Мониторинг и биоразнообразие экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2012. – 140 с.
Список авторов публикаций. С. 139. Предисловие C.6-8.
Галанин А.В., Галанина И.А. Динамическая парадигма в геоботанике и ботанической географии. С. 9-24.
Урусов В.М., Петропавловский Б.С., Варченко Л.И. К корректировке ботанико-географического и флористического районирования Дальнего Востока России С. 25-45.
Сухомлинов Н.Р. Мониторинг пирогенной трансформации экосистем: проблемы, методы, подходы. С. 46-52.
Игуменова О.П. Экологический мониторинг юго-западной территории республики Башкортостан. С. 53-60.
Киреева Н.А., Якупова А.Б., Григориади А.С. Интегральная оценка устойчивости почвы к нефтяному загрязнению. С. 61-66.
Сухомлинова В.В. Изменение межвидовых стратегических взаимодействий при пирогенной трансформации экосистем. С. 67-74.
Галанин А.В., Долгалева Л.М. К вопросу о применении методики ранжированных невзвешенных вариационных рядов для анализа древостоев на постоянных пробных площадях в Сохондинском биосферном заповеднике. С. 75-92.
Галанин А.В., Галанина И.А. Корейско-Хасанская ботанико-географическая подобласть (очерк растительности). С. 93-114.
Миронова Л.Н., Реут А.А. Итоги репродукции редких и исчезающих видов декоративных травянистых растений в республике Башкортостан. С. 114-118.
Энхтайван Н. Устойчивое лесоуправление в северной Монголии. С. 119-123.
Урусов В.Н., Варченко Л.И. Врищ Д.Л., Петропавловский Б.С. Пути улучшения озеленения Владивостока. С. 123-138.
|

|
УДК 58.00 ББК 72 я 43 М77
Рецензент Христофорова Н.К., д.б.н., проф. ДВФУ
Мониторинг и биоразнообразие экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2012. – 140 с. ISBN 978-5-89694-136-1
Редакционная коллегия: Долгалева Л.М., к.б.н.; Беликович А.В., д.б.н.; Галанин А.В., д.б.н., проф; Петропавловский Б.С., д.б.н.
В сборнике представлены результаты научно-исследовательской и практической работы ученых, преподавателей, аспирантов Дальневосточного региона, в которых отражены глобальные изменения климата и трансформация экосистем, флора, биология и экология растений Сибири и Дальнего Востока, экология природных сообществ, информационные технологии и системы в исследовании экосистем, экологические проблемы сохранения биоразнообразия природных систем Сибири и Дальнего Востока, проблемы охраны и рационального использования животного мира Сибири и Дальнего Востока.
Предназначен для студентов, аспирантов, ученых, специалистов и профессорско-преподавательского состава колледжей и университетов.
© НОУ ВПО «Институт технологии и бизнеса», 2012
|
УДК 57.033
ИЗМЕНЕНИЯ МЕЖВИДОВЫХ СТРАТЕГИЧЕСКИХ ВЗАИМОДЕЙСТВИЙ ПРИ ПИРОГЕННОЙ ТРАНСФОРМАЦИИ ЭКОСИСТЕМ
В.В. Сухомлинова
V.V. Sukhomlinova. CHANGES IN INTERSPECIFIC STRATEGIC INTERACTIONS AT PYROGENIC ECOSYSTEM TRANSFORMATION.
The author proved that fire changes both special composition of plant community and interspecific strategic interactions that helps changed community to survive in conditions of regular fires and to form its stable state which hampers or prevents demutation. The author concluded that even at the local level, pyrogenic factor always has a character of biosphere destroyer. This conclusion contradicts modern trend in the attitude towards fire as a phenomenon helpful for nature.
Tabl. 1. Bibl. 5.
В экосистеме системообразующую функцию выполняет вся совокупность живых организмов, которые занимают нишу в соответствии со своими возможностями к адаптации и использованию ресурсов. Эти закрепленные в геноме возможности формируют стандарт адаптаций вида. При этом, под адаптациями подразумеваются возможности жить в определенных условиях природной среды, вступая во взаимодействия с другими видами. Адаптация к природным условиям и взаимодействию с другими видами формирует биоценотические стратегии, под которыми мы понимаем способ выживания популяций растений в экосистемах [3]. Биоценотические стратегии видов это их способность заселять территорию и встраиваться в биоценоз, захватывая свою нишу и выполнять свою постоянную биоценотическую функцию, то есть это путь вида к своей роли в биоценозе. Исходя из этого межвидовые стратегические взаимодействия можно определить как путь вида к своей нише в иерархии видов.




|
В любых сообществах при любой динамике межвидовые стратегические взаимодействия могут делиться на следующие категории.
1. Синхронизаторные.
Сюда относится совокупность видов, синхронизирующих свои усилия по закреплению данного состояния биоценоза или его восстановлению после не радикальной трансформации. К этой категории относятся прежде всего виды-эдификаторы, субэдификаторы, доминанты-асектаторы, иногда краевые виды. Вся совокупность видов, синхронизирующих друг друга, обеспечивает закрытость биоценоза и способствует его устойчивости.
2. Комплементарные.
Комплементарные взаимоотношения видов можно охарактеризовать как слабый вариант синхронизации, определяемый тем, что один вид играет ведущую роль в динамике биоценоза, а другой, комплементарный ему, усиливает его действие, заполняя второстепенные ниши и усиливая конкурентоспособность ведущего вида. Это касается прежде всего отношений доминирующих и второстепенных видов.
3. Антагонистические.
Эти стратегии во взаимоотношениях видов в рамках данного биоценоза влияют на формирование сукцессий, имеющих противоположную направленность. Вектор развития биоценоза в итоге определяется как пропорцией видов-антагонистов, так и обилием одного вида, являющегося антагонистом по отношению ко всем остальным видам, формирующим биоценоз.
Деление видов по категориям межвидовых стратегических взаимодействий основано не столько на присутствии видов в одном и том же биоценозе определенной сукцессионной стадии, сколько на роли, которую они играют в сукцессионном процессе, взаимодействуя друг с другом.
Пирогенный фактор изменяет среду обитания организмов и способствует элиминации или ослаблению биоцетических функций самих организмов. Последнее изменяет пропорции обилия видов в экосистеме и, соответственно, изменяет их биоценотические и межвидовые стратегии. Пожары, так или иначе, влияют на организмы, выполняющие различные биоценотические функции и обладающие различными межвидовыми стратегиями, но наиболее показательными являются организмы, обладающие в данном биоценозе антагонистическими стратегиями, поскольку антогонисты – это виды, которые «тянут» биоценоз в разные стороны и сосуществуют друг с другом в одном биоценозе только потому, что находятся в пропорциях, обеспечивающих равновесие сил.
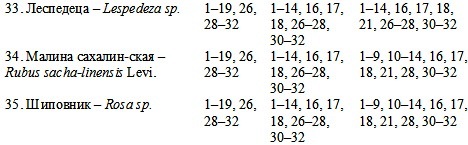
Исследования, проведенные автором в зоне темнохвойно-широколиственного экотона Среднего Приамурья, позволяют проследить изменение межвидовых антогонистических стратегий по мере усиления степени пирогенной трансформации темнохвойно-широколиственных фитоценозов [5] (табл. 1).
В таблице 1 всем индикаторным видам присвоены инвентарные номера, которые проставляются против других видов в том случае, если они являются антагонистами данных видов на данной стадии сукцессионного развития. К естественному климаксу относится такое состояние фитоценозов темнохвойно-широколиственного экотона, при котором в сукцессионно значимый период отсутствуют внешние разрушительные факторы, в том числе и пирогенный. Зональный экотон формируется в условиях поликлимакса и сверхразнообразия [1]. Экотон – это набор различных вариантов климакса с участием определенного комплекса видов, которые «перетекают» из одного фитоценоза в другой, меняя свои биоценотические и межвидовые функции и стратегии. В Среднем Приамурье фитоценотическая мозаика климакса представлена следующими крупными блоками [2].
1. Лиственничники багульниково-бруснично-голубиково-зеленомошные на переувлажненных низинах с многолетней мерзлотой или без нее.
2. Елово-пихтовые леса на крутых склонах (15–60°) преимущественно северной экспозиции с многолетней мерзлотой, сформированной моховой подушкой, на высоте от 500 до 1500 м н.у.м или на менее крутых склонах без моховой подушки и мерзлоты.
3. Преимущественно широколиственные леса с доминированием кедра корейского, дуба монгольского, липы, ясеня маньчжурского и т.п. с участием темнохвойных видов в качестве соэдификаторов, субэдификаторов или проникающих видов.
4. Широколиственные леса, сформированные по поймам рек и прилегающим низкогорьям.
5. Высокогорные сообщества, представленные узкими полосами зарослей кедрового стланника, березы шерстистой и горными тундрами.
Представленная таблица отражает видовой набор фитоценозов среднегорья, где наблюдается максимальное взаимопроникновение видов темнохвойного и широколиственного зонального генезиса.
Под ранним пирогенным омоложением понимается такое состояние экосистемы, при котором под воздействием пирогенного фактора меняются пропорции видов климакса, их биоценотические функции и межвидовые стратегии. Эти изменения позволяют проникать в фитоценоз «чужим» видам, которые начинают выполнять свои функции в обновленном фитоценозе, осваивая стратегии, которые могут реализовываться только в данном варианте фитоценоза. Отличительной особенностью раннего омоложения является обратимость сукцессионного процесса при ослаблении пирогенного фактора.
Под радикальным омоложением понимается формирование фитоценоза в условиях частых пожаров, которые не позволяют проявляться процессам демутации. На этой стадии пирогенной деградации ряд видов зонального климакса из биоты исчезают, их ниши занимают виды, ранее в климаксовые стадии не входившие. Это меняет не только состав видов, но и биоценотические их функции. Поскольку доминирующими видами становятся виды-эксплеренты, то меняется и вся картина межвидовых стратегических взаимодействий.
С этой точки зрения все виды, приведенные в таблице, можно разделить на группу стабилизации и группу динамики. Виды стабилизаторы либо удерживают экосистему на данной сукцессионной стадии, несмотря на воздействие пирогенного фактора, либо закрепляют ее измененное состояние. Фактически это смены эдификаторов, хотя при регулярном пирогенном факторе понятие эдификатора смазывается и отличить эдификатора от доминанта бывает затруднительно.
К видам, влияющим на динамику экосистемы, относятся те виды, которые в силу своих свойств (скорость роста, возможности вегетативного возобновления, общая толерантность, адаптации фотосинтеза и т.п.) либо находятся в положении ассектаторов, либо не входят в состав видов данной сукцессионной стадии, но способны быстро занять освободившиеся ниши. Суть действия видов динамики заключается не только в быстром заполнении освободившихся ниш, но и в ослаблении позиций эдификаторов и доминантов, изменении их биоценотических функций и межвидовых стратегий.
Наиболее ярким примером изменения функций и стратегий в условиях пирогенного фактора являются дуб монгольский и леспедеца, преимущественно двуцветная. В климаксовом состоянии темнохвойно-широколиственного леса эти два вида практически не взаимодействуют. Дуб монгольский выполняет функцию со- или субэдификатора и входит в состава смешанных хвойно-широколиственных фитоценозов. Леспедеца если и входит в состав таких фитоценозов, то только в |
низкогорьях в составе опушечных видов. В фитоценозах раннего пирогенного омоложения позиции темнохвойных видов существенно ослабляются вплоть до временной элиминации. Это осветляет лес и леспедеца проникает в окна и под полог. При ослаблении пирогенного фактора ее вытесняет подрост хвойных растений и леспедеца опять занимает свою нишу краевого вида. Дуб при осветлении получает преимущество и усиливает свое присутствие в фитоценозе за счет подроста. При демутации его позиции опять ослабляются, но он долго еще удерживает свое повышенное обилие за счет молодых особей, вышедших в верхний ярус.
При радикальном омоложении хвойные виды исчезают из состава фитоценоза, широколиственные существенно теряют свои позиции и постепенно выдавливаются эксплерентами. В среднегорье, в условиях комфортного для широколиственных видов влажностно-температурного режима начинает доминировать дуб монгольский, преимуществом которого является быстрый рост на ранних стадиях онтогенеза, высокая способность к вегетативному восстановлению и способность длительно вегетировать при пиротравмах. В осветленные леса с доминированием дуба монгольского проникают кустарники, которые способны быстро отрастать от корня при беглых низовых пожарах. К таким видам относятся, прежде всего, лещина и леспедеца. Причем, леспедеца как бобовое растение обладает большей пиротолерантностью, самостоятельно компенсируя недостаток азота в почве, который возникает при ежегодном выгорании опада. В конце концов, при частых пожарах формируются дубово-леспедецевые фитоценозы, которые можно рассматривать как проявление монополизма двух видов, сменивших антогонистические отношения на комплементарные.
Фактически описанная пирогенная динамика представляет собой катастрофу для экосистем данной природной зоны, поскольку дубово-леспедецевые фитоценозы – это результат элиминации большого количества видов и практическое отсутствие возможности демутации даже при устранении пирогенного фактора. Восстановить хотя бы подобие климакса
теперь возможно только с помощью искусственных мер. Впрочем, активизировать восстановление естественной растительности на месте столь радикально омоложенных экосистем все равно будет затруднительно, поскольку данные сообщества представляют собой подобие климакса. Эти экосистемы обладают своей устойчивостью, которая обеспечивается набором видов с преобладанием стратегий комплементарности. Иными словами, данный биоценоз обладает высоким уровнем закрытости от проникновения видов, не относящихся к стандарту данного биоценоза [4].
Таким образом, пирогенный фактор изменяет не только видовой состав биоценоза, но и межвидовые стратегические взаимодействия, что позволяет измененному биоценозу не только выживать в условиях крайней разрушительности регулярных пожаров, но и формировать свое устойчивое состояние, затрудняющее демутацию или делающее ее невозможной. Все это позволяет сделать еще один вывод – пирогенный фактор даже на локальном уровне всегда носит характер биосферного разрушителя, что несовместимо с распространившимся в последнее время отношением к пожарам как явлению, имеющему свою пользу.
Литература
1. Галанин А.В., Беликович А.В. Зона тихоокеанского муссона: ботанико-географическое районирование, миграции растений и особенности видообразования // Растения в муссонном климате: Материалы V научной конференции «Растения в муссонном климате» (Владивосток, 20-23 октября 2009 г.) Ред. А.В. Беликович. – Владивосток: Дальнаука, 2009. – 353 с., С. 33–43.
2. Колесников Б.Н. Очерк растительности Дальнего Востока. – Хабаровское книжное издательство, 1955. – 104 с.
3. Миркин Б.М., Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов современной фитоценологии. – М.: Наука, 1989. – 223 с.
4. Реймерс Н.Ф. Экология (теория, законы, правила, принципы и гипотезы). – М.: Журнал «Россия Молодая», 1994. – 367 с.
5. Сухомлинов Н.Р., Сухомлинова В.В., Анпилов М.М. Методы изучения пирогенного фактора. // Естественно-географическое образование на Дальнем Востоке: сборник материалов II региональной научно-практической конференции (Благовещенск, 29–30 марта 2010 г.) / под ред. Н.Г. Павлюк. – Благовещенск: Изд-во БГПУ, 2010. – 160 с., с. 79–82.
|

