводе на материк Сахалино-Хоккайдской. Это более отвечает контуру Восточноазиатской МЦТ второго порядка (Ежов, Андреев, 1989), а также ареалам дальневосточной каменной березы Эрмана, рябины бузинолистной и крупнотравного комплекса и комплекса бореальных и субальпийских видов рода Sasa. В то же время центральные хребты Среднего и Северного Сихотэ-Алиня вслед за В.К. Арсеньевым (1912) и В.А Недолужко (1995) также есть осно-вание сближать с Сахалино-Хоккайдской флористической провинцией – они составляют переход между Маньчжурской и Сахалинской флорами. Эти выявленные ботанико-географические соотношения позволяют уточнить флористическое районирование для всего ДВ (рис. 5).
Выводы
1. Результаты исследований подтвердили гипотезу о синхронности геологических процессов, филогенеза и генезиса сообществ. МЦТ являются важным критерием целостности, естественности флористических областей, их географической определенности и облегчают познание генезиса флор. МЦТ 3-го порядка являются не только исходными единицами для регионального морфоструктурного районирования, но и для районирований флористического и геоботанического.
2. Гляциальные циклы из-за их большой скорости были не столько факторами эволюции, сколько прежде всего элиминации, гибридизации, смещения на юг и юго-восток и «сжатия» восточноазиатских флор и только в третью очередь – расо- и подвидообразования (крайне редко видообразования) за счет жестких популяционных волн, сохраняющих уникальные изореагенты или формы.
3. Флоры в пределах крупных МЦТ на востоке Азии были изолированы в позднем мезозое и значительную часть кайнозоя и даже весь кайнозой (преимущественно субальпийские и бореальные виды) или в неоген-четвертичное время (ультранеморальные, т.е. более теплолюбивые дубравные и неморальные виды). И только деструкция морфоструктур облегчила перемещение растений за их пределы и встречу викарных видов.
4. Собственно миграции свойственны прежде всего бореальным, в том числе криофильным и ксерофильным элементам, которым благоприятствовала климатическая ситуация перехода к стадиалам. Представители квазибореальных южнотаежных и неморальных формаций не отступают, а гибнут на месте или выживают как малочисленные ценопопуляции речных пойм, бортов долин, южных склонов в низкогорьях, т.е. участков с оптимальным местным климатом.
5. Есть все основания связать происхождение видов с циркумполярными ареалами с развитием Арктической МЦТ, а Восточноазиатскую флори-стическую область расширить за счет Охотско-Камчатской провинции Циркумбореальной флористической области А.Л. Тахтаджяна.
6. МЦТ 3-го порядка юга ДВ составлены горными системами, маркируемыми разными субальпийскими стланиками-почвообразователями, что само по себе может быть использовано для разделения флористических провинций на подпровинции и, возможно, отнесения Северного Сихотэ-Алиня к Сахалино-Хоккайдской провинции А.Л. Тахтаджяна в ранге подпровинции.
7. Целостность простирания Охотско-Камчатской флористической провинции на побережье Охотского моря, в т.ч. Магаданской области подтверждается произрастанием здесь лесов из Betula ermanii с Sorbus sambucifolia, Maianthemum dilatatum (Wood) Nels. et Macbr., что обеспечено низкой континентальностью климата – коэффициент континентальности около 2,5 (Скрыльник, Скрыльник, 1976), а следовательно, смягчаемыми влиянием океана зимами береговой зоны.
Литература
1. Аблаев А.Г., Тащи С.М., Васильев И.В. Миоцен Ханкайской впадины Западного Приморья. – Владивосток: Дальнаука, 1994. – 168 с.
2. Алехин В.В. Основные понятия и основные единицы в фитоценологии // Сов. Ботаника. 1935. – № 5. – С. 21–34.
3. Алехин В.В. Методика полевого изучения растительности и флоры. 2-изд. – М.: Изд. Наркомпроса, 1938. – 208 с.
4. Арсеньев В.К. Краткий военно-географический и военно-статистический очерк Уссурийского края 1901–1911 гг. – Хабаровск: Штаб Приамурского военного округа. 1912. – 341 с.
5. Барабаш. Записки о Маньчжурии // Сб. геогр., топогр. и стат. материалов по Азии. – Спб., 1883. Вып. 1. С. 68–98.
6. Быков Б.А. Геоботаника. 2-е изд., испр. – Алма-Ата: Изд-во АН Каз-ССР, 1957. – 382 с.
7. Бобров Е.Г. Лесообразующие хвойные СССР. – Л.: Наука. 1978. – 188 с.
8. Будищев А.Ф. Описание лесов Приморской области. Сб. главнейших официальных документов по управлению Восточной Сибири. Т. 5. Леса Приморского края, вып. 1. Иркутск. 1883. – 537 с.
9. Васильев В.Н. Закономерности процесса смен растительности // Матер. по истории флоры и растительности СССР. Т. II. – М.: Л.: Изд-во АН СССР, 1946. – С. 365–406.
10. Васильев В.Н. Растительность Анадырского края. – М.: Л.: Изд-во АН СССР, 1956. – 218 с.
11. Васильев В.Н. Флора и палеогеография Командорских островов. – М.; Л.: Изд-во АН СССР. 1957. – 260 с.
12. Васильев В.Н. Происхождение флоры и растительности Дальнего Востока и Восточной Сибири // Матер. по истории флоры и растительности СССР. Т. III. – М.: Л.: Изд-во АН СССР, 1958. – С. 361–457.
13. Вебель. Поездка в Корею летом 1889 г. // Сб. геогр., топогр. и статист. Материалов по Азии. – Спб., 1890. Вып. 41. – С. 143–252.
14. Величко А.А. Природный процесс в плейстоцене. – М.: Наука, 1973. – 256 с.
15. Воробьев Д.П. Растительность южной части побережья Охотского моря // Тр. ДВФ АН СССР. Сер. бот. Т. 2. – Л.: Изд-во АН СССР. 1937. – С. 19–102.
16. Воробьев Д.П. Растительность Курильских островов. – М.: Л.: Изд-во АН СССР, 1963. – 92 с.
17. Воробьев Д.П. Дикорастущие деревья и кустарники Дальнего Восто-ка. – Л.: Наука, 1968. – 227 с.
18. Галанин А.В. Гипотеза пульсирующей Земли с точки зрения биогеографа // Исследование и конструирование ландшафтов Дальнего Востока и Сибири. Вып. 4. – Владивосток:, 1999. – С. 291–329.
19. Горовой П.Г., Кетриц Л.М., Гриф В.Г. Таксономическое и кариоло-гическое изучение Bupleurum komarovianum Lincz. и Bupleurum scorzonerifolium Willd. (Apiaceae) из Приморья // Бот. ж. 1979. Т. 64. № 1. С. 42–46.
20. Ежов Б.В., Андреев В.Л. Оруденение в морфоструктурах центрального типа мантийного заложения. – М.: Наука, 1989. – 126 с.
21. Кабанов Н.Е. Хвойные деревья и кустарники Дальнего Востока. – М.: Наука, 1977. – 175 с.
22. Колесников Б.П. Растительность восточных склонов Среднего Си-хотэ-Алиня // Тр. Сихотэ-Алинского государственного заповедника. 1938. – Вып. 1. – С. 25–207.
23. Колесников Б.П. Кедровые леса Дальнего Востока. – М.: П.: Изд-во АН СССР, 1956. – 262 с. (Тр. ДВФ АН СССР. Сер. Бот. Т.2 (4).
24. Колесников Б.П. Растительность // Южная часть Дальнего Востока. – М.: Наука, 1969. – С. 206–250.
25. Комаров В.Л. Флора Маньчжурии. Т. 1. – СПб.: 1901. – 559 с.
26. Коржинский С.И. Растительность России // Энциклопед. словарь Брокгауза и Эфрона. 1899. – Т. 27а. – С. 42–49.
27. Коропачинский И.Ю. Происхождение и состав арборифлоры Сибири // Проблемы лесной биогеоценологии. – Новосибирск: Наука, 1980. – С. 56–72.
28. Короткий А.М., Караулова Л.П., Троицкая Т.С. Четвертичные отложения Приморья. – Новосибирск: Наука, 1980. – 234 с.
29. Красилов В.А. Эволюция и биостратиграфия. – М.: Наука, 1977. – 256 с.
30. Криштофович А.Н. Основные пути развития флоры Азии // Уч. Зап. ДГУ. 1936. – Т. 9. – вып. 2. – С. 95-113.
31. Крылов Г.В. Лесные ресурсы и лесорастительное районирование. – Новосибирск: Изд-во СО АН СССР, 1962. – 240 с.
32. Кулаков А.П. Четвертичные береговые линии Охотского и Японского морей. – Новосибирск: Наука, 1973. – 187 с.
33. Кулаков А.П. Морфотектоника и палеогеография материкового побережья Охотского и Японского морей в антропогене. – М.: Наука, 1980. – 175 с.
34. Кулаков А.П. Морфотектоника и особенности развития северного побережья Охотского моря в антропогене // Геолого-геоморфологические конформные комплексы Дальнего Востока. – Владивосток: ДВНЦ АН СССР, 1980б. – С. 48–70.
35. Кулаков А.П. Мегаморфоструктуры центрального типа – основа региональных иерархических систем востока Азии // Морфоструктуры центрального типа Дальнего Востока. – Владивосток: ДВНЦ АН СССР, 1984. – С. 14–26.
36. Кулаков А.П. Морфоструктура востока Азии. – М.: Наука, 1986. –175 с.
37. Куренцова Г.Э. Очерк растительности Еврейской автономной области. – Владивосток: Дальиздат, 1967. – 63 с.
38. Куренцова Г.Э. Растительность Приморского края. – Владивосток: Дальневост. кн. изд-во, 1968.
39. Куренцова Г.Э. Реликтовые растения Приморья. – Л.: Наука, 1968. –72 с.
40. Маак Р. Путешествие по долине реки Уссури. Сиб., 1861. – Т. 1. – 203 с.

