Типы экофонов
Пролюво-аллювиальный ЛЭ (ПАЛЭ) охватывает поймы рек и речек, отмершие протоки, свежие старицы, днища долинок горных ручьев. Наивысшая континентальность ПАЛЭ среди прочих ЛЭ — следствие закона Воейкова — «На выпуклых формах рельефа местности (на вершинах и на склонах гор и холмов) суточная амплитуда температур воздуха уменьшена в сравнении с равнинной местностью, в вогнутых формах рельефа (в долинках, оврагах и лощинах) увеличена» (Хромов, 1968) вследствие стекания холодного воздуха вечером и ночью (Солнцев, 1960а). При этом резко увеличивается влажность воздуха и нередко можно видеть выпадение росы. Это явление происходит преимущественно в ПАЛЭ. Таким образом, микроклимат ПАЛЭ в значительной мере формируется как функция геоморфологической геосистемы, в которую кроме ПАЛЭ входят и другие ЛЭ.
В эдафическом отношении особенностью ПАЛЭ является резкий градиент проточного увлажнения (от обильного до недостаточного на слабых возвышениях и от периода паводка к низкому уровню воды в реках). Этот градиент является и пространственным, и временным. По механическому составу субстрат делится на галечники, пески, наилки и валунники; имеются небольшие участки с сухими торфянистыми и гумусированными почвами; по отмершим протокам идет процесс заболачивания, который, однако, замедлен, поскольку в половодье протоки функционируют. Диагностическим признаком ПАЛЭ является поемность, а внутренняя его гетерогенность обусловлена особенностями поёмности больших и малых, горных и равнинных водотоков. Необходимо отметить, что хотя ПАЛЭ характеризует поймы, он распространяется только на аллювии и, например, пойменные болота к этому ЛЭ, разумеется, не относятся. Растительность в ПАЛЭ составляют различные луговины и ивняки, изредка рощи деревьев.
ПАЛЭ является каналом, по которому глубоко в тундру проникают бореальные элементы флоры. При этом существует определенная закономерность в соотношении бореальных биоморф, что очень хорошо прослеживается на изученной территории. ПАЛЭ района Телекайской рощи имеет две древесные породы, из них чозения слагает рощу, a Salix udensis встречается единично. Наличие двух деревьев в ПАЛЭ дополняется третьей для района древесной формой, специфичной для КДЛЭ (см. ниже); здесь отмечается большое для Чукотки количество видов кустарников и бореальных травянистых растений.
|

Пойма реки. Пролюво-аллювиальный ЛЭ. Ивняки и редкотравные луга на галечниковых пляжах. Фото А.В. Галанина.
|
В районах, где развитие кустарников подавлено, они встречаются только в ПАЛЭ вместе с травянистыми бореальными видами, которые проникают вдоль рек почти до северного побережья. Наличие кустарниковой растительности обычно обусловлено грунтовыми условиями, а размещение их в долине, в зимнее время забиваемой снегом, обеспечивает их сохранность. В зависимости от различий пойменного режима при удалении от речного русла часто могут быть прослежены специфические полосы растительности, по которым можно составить представление об этапах заселения растительностью затухающих аллювиев. Б. А. Тихомиров (1945) даже считал, что эколого-ценотический ряд, построенный на режимах местообитаний, служит и схемой сукцессионных смен растительных ассоциаций. Однако это происходит далеко не всегда. Значительно чаще сукцессии не детерминированы жестко, а когда они происходят под влиянием экзогенных факторов (что, по всей вероятности, более распространено в природе, чем под влиянием эндогенных факторов), то растительность может перестраиваться по нескольким путям (Кожевников, 19806). Представление о временных сменах растительности, основанное на ее пространственном размещении, может быть только самым общим, не подтверждающимся во многих конкретных ситуациях.
В условиях ПАЛЭ нередко формируется растительность с сильными ценотическими связями и, соответственно, ценотическим отбором. Характерными видами чукотского ПАЛЭ являются: Сагех eleusinoides, Rumex graminifolius, Ranunculus sulphureus, Antennaria villifera, Artemisia borealis, Poa pratensis, |
Salix boganidensis, Aster sibiricus и др., многие из которых достигают на Чукотке северо-восточного предела распространения. Преимущественно для ПАЛЭ характерны: Salix alaxensis, S . krylovii, S. saxatilis, Arabis media, Pedicularis verticillata, Merckia physodes, Festuca cryophila, Artemisia borealis и др. Для любого местного ПАЛЭ очень характерны виды с горной экологией, в том числе редкие в данном районе.
В субконтинентальной полосе и океанических районах ПАЛЭ, по крайней мере некоторых речек, возможно имеет менее континентальный режим среды, чем СЛЭ и КДЛЭ (см. ниже), вследствие длительного залеживания огромных прибрежных снежников, таяние которых уменьшает колебания температур всю первую половину лета. Однако в этих же районах заросли высоких ивняков встречаются уже только в поймах, указывая тем самым на максимум местной континентальности климата именно здесь. Нередко ивняки располагаются вплотную со снежниками, а иногда частично скрыты ими даже в середине июля.
Скальный ЛЭ (СЛЭ). По местоположению этот ЛЭ охватывает всевозможные скалистые уступы (по бортам долин, останцы и пр.). Как режим среды, так и флористическое богатство СЛЭ связаны с величиной отдельных скальных массивов. На небольших по протяженности скалах находится немного видов, особенно специфических, тогда как на крупных массивах скал обнаруживается до 25% выявленной в районе флоры.
СЛЭ характеризуется резкими градиентами среды в сочетании уступов и осыпей (в общем специфическом мезоклимате скал). До определенного размера уступов и осыпей микроклиматические изменения таковы, что растения селятся с равной вероятностью и на тех, и на других. Однако осыпи представляют собой неустойчивый элемент микрорельефа, тогда как уступы более долговечны. В том многочисленном ассортименте микроусловий, которые существуют на скалах, могут сохраняться многие виды, не встречающиеся с некоторых пор в зональных группировках.
Состав горных пород, слагающих скалы, имеет огромное значение и как фактор минерального питания растений, и как фактор, от которого зависит характер эрозии, что связано с накоплением мелкозема в скалах, а также их расчлененности; от последней зависит разнообразие микроусловий обитания. Скалы, образованые кислыми породами, разрушаются медленно, поэтому они обычно слабо расчленены, а образующийся мелкозем выносится за пределы скального массива. Горные породы среднего и основного состава разрушаются быстрее, в результате образуется множество элементов скального нанорельефа и обилие мелкозема, который не успевает выноситься полностью.
СЛЭ имеет хорошую ветрозащиту, так как сила ветра гасится на выступах, возникает турбулентность струй воздуха, которая в зимнее время способствует накоплению снега в скалах, что обеспечивает надежное зимнее укрытие и хорошее водное питание при стаивании снега. Однако таяние снега в скалах происходит весьма быстро, поскольку температурный режим здесь в целом для района особо благоприятный. Лучи солнца падают на некоторые участки перпендикулярно поверхности (при этом под каким бы углом не стояло солнце, оно оказывает максимальный тепловой эффект именно на скалы, так как среди них находятся поверхности, перпендикулярные к любому положению солнца, даже если экспонированы строго на север; об этом можно судить по подтаиванию огромных снежников, забивающих долины, северный склон которых скалистый, а южный — пологий). Такой снежник протаивает у скалистого (северного) склона во всю свою 3—5—7-метровую толщу и не стаивает близ южного (нескалистого) склона в долину. Между снежником и скалами образуется своеобразный коридор. Снежник как бы отодвигается от скал. Прогрев действует не только на нагреваемый участок, но складывается с теплом других участков и в результате создается микро- и мезоклимат скал в целом. Поэтому в летнее время скалы большей частью сухи, если в них нет сочения грунтовых вод. Сказанное объясняет, почему целый ряд ксерофитов на Чукотке, добравшись до своего географического предела здесь, «перебирается» со склонов рыхлой четвертичной толщи (КДЛЭ) на скалы в тех районах, где останцы этой толщи распространены очень широко, но макроклимат изменяется от континентального к океаническому. Однако, если скалы прорезает ложбина, то по ней сверху обычно скатываются холодные воздушные массы, которые задерживают стаивание снежников, лежащих в этой ложбине.
Набор видов и растительность скал зависят от степени их расчлененности и от характера горных пород. На крупных, сильно расчлененных скальных массивах, образованных породами основного состава, набор видов очень богат, а растительность включает фрагменты различных луговин, горных и кустарничковых тундр, но кустарники почти не встречаются, поскольку в зимнее время на скалах не образуется мощный снеговой покров, способный укрыть их. Отчетливое тяготение к СЛЭ обнаруживают: Saxifraga tenuis, Campanula uniflora, Woodsia glabella, Cystopteris dickieana, Poa glauca и др. В разных районах флористическое богатство СЛЭ непостоянно, что в немалой степени отражает макроклимат района. Так, в резко континентальных районах СЛЭ неоригинален флористически и вместе с тем беден по набору видов. Это объясняется и кислыми горными породами, образующими скалы в этих районах, и спецификой мезоклимата скальных массивов, заключающейся в иссушении скал. Об этом свидетельствует, в частности, тот факт, что в резко континентальных районах крайне редко встречаются скалы, отдельные участки которых покрыты моховым ковром.
Коллювиально-делювиальный ЛЭ (КДЛЭ). Определяемый по характеру поверхностных отложений, КДЛЭ охватывает отложения, образованные очень слабо разрушающимся каменистым материалом склонов, перемещающимся в основном за счет гравитации (коллювий), а также делювием — материалом, заметно разрушающимся при движении по склону, обусловленном гравитацией, сносом, солифлюкцией и др. причинами. Эти отложения отличаются, в частности, количеством мелкозема, что создает неодинаковые условия для развития корневых систем.
Коллювий содержит очень мало мелкозема, который большей частью уходит из сферы обитания растений, проваливаясь под крупные обломки горных пород. К коллювию относятся также рыхлые четвертичные отложения склонов холмов, бугров, надпойменных и надозерных террас и расчлененных шлейфов гор (морены, водно-ледниковые и морские отложения). И горные склоны, и склоны холмов никогда не бывают образованы с поверхности только коллювием или только делювием, но всегда имеется сочетание этих двух типов отложений приблизительно в равном соотношении или с доминированием какого-либо одного типа. Раздельное рассмотрение коллювия и делювия по доминированию того или другого является скорее геологической задачей, чем экологической, т. к. среди россыпей коллювия всегда обнаруживаются «карманы» или очень мелкие скопления делювия, на котором и поселяются растения. Эти скопления, по всей вероятности, отчасти являются элювием, но последний не входит в число характерных отложений склонов.

Чукотский п-в. Коллювиально-делювиальный ЛЭ в Провиденских горах. Фото А.А. Галанина. |
По местоположению КДЛЭ всегда представлен склонами, т. к. подвижность каменистого субстрата является одной из основных особенностей физического режима среды этого ЛЭ. По степени каменистости выделяются крупнокаменистые, щебнистые, дресвяные и мелкоземистые участки; на склонах морен — валунчиковые и песчано-галечниковые. КДЛЭ характеризуется режимом среды, в большинстве случаев полностью определяемым ее абиотической составляющей в условиях гор тундровой зоны, т. е. безлесных гор.
Местообитания, относимые к КДЛЭ, обычно сухи, нередко подвержены действию сильных ветров, покрытие растительности в пределах контура фитоценоза колеблется от 0 до 80%, обычно 30—40%. Моховая синузия нехарактерна, но лишайниковая вполне обычна. В целом растительность КДЛЭ чрезвычайно разнообразна. В этих условиях наиболее обычны различные горные тундры, реже на щебнистых субстратах встречаются ивняки. Покрытие снегом зимой может быть различным, но, как правило, оно незначительно. Многие участки склонов гор остаются зимой обнаженными из-за сдувания снега сильными ветрами. В летнее, время прямое влияние сильного ветра не оказывает угнетающего влияния на растительность, если ветер не лобовой. Уже небольшого угла достаточно, чтобы у поверхности земли происходило скрадывание, ослабление силы ветра. В мелких ложбинах, расположенных перпендикулярно направлению постоянных сильных ветров, нередко существует пышная растительность, т. е. иссушающее влияние ветра сведено к минимуму. Это находит отражение и в разнице температур в 2 раза на поверхности почвы и уже на выc.е. 0,5 м. |
КДЛЭ, так же как и ПАЛЭ, является каналом, по которому далеко на северовосток проникает древесная биоморфа (Betula platyphylla ssp . minutifolia), и в котором происходит «затухание» роли кустарниковой биоморфы по мере усиления океаничности климата. Это особенно наглядно прослеживается в поведении Alnus fruticosa (Кожевников, 19736). Структура КДЛЭ, т. е. та совокупность местообитаний, которая его составляет, является показателем степени континентальности - океаничности макроклимата района. В континентальных районах КДЛЭ более гетерогенен, чем в океанических за счет ксеротермных местообитаний. Согласно Т. В. Бехеру (Bocher, 1949), на аналогичных местообитаниях в юго-западной Гренландии суточные амплитуды температур поверхности грунта достигают 50° даже в августе (очевидно, в июле они еще выше).
В субконтинентальной полосе гетерогенность КДЛЭ значительно уменьшается. Суточные амплитуды температур, по нашим данным, не превышают 35°. На ксеротермных местообитаниях существует остепненная растительность, среди которой встречаются, как большая редкость, фрагменты каменистых степей. Сообщества этих степей характеризуются значительной сомкнутостью растительного покрова, достигающей покрытия 80%. Однако большая часть ксеротермных местообитаний имеет разреженную растительность - степоиды.
Кроме ксеротермного варианта КДЛЭ существует карбонатный (на известь содержащих горных породах). И для ксеротермного, и для карбонатного фрагментов КДЛЭ характерны специфические флористические и экологические черты, однако они могут быть представлены как модификации КДЛЭ или КЭЛЭ, а не как самостоятельные ЛЭ. Кроме того, если карбонатный вариант КДЛЭ (а также и КЭЛЭ, см. ниже) выделяется надежно по абиотическому признаку, то ксеротермный вариант определяется скорее флористически. Уже было сказано, что такой подход на Чукотке к интерпретации среды ненадежен. Данные по ландшафтным процессам в якутских степях (Юрцев, 1981) не могут переноситься на Чукотку из-за климатических различий.
Поскольку Чукотка - это территория преимущественно гористая, КДЛЭ представлен на ней очень широко в различных модификациях по интегральному режиму среды и по различию отдельных ее факторов. Растительность КДЛЭ представляют, прежде всего, разности дриадовых тундр; обычны и полидоминантные разреженные тундры, а также куртинные лезлериевые, шикшевые и арктоусовые тундры; характерной растительностью являются также сильно разреженные группировки трав и кустарников. Следует отметить, что куртинные дриадовые тундры, относимые по эдификатору и по физиономической характеристике к одному типу, могут сильно различаться флористически. Из числа наиболее характерных для КДЛЭ видов назовем: Senecio resedifolius, Ermania parryoides, Silene stenophylla, Astragalus tugarinovii, Crepis nana, Arnica frigida и др. Для Центральной Чукотки С. А. Баландин (1978а, б) дал характеристику сухих щебнистых тундр, многие из которых относятся к КДЛЭ, но часть — к следующему ЛЭ. Он отметил относительное возрастание в наборе видов этих тундр берингийских, сибирских и восточносибирских аркто-альпийских видов, а также подушковидных биоморф.
Коллювиально-элювиальный ЛЭ (КЭЛЭ) объединяет местообитания с горизонтальной и полого-наклонной каменисто-щебнистой поверхностью; плоские вершины сопок, незаболоченные нагорные террасы, щебнистые поверхности возвышений (холмов, бугров и плато) и таликов в межгорных долинах. Основные факторы среды: сильная обдуваемость в любое время года (зимой слабое укрытие снегом или оголенность); сухость; преобладание физического выветривания над химическим, поэтому почвы, как правило, не выражены или представлены примитивными скелетными разностями. Растительный покров КЭЛЭ часто сильно разрежен (растительность теряется из виду на близком расстоянии), но в незначительных понижениях проективное покрытие может достигать 70%, здесь же обнаруживается гумусированность щебнистых почв.
|

На переднем и среднем планах коллювиально-элювиальный ЛЭ, на заднем скальный ЛЭ и коллювиально-делювиальный ЛЭ. Фото А.А. Галанина.
|
На примере местоположений КЭЛЭ, обычно имеющих слабо выраженный микрорельеф, можно видеть особенно ясно влияние субстрата на режим факторов среды и ее холоценотическое действие на растения. На элювии щебнистых возвышений в межгорных впадинах набор видов и экологическая обстановка мало отличаются от таковых высоких нагорных террас или плоских щебнистых вершин сопок. Вместе с тем суточные циклы отдельных факторов среды, так же, как и их сезонная динамика, будут заметно отличаться на нижнем и верхнем элювиях.
КЭЛЭ наиболее гомогенен по разнообразию условий обитания среди других ЛЭ, т. е. различные местообитания, относимые к КЭЛЭ, меньше отличаются друг от друга по режиму среды, чем местообитания других ЛЭ. Это объясняется, в частности, тем, что все местообитания КЭЛЭ имеют практически один и тот же нагрев солнечными лучами. При этом, хотя в отдельных случаях поверхность местообитаний КЭЛЭ может нагреваться весьма значительно (до 42°), сумма эффективных температур местообитаний КЭЛЭ ниже, чем местообитаний КДЛЭ, так как лучи солнца никогда не падают перпендикулярно на их поверхность.
Кроме того невелирующим фактором местообитаний КЭЛЭ является выщелачивание поверхностных слоев грунта, в результате чего создаются олиготрофные условия. Почвы на горизонтальных участках существенно отличаются от почв в том же климате и на тех же самых породах, но образующих наклонные поверхности (Dauhenmire, 1947). Однако растительность и набор видов КЭЛЭ отличаются от растительности и набора видов КДЛЭ только обедненностью. По мнению П. Дансеро (Dansereau, 1957), почвы с |
незаконченной эволюцией (incomplete evolution) являются интразональными, а неразвитые и очень молодые (или скелетные) почвы — азональными. В отношении к горным тундрам едва ли можно утверждать, что их почвы азональные, поскольку они сплошь почти скелетные и будут оставаться такими, пока не изменится экологический режим, что может быть результатом неотектонических движений или исторической смены климата.
Р. Бешел (1969) правильно отметил, что, хотя многие виды обязаны своим широким распространением свойственной им значительной экологической выносливости, очень большое число растений может пристутствовать в различных макроклиматических районах Арктики только потому, что микроклиматические условия каждого района сильно варьируют на небольших расстояниях. Однако вариабельность эта связана с поверхностными отложениями, т. е. вариабельность на делювиях отличается от вариабельности на элювиях, на торфяной поверхности и т. д. Именно поэтому каждый тип отложений несет в значительной мере специфичное флористическое обличие и, чем большую площадь этот тип занимает, тем разностороннее его собственная вариабельность и богаче фракция местной флоры, связанная с этими отложениями. Эта фракция была названа «экологическим элементом, комплементарным какому либо ЛЭ» (Кожевников, 1979ж). Не учитывая это обстоятельство, Р. Бешел открыл «бесплодный клин» в Канадском архипелаге, который, якобы, проглядел Э. Хультен. Он пишет: «В то время как многие виды заходят далеко к северу в горных областях островов Элсмира и Аксель Хейберг, к юго-востоку от моря Бофорта простирается обширный «бесплодный клин» («barren wedge») — зона, особенно сильно обедненная видами» (Бешел, 1969). Причина этой обедненности экологическая, а не географическая, на что указал сам Р. Бешел: «Причиной слабого количественного и качественного развития растений и видового однообразия в пределах бесплодного клина, возможно, являются прежде всего свойства субстрата и топография, и во вторую очередь климат. Острова, располагающиеся в пределах бесплодного клина, характеризуются преимущественно карбонатными равнинами и плато, где морозное вспучивание подняло валуны и гальку на поверхность, тогда как мелкозем скрыт в глубине, часто под таликом. Территория бесплодного клина не имеет гор, тогда как прилегающие к ней области с низкими горами на западе (например, на о-ве Мелвилл) и в особенности почти альпийские ландшафты центра о-ва Аксель Хейберга и севера о-ва Элсмира на востоке создают большое разнообразие микро- и мезоклиматов» (Бешел, 1969). Таким образом, бедность «бесплодного клина» обусловлена тем, что на его территории существует почти сплошной КЭЛЭ, экологический элемент которого в любой части Арктики отличается бедностью по сравнению с другими ЛЭ. Согласно Б. Б. Полынову (1956), различается неоэлювий (элювий наносов) и ортоэлювлй (остаточдые продукты выветривания пород in situ ). По экологическому режиму КЭЛЭ существенно отличается от КДЛЭ, но, как будет показано ниже, флористически эти ЛЭ иногда очень близки. Особенно характерные виды КЭЛЭ: Salix phlebophylla, Pedicularis kanei ssp. kanei, Artemisia furcata, Potentilla elegans, Minuartia arctica и др.
Гумусный ЛЭ (ГЛЭ) распознается по богатому содержанию гумуса в почве (пусть даже сильно хрящеватой), что сразу заметно и по характеру растительности, покрытие которой достигает максимума, а флористический состав насыщен весьма редкими в районе видами. Местообитания, относимые к этому ЛЭ, обнару живаются на различных гипсометрических уровнях, в разном окружении, на разных элементах рельефа. Особый вариант этого ЛЭ представляют пятнистые тундры (но не пятнистые болота!), включающие две фации: пятна' голого субстрата и растительность, в которую вкраплены эти пятна.
По местоположению ГЛЭ занимает слабо наклоненные поверхности солифлкционных террас и шлейфотеррас, широкие ложбины стока на склонах сопок, отдельных участков шлейфов и плоских седловин, разделяющих верхние части невысоких сопок. Иногда фрагменты ГЛЭ обнаруживаются не в понижениях, а, напротив, на повышениях, на щебнистых склонах, а особенно на небольших уступах. Такие случаи нетипичны и заслуживают специальных экологических исследований, так как в одних случаях на повышениях (взлобках} на склонах сопок с кустарничковой тундрой развиты «пустошные» или лишайниковые группировки растений, в других, как сказано выше, существует обстановка ГЛЭ с весьма пышной растительностью. Последняя выделяется в виде маленького оазиса на фоне серого щебнистого склона.
|

Гумусный ЛЭ переходный к торфяному ЛЭ в Нижне-Анадырской равнине. Фото А.В. Галанина.
|
Грунтовая основа ГЛЭ часто представляет сухой торфянисто- грубогумусный субстрат мощностью 15—20 см, подстилаемый щебнем. Если такой субстрат пропитан влагой, например, после дождя, он может быть принят за ТЛЭ (см. ниже), однако всегда отличается от последнего полным отсутствием глеевого горизонта. Вымываемые соли железа уходят по щебню, подстилающему дернину.Солифлкция (смещение по уклону оттаивающих горизонтов по поверхности мерзлых пород) часто приводит к захоронению дернины, причем, с тем большей скоростью, чем крупнее оплывина, размеры которой зависят от механических свойств субстрата и его влажности (Kaiser, 1980). На поверх ности оплывины растительность и почвенный слой обычно сохра няются ненарушенными или нарушенными лишь сетью трещин.
А. П. Шенников (1938) выделял среди луговых классов формаций торфянистые луга. На Чукотке настоящие луга замещены луговинами, т. к. среди травянистых мезофитов всегда есть большая или меньшая примесь кустарничков. Однако экологическим аналогом торфянистых лугов являются торфянистые луговины. Некоторые низкие кустарники (Spiraea stevenii, Dasiphora fruticosa), а также кустарничек Salix chamissonis, определенно тяготеют к ГЛЭ, при этом Salix chamissonis становится из кустарничка низким кустарником. На примере отдельных местообитаний ГЛЭ можно видеть насколько характер растительности и набор видов связаны с особенностями поверхностных отложений, независимо от их местоположения. Конечно, в этом видное |
участие принимает и сама растительность, как основа гумусообразования, но определяющую роль для развития растительности имеют абиотические факторы: местное накопление мелкозема, вследствие его легкого вноса и затруднительного выноса; морозное вспучивание грунта, приводящее к образованию голых пятен суглинка и, вместе с тем, являющееся фактором обогащения почв элементами минерального питания вследствие перемешивания; оптимальные условия увлажнения из-за влагоемкости почв и водосборных свойств местоположений; большей частью хорошее снеговое укрытие зимой и слабая обдуваемость ветрами летом (так как сила ветров сказывается пагубно лишь в случае лобовых ветров, боковые ветры в значительной степени гасятся близ поверхности земли).
ГЛЭ нередко является переходным к другим ЛЭ. Особенно распространен случай, когда на древнем элювии (в прошлом -- ПАЛЭ) в настоящее время развит ГЛЭ. Это низкие (1—2 м) надпойменные незаливаемые террасы со смешанно кустарничковыми сомкнутыми тундрами; основу покрова образуют: Dryas punctata, Empetrum nigrum, Vaccinium uliginosum, V . vitis - idaea, Salix pulchra, Cassiope tetragona, Betula exilis, Diapensia obovata, Polygonum tripterocarpum, Hierochloe alpina. В условиях ГЛЭ развивается следующая растительность: различные мезоморфные луговины, кустарничковые полидоминантные или дриадовые, шикшевые, диапенсиевые тундры, пятнистые («медальонные») кустарничково-моховые эутрофные разности тундр, а также ивняки из Salix krylovii, S. lanata ssp. richardsonii. К ГЛЭ следует, по-видимому, относить и сусликовины, охарактеризованные Б. А. Тихомировым (1959, 1960). Относительно верные виды: Saussurea tilesii, Pedicularis oederi, Salix reticulata, Aconitum delphinifolium, Oxytropis maydelliana, Viola epipsila ssp. repens, Gentiana glauca и др. Видовое богатство ГЛЭ тесно связано с богатством всей конкретной флоры.
Озерный ЛЭ (ОЛЭ) имеет с ПАЛЭ то общее, что озерный аллювий столь же неоднороден, как и речной. Его составляют то озерные наилки, то пески, то галечники, нередко берега озер каменисты. Часто берег озера неоднороден на своем протяжении: каменистые или галечниковые участки сменяются участками с болотной дерниной, последние мы относим к ТЛЭ.
Среди других ЛЭ ОЛЭ обладает наибольшей гетерогенностью, что хорошо заметно во впадине уже одного озера и что говорит о прямом влиянии водной поверхности только на берега озер. Кроме того, озера в одном и том же районе нередко различаются по местоположениям, по гидрологическому режиму, что оказывает влияние на экологию их берегов и прибрежный растительный покров. К ОЛЭ относятся почти целиком днища аласов (незаторфованные участки), встречающиеся в южных районах Чукотки. Днища эти покрыты озерными аллювиями мигрирующих в пределах аласа водоемов, с развитым мощным травостоем. Аласы стали уже угодьями по заготовке сена, преимущественно из Arctophila fulva.
Увлажнение в условиях ОЛЭ варьирует от постоянного избыточного до периодически недостаточного. Растительность включает гидро- и мезоморфное редкотравье, луговины, пушичники из Eriophorum scheuchzeri, заросли Senecio congestus и т. д. Флористически весь исследованный ОЛЭ почти лишен специфики, однако именно в «полосе ОЛЭ» происходит затухание или исчезновение некоторых «западных» видов (Utricularia minor, Sparganium hyperboreum). Характерными для ОЛЭ являются заросли Arctophila fulva, Hippuris vulgaris и др. Часто ОЛЭ имеет переходный к ТЛЭ характер.
Торфяной ЛЭ (ТЛЭ) занимает большую часть днищ межгорных долин с осочниками, кочкарниками, бугристыми и валиковыми болотами, а также заболоченные шлейфы сопок, седловины, участки нагорных террас. Основные факторы среды: застойное или полузастойное переувлажнение, низкое рН, торфяные почвы. ТЛЭ всегда выделяется как заболоченное пространство со сплошной дерниной, чаще всего имеющей в основе зеленые мхи. Торф образуется и на щелочных почвах, и в таких случаях он содержит свободные карбонаты, а его рН часто составляет 7 (Coombe, White, 1951). Набор видов ТЛЭ в таких случаях обогащен кальцефитами и базифилами.
|

Кочкарная тундра после пожара. Торфяной ЛЭ. Фото А.В. Галанина.
|
Велика средообразующая роль болотной дернины, образование которой влечет поднятие вечной мерзлоты (залегающей здесь обычно на глубине 20—25(30) см); вслед за этим начинается криогенное вспучивание (образование бугров или валиков, несколько разнообразящих экологию болот). Торфяная дернина оказывает огромное влияние на тепловой баланс, понижая температуру на своей поверхности (много тепла тратится на испарение) и уменьшая ее суточную амплитуду. Этот процесс, очевидно, мало связан с характером общего климата во всех районах Чукотки, вследствие чего ТЛЭ в разных пунктах является неизменным. Еще Ю. Д. Цинзерлинг (1938) отмечал огромную роль микроклимата болот, который нередко сходен даже в разных природных зонах, обусловливая сохранность ряда болотных видов реликтов холодных эпох.
Торфяная дернина представляет также в высокой степени специфический эдатоп. В. Б. Сочава (1956) отметил, что поселение куртин лишайников на моховом покрове представляет в тундре обычное явление, однако это явление характерно лишь для южных тундр. На нашей территории поселение лишайников на моховом покрове наблюдается в основном в резко континентальных районах, но не представляет собой массового явления. Гидро- и гигроморфная растительность ТЛЭ представляет варианты олиготрофных тундр. Некоторые авторы не относят к болотам кочкарные сообщества из Carex lugens и Eriophorum vaginatum, полагая, что их нужно рассматривать как «кочкарную заболоченную тундру» (Боч, Мазинг, 1979. С. 126). |
Кочкарники из названных видов действительно иногда имеют слабо выраженный торфяный слой, очевидно, представляя ранние стадии заболачивания. Но как правило, в развитых кочкарниках слой торфа существует и режим среды направлен на его накопление, поэтому такие сообщества мы относим к ТЛЭ, тем более, что в этом нет противоречия с общими свойствами болотных экосистем согласно М. С. Боч и В. В. Мазингу (1979). Б. Н. Городков (1938) считал, что по строению и составу кочкарники занимают срединное положение между болотами и тундрами. Они могут быть с равным правом отнесены и к тому и к другому типу растительности. Он предпочитал причислить эту широко распространенную в южной половине Арктики формацию к тундрам лишь потому, что кочкарники являются одной из позднейших стадий в трансформации третичных болот и болотистых лесов в тундры, и потому, что в них не происходит стойкого накопления торфа, непрерывно разрушаемого при процессах выпячивания мине рального грунта. Однако выпячивание грунта происходит не всегда, а кроме того, оно происходит и в несомненных болотах, из-за чего они не перестают оставаться болотами. В другом месте той же работы Б. Н. Городков писал: «Взамен моховых тундр широко распространяются кочкарные тундры с гипново-сфагновой, торфянистой дерновиной, не позволяющей почве оттаять глубже 50— 100 см». Здесь он отметил наличие торфянистой дерновины. Но его наблюдения о замене кочкарниками моховых тундр основано, видимо, на каких-то специфических районах, потому что по всей Чукотке моховые тундры и кочкарники распространены совместно. Можно думать, конечно, что кочкарники более конкуренто способны, чем моховые тундры, однако они так распределяются в ландшафте, что не мешают друг другу. Моховые тундры занимают часто нижние части склонов гор и шлейфы, тогда как кочкарники из пушицы влагалищной распространены на равнинных или слабо наклоненных участках.
Ранее считалось, что микроклимат на торфяных поверхностях более холодный, чем на минеральных грунтах. Однако было показано, что это ошибочное представление (Ruuhijarvi, 1969). Он оказывается благоприятнее, чем в окрестных лесах. В дневное время температура торфяной поверхности несколько понижена за счет расходования тепла на испарение, а в ночное — повышена на 2—3° за счет нагретой днем воды, содержащейся в торфе. В континентальных районах, однако, при ежедневном нагреве, торфяной пласт сильно высыхает и нагревается так же, как минеральная поверхность. Но в последнем случае тепло распространяется вглубь, тогда как торф выполняет функцию теплоизолятора и не пропускает вглубь тепло, так что мерзлота может располагаться на небольшой глубине, а ледяные клинья почти выходят на поверхность даже в середине лета. Специфические для ТЛЭ виды: Pedicuiaris parviflora ssp. pennelii, Hierochloe pauciflora, Carex vaginata, C. rodundata, Salix fuscescens, Senecio atropurpureus, Eriophorum russeolum, E. vaginatum, Luzula wahlenbergii и др.
Нивальный ЛЭ (НЛЭ) объединяет все участки с залеживанием снега, по крайней мере, до июля. Снежники создают совершенно особый режим среды, отличающийся по многим показателям: температуре, влажности воздуха и влагосодержанию субстрата, который целиком является продуктом нивации (Солнцев, 1949). Расположение снежников и, соответственно, нивальных группировок не зависит от свойств рельефа, в том числе от экспозиции. Долгое лежание снега происходит на различной высоте и является результатом ветрового режима (перераспределение снега зимой). Нередко обширные участки НЛЭ занимают берега озер, поскольку ледовый покров на озере способствует долгому сохранению снежников под склонами в озерную впадину. Имеются также участки нивального ПАЛЭ, где речной аллювий смешан с нивальным наилком под снежником.
Растительность НЛЭ тесно связана с повторяющейся из года в год относительной продолжительностью лежания снега. Можно выделить, по крайней мере, три типа нивальных группировок: кассиоповые (недолгое залеживание снега), ковриковые ивковые из (более продолжительное лежание снега); приснеговые редкотравные (снег стаивает к концу лета, в отдельные годы не успевает стаять). На юго-востоке Чукотского полуострова, в условиях океанического климата установлены две категории сообществ, формирующихся в НЛЭ в зависимости от мощности снега: хионофитные (толща снега до 4 м и более) и гемихионофитные (толща снега от 4 до 0,7 м) (Баландин, Разживин, 1980).
Развитие НЛЭ в районе служит показателем континентальности—океаничности климата. Так, в резко континентальных районах НЛЭ может практически отсутствовать, но в океанических районах, кроме типичного НЛЭ (близ снежников), местообитания с различными оттенками нивальности можно встретить в любом другом ЛЭ. Нивальные виды в океанических районах представлены крупными местными популяциями, и, соответственно, имеют расширенную экологическую амплитуду, часто встречаясь не в собственно нивальных условиях, но зато индицируя малейшее уклонение режима среды в сторону нивальности. Группа нивальных видов является наиболее показательным «фитометром» тенденций климата на любом его уровне. Вместе с тем, в нивальных группировках в большинстве случаев отсутствуют ценотические отношения (Тихомиров, 1945). Типично нивальные виды: Salix polaris, Carex tripartite, Poa paucispicula, Phippsia algida, Saxifraga hyperborea, Ranunculus pygmaeus, Koenigia islandica и др.
Лайдовый ЛЭ (ЛЛЭ) охватывает приморские осоковые луга и болота на тяжелых суглинках, часто со множеством непересыхающих луж и озер. Так же, как и в ТЛЭ, здесь образуется болотная, но солоноватая дернина, покрывающая древние галечники, которая служит одним из основных элементов режима среды ЛЛЭ. В настоящее время ЛЛЭ не заливается морской водой или заливается крайне редко. Как правило, ЛЛЭ распространяется только на низинные приморские пространства, т. е. связан с характером рельефа так же, как и некоторые другие ЛЭ.
В самом ЛЛЭ происходит образование микрорельефа, поэтому встречаются равнинные, бугристые и валико-озерковые лайды. Толща соленого торфа иногда достигает 0,5 м и насыщена газами. Участки ЛЛЭ обычно имеют ржавый цвет. Кроме засоления и застойного переувлажнения, в число основных факторов среды входят позднее освобождение от снега, сильные морские ветры, холодные туманы. Наиболее характерные виды: Carex subspathacea, С. glareosa, Calamagrostis deschampsioides, Puccinellia tenella, P. phryganodes и другие гидрогалофиты.
|

Лайды на берегу Бериногова моря. Фото А.В. Галанина.
|
Приморско-галечниковый ЛЭ (ПГЛЭ) индицирует галечники и пески берегов южного и северного побережий. В данном случае совершенно очевидно, что в отношении отдельных факторов среды различия побережий будут велики, но столь ли велико будет различие холоценотического действия среды на живое? По визуальным оценкам существующая разница экологических режимов разных побережий не превышает гетерогенности других ЛЭ, так же, как и гетерогенности всего ПГЛЭ в пределах одного побережья (северного или южного). Субстрат ПГЛЭ постоянно обновляется под воздействием прибоя и приливов, и поступания нового материала. Арктические берега подвергаются интенсивному разрушению во время штормов. Материал от разрушения формирует аккумулятивные прибрежные формы, возраст которых от десятков до первых сотен лет (Harry et al., 1983). Во фьорде Айси-Бей в зал. Аляска после отступания ледника к 1913 г. начала активно расти коса за счет вдольберегового переноса материала (Molnia, 1977). К 1922 г. она имела длину 3,18 км, в 1976 г. — 6,6 км (средняя скорость роста 92 м/год). За период 1922—1971 гг. коса получила много наносов. Если скорость ее роста сохранится, она через 20 лет полностью перекроет устье фьорда. Материал к косе поступает за счет интенсивного размыва берегов зал. Аляска восточнее фьорда. Сравнительный анализ карт и аэрофотоснимков разных лет показывает, что с 1922 г. берег здесь отступил на 1,5 км, с 1941 г.— на 1,3 км (средняя скорость отсупания 37 м/год). Западнее устья фьорда отступание берега с 1922 г. достигло 4,8 км, а в период 1922—1976 гг. потеря суши составила здесь более 8,2 км. Средняя скорость отступания абразионных берегов моря Лаптевых 3—7 м в год, при |
этом более интенсивно отступают берега с невысоким (8—12 м) клифом, образованным аласными отложениями, а скорость отступания клифов выс. 20—30 м, образованных сильнольдистыми верхне-плейстоценовыми алевритами, составляет 3,5—5 м в год (Новиков, 1981).
|

Приморский злаковый луг. Фото А.В. Галанина.
|
К сожалению, по Чукотке подобных данных не имеется, но некоторые косвенные показатели позволяют судить о грандиозности процесса размыва берегов и выноса материала на производство кос и баров. Так, близ пос. Пинакуль на высокой приморской террасе (порядка 50 м) находится кладбище. Еще в 1970 г. видны были гробы (из захоронений сделанных в 30-е годы) поскольку терраса интенсивно разрушалась морем снизу, а верхние ее части оплывали и отваливались. Ясно, что, когда кладбище закладывалось, край террасы был еще на значительном удалении. Таким образом, огромная масса суглинка, слагающего террасу, оказалась в заливе и была вынесена в его устье вдольбереговым течением. Эти течения разносят и аккумулируют разный материал. Часть его транспортируется на припайных льдинах. Льды способствуют формированию микрорельефа. В верховьях заливов, куда впадают речки, часто образуются наледи, распадающиеся в течение лета. Крупные льдины сохраняются на осушках, долгое время препятствуя размыву покрова рыхлых осадков и в то же время перепахивая его. Иногда льдины создают ложбины выпахивания рытвины глубиной 10— 30 см, а также создавая валунные гряды до 2—3 м высоты (Martini, 1981).
Близ «материкового» края субстрат ПГЛЭ обычно более закреплен, особенно, когда он представлен галечником. Чаще же он представлен песками. Развитие растительности на перевеваемых дюнах смягчает ветровой режим в приземном слое воздуха, что способствует стабилизации растительности. Внешний край полосы ПГЛЭ, как правило, постепенно захватывается ТЛЭ или ГЛЭ, в зависимости от того, какая дернина на него наползает. Комплекс действующих |
факторов ПГЛЭ: рыхлость субстрата и его засоленность, действие нагонных волн и льдин, постоянная высокая влажность и низкая сумма эффективных температур. Вообще в климатическом отношении ПГЛЭ не имеет преимущества перед ЛЛЭ, но существование огромного различия в приземных режимах ЛЛЭ и ПГЛЭ не нуждается в доказательстве. Это — один из ярких примеров средообразующей роли поверхностного субстрата. Флористическую основу ПГЛЭ составляют: Mertensia maritima, Honkenya peploides ssp. diffusa, Dendranthema hultenii, Ley mus villosissimus, Cochlearia arctica и др. Всего в составе приморской флоры Чукотки насчитано 102 вида (Сергиенко, 1983).
О функционировании ЛЭ во времени
На примерах ЛЛЭ и ПГЛЭ особенно наглядно прослеживается влияние субстрата на набор и численность видов, поскольку оба они находятся в одних и тех же мезоклиматических условиях, определяемых влиянием моря. Если сопоставлять только флористический состав, то обнаружится, что оба этих ЛЭ практически не различаются, поскольку хотя бы одиночные особи встречаются на том ЛЭ, для которого они, судя по численности на смежном ЛЭ, не характерны. В этой связи можно отметить различие подходов к оценке представленности видов в определенной растительности. Л. Г. Раменский (1925), никогда не бывавший в Арктике, расценивал количественные отношения массовых растений, как лучший показатель характеристики местообитания, чем наличие или отсутствие единично представленных видов. Другой точки зрения придерживался исследователь растительности Арктики Н. Полунин, считавший, что в целом стенотопные виды являются лучшими индикаторами среды, чем эвритопные виды «по причинам, которые очевидны» (Polunin, 1960). На стенотопных видах, принимаемых за индикаторные, как известно, построена система растительности Браун-Бланке.
Сравнение ЛЛЭ и ПГЛЭ показывает, что режим местообитаний лучше определяют массовые растения. Однако флористический состав способен дать экологическую информацию в других случаях, но значимость в качестве индикаторных видов стенотопных представителей флоры очень невелика и зачастую весьма двусмысленна. Многие стенотопные виды существуют на местообитаниях, относящихся к некоренным фациям, попав сюда случайным образом. Так, Е. В. Дорогостайская (1978) выявила на Чукотке 71 апофит, т. е. растений туземной флоры, поселяющихся на антропогенных, в том числе техногенных, местообитаниях. Среди таких видов названы Elymus mollis, Poa glauca, Arctagrostis latifolia и др. Кроме того, автор привела для Чукотки 85 видов антропохоров, индикаторная роль которых ничтожна. Заносные растения поселяются на Чукотке не только на антропоенных местообитаниях, но могут встречаться и на естественных. В то же время некоторые виды исчезают из состава местных флор, благодаря естественным процессам, в частности, усилению океаничности климата. На одних местообитаниях общее изменение климата сказывается быстро, на других замедленно.
По словам А. А. Крауклиса (1974), «Любой фации присущи, с одной стороны, признаки, обусловленные зонально-региональными (фоновыми) факторами, общими для всего ландшафта, и с другой свойства, определяемые влиянием локальных окрестностей и «саморазвитием» (функционированием) фации. Фоновые факторы и локальные окрестности создают внешнюю среду, а в результате функционирования фации формируется ее внутренняя среда». Эти представления применимы и к ЛЭ, которые составлены рядами фаций. «Внешняя и внутренняя среда,— как полагает далее А. А. Крауклис,— находятся в определенных соотношениях, о которых косвенно можно судить по величинам, характеризующим локальные контрасты и отклонение признаков данной фации от зонально-региональной нормы».
Локальные контрасты каждого ЛЭ могут быть охарактеризованы растительностью, относимой к разным ее типам, хотя, разумеется, могут быть осуществлены инструментальные наблюдения и получены количественные данные, например, с помощью относительных индексов различий (Jackson, Newman, 1967) или по той методике, которую мы применяли на Чукотке (см. выше) и на плато Путорана (Кожевников, 1981). Представление о зонально-региональной норме складывается из высокой частоты встреч определенной растительности на местообитаниях каждого из ЛЭ. Соответственно низкая частота характеризует отклонения от этой нормы. Оценить эти показатели совсем не сложно, поскольку ЛЭ определяются по характеру отложений, т. е. по собственному признаку.
Согласно А. А. Крауклису (цит. соч.) существует три категории фаций: 1) уравновешенная, 2) частично уравновешенная, 3) неуравновешенная, т. е. испытывающая сильное влияние окружения, часто прерывающее процесс саморазвития фации. Частично уравновешенные фации это промежуточные между коренными фациями и сериями. Среди них выделяются мнимокоренные, полукоренные и полусерийные фации. Все это разнообразие складывается в факториально-динамические ряды фаций, которые выражают связи между крайними звеньями (коренные фации — серии). В качестве примеров рядов названы субгидроморфный, сублитоморфный, субкриоморфный (заметна склонность автора выделять ведущий фактор режима фаций). Крауклис различает следующие типы переменных состояний фаций: активизацию, нормализацию, стагнацию (микрорельеф «законосервирован»). Каждый ЛЭ объединяет эти состояния, чем и объясняется его гетерогенность.
|
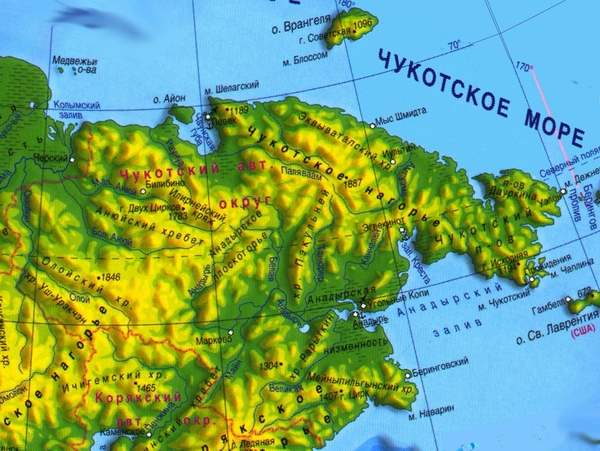
На этой карте Чукотки хорошо видно, как глубоко в глубь материка могло заходить море в эпохи трансгрессий - это территории, закрашенные в зеленый цвет.
|
В силу своей объемности ЛЭ является той ландшафтно-экологичестсой (экотопологической, геосистемной) единицей, которая позволяет исследовать одну и ту же совокупность видов одновременно в историческом и современном экологическом аспектах, поскольку ЛЭ имеют значение миграционных каналов и наиболее гомеостатичных (инвариантных) комплексов среды. Среди ландшафтоведов признано, что в каждом ландшафте имеются реликтовые элементы и прогрессивные, и с изменением одного фактора среды меняется весь их комплекс (принцип Ле Шателье) (Калесник, 1952; Арманд, 1975). При климатических изменениях инвариантность отдельных местообитаний способствует поддержанию интегрального эффекта среды бывшего здесь до наступления изменения. Если же интенсивность изменения перерастает возможности инвариантности отдельных местообитаний, то их среда становится подобной среде других местообитаний того же самого ЛЭ, т. е. вступает в силу инвариантность ЛЭ. При этом на новых местах постепенно складывается набор видов, существовавших до того в других местах. В этом отношении уместно вспомнить слова Л. Г. Раменского: «Устойчивы не группировки, а только законы сочетаемости растений, они и подлежат изучению».
В этом отношении очень показательны случаи как реликтовой, так и пионерной растительности. Примером реликтовой растительности, как бы переползающей с места на место, является растительность, включающая целый ряд арктоальпийских видов, на крутых приречных склонах, сложенных гипсом, в северной тайге Европейской части СССР. Постоянное подмывание рекой обновляет склоны, и аркто-альпийская растительность на них имеется, но как только река отступает в глубь долины, начинается облесение склона, и аркто-альпийская растительность вытесняется. Однако река начинает подмывать другой склон, с которого постепенно исчезает лес, и он столь же постепенно |
покрывается аркто-альпийской растительностью. Этот процесс длится там уже 10000 лет. Аналогично обстоит и с пионерной степеподобной растительностью в северной тайге Якутии. Выгоревший на склонах лес замещается группировками ксерофитов, которые с течением времени вытесняются восстановившимся лесом, но формируются на другом месте, где лес выгорел или исчез по другой причине. Степные участки на месте горелых лесов на Колыме иногда имеют возраст менее 50 лет (дата пожара), а не являются поздне плейстоценовыми.
По всем или большинству ЛЭ, выделяемых в районе, распределяются: а) виды каждого географического элемента флоры, б) редкие виды, в т. ч. находящиеся на пределе своего географического распространения, в) экоморфы, г) биоморфы, д) виды одного рода. Эти распределения не зависят от объема флоры (Кожевников, 1975а, 19786, 19796). Следовательно, каждый групповой признак растений имеет тенденцию «растекаться» по различным комплексам среды обитания. В каждом комплексе поэтому обнаруживается набор видов с различными свойствами, перечисленными выше. Это хорошо заметно и на отдельных местообитаниях, где сосуществуют виды с разными ареалами, экоморфами, биоморфами, принадлежащие к разным родам и имеющие различную численность и встречаемость в целом районе. Существование впределах одного комплекса среды набора видов, каждый из которых отличен от других каким-либо качеством, обусловлено оптимальным использованием растительным покровом материально-энергетических ресурсов данного комплекса.
При исторических изменениях климата и флуктуациях объема местных флор происходит изменение объема всех экологических элементов, но, возможно, с разной скоростью. Массовая миграция растений в новый район (например, после периода оледенения) происходит сразу по всем ЛЭ, хотя по разным ЛЭ может мигрировать разное число видов. Обратный процесс — обеднение вследствие ухудшения климата — также происходит по всем ЛЭ сразу, о чем свидетельствует распределение по ЛЭ редких, подчас исчезающих видов на периферии ареалов в каком-то одном районе (Кожевников, 19786).
Среда обитания находится в постоянном эволюционном процессе. Ее изменения за достаточно длительные промежутки времени (превышающие, по крайней мере, смену нескольких поколений растений, не отличающихся долгожительством) необратимы. Эволюция среды является основным фактором эволюции организмов, особенно растений, которые привязаны к своему местожительству. При этом существенны два момента: темп эволюции и неравнозначность ее проявления в элементах или на разных уровнях, преемственность органических форм развития.
Как уже говорилось, ландшафтные экофоны характеризуются (и определяются) субстратом. Каждый из них обладает гетерогенностью в принятом масштабе определения этого понятия. Так, коллювий-элювий может обнаруживаться на разных гипсометрических уровнях и, в сущности, это касается любого ЛЭ. Естественно, что микроклимат на разных гипсометрических уровнях будет в немалой степени зависеть от факторов макро и мезомасштаба. Однако микропоправка, создаваемая субстратом, может быть большой. Можно утверждать, что, по крайней мере, в ряде случаев именно характер субстрата, а не факторы макромасштаба, определяет околоземную среду и, собственно, растительность. Например, грубогумусная или торфяная поверхность плоских вершин низких гор определяет практически тот же набор видов, что и подобная поверхность в межгорных впадинах. На щебнистых вершинах подобных гор набор видов совсем иной и он соответствует наборам щебнистых поверхностей нижних уровней. В ходе климатических изменений среда на разных уровнях одного ЛЭ подвергается разным перестройкам. Например, при потеплении климата нижние щебнистые поверхности будут задерновываться, но верхние сохранятся еще долгое время; при похолодании климата грубогумусные и торфянистые поверхности верхних уровней будут разрушены и снесены, но уцелеют на нижних уровнях.
Существование и структура каждого ЛЭ отражают современные климатические условия. Все ЛЭ, взятые вместе в их взаимодействии, позволяют говорить не только о современном климате региона, но и о направленности его изменений в ближайшем будущем. Это становится возможным при сравнительном изучении ЛЭ и их экологических элементов в разных в климатическом отношении районах. Эволюция каждого ЛЭ отражает одну из особенностей развития ландшафта. Некоторые ЛЭ являются климатогенными (ГЛЭ, НЛЭ, ГЛЭ), другие — тектоногенными (СЛЭ, отчасти ПАЛЭ), третьи — геоморфогенными (ОЛЭ, разности КДЛЭ и КЭЛЭ). Но многие местообитания даже одного ЛЭ, во-первых, образовались благодаря влиянию нескольких факторов, и во-вторых, главенствующие факторы могут быть разными. Например, условия КЭЛЭ складываются на неоэлювии, т. е. на участках, не так давно вышедших из поемного режима вследствие углубления русла реки или изменения его направления, на участках высоких надпойменных террас, имеющих ледниковое, водноледниковое или мор ское происхождение, на участках нагорных террас, образовавшихся в результате поднятия гор и при осыпании материала со склонов (что также может быть результатом поднятия гор) и т. д. Условия ТЛЭ складываются в бессточных впадинах (где, однако, не накапливается слишком много воды), на зарастающих мелких озерах, на равнинах и в речных долинах, на шлейфах гор, если с выше расположенных участков происходит постоянный подток воды, и на равнинах в связи с поселением пушицы влагалищной или мхов. Очаги заболачивания на Чукотке возникают легко, поскольку часть выпавших осадков не испаряется. Однако исчезают они редко. Заболачивание прекращается, по-видимому, только в связи с поднятием и, соответственно, усилением стока, либо при образовании дренажной ложбины или целой сети их.
В ландшафтоведении рельеф и литология являются ведущими факторами формирования типов местности лишь на определенном, хотя и достаточно широком климатическом фоне (Мильков, 1964). Если в этой формулировке заменить «типы местности» «типами ЛЭ», то она также будет справедлива, поскольку рельеф, литология и макроклимат являются фоновыми элементами существования и степени выраженности ЛЭ. Если, например, в районе нет гор, то отсутствует скальный ЛЭ, а если в районе сплошные горы, то в нем не находится места для торфяного или озерного ЛЭ, а благодаря литологии могут быть выделены крупные подразделения в каком-либо ЛЭ, например, кальциевый или базальтовый. В условиях резкоконтинентального климата на низших гипсометрических уровнях отсутствуют снежники, поэтому процесс нивации (Солнцев, 1949) там не проявляется; соответственно, отсутствует нивальный ЛЭ.
Пытаясь определить возраст ЛЭ, мы неизбежно входим в область палеогеографии. Непрерывное существование наиболее древних ЛЭ могло начаться только после максимального оледенения, поскольку, согласно М. Т. Кирюшиной, «в эпоху максимального оледенения Чукотско-Анадырский край представляется нам в виде обширной горной страны, «затопленной» ледяным морем, над поверхностью которого высились отдельные группы гор или целые кряжи с крутыми, лишенными снега склонами и цирками на вершинах» (по Тюлиной, 1936). Это оледенение происходило в среднем плейстоцене (Обручев, 1939; Колосов, 1947; Сакс, 1948; Петров, 1965, 1976; Petrov, 1967; Hopkins, 1972 и др.).
Во время максимального оледенения климат был чрезвычайно суров, и растительность на Чукотке современных очертаний, по-видимому, была уничтожена, но на осушенном шлейфе происходила миграция резистентных растений и животных, минуя Чукотку (табл. 5). В Восточной Сибири климат был континентальным, и поэтому значительные ледовые покровы там не сформировались (Григорьев, 1946). Этот же автор полагал, что южная граница тундр проходила в то время там же, где и теперь, или лишь немногим южнее. В свете новых данных, полученных в Сибири (Баранова, Бискэ, 1964; Боярская, Малаева, 1967; Гитерман и др., 1968; Попова, 1970, 1973; Равский, 1972 и др.), хорошо видно, что южная граница тундр сдвигалась к югу, хотя ее конкретное местонахождение вряд ли может быть указано с такой определенностью, как в работе Р. Е. Гитерман и др. (1968).
 |
В последней реконструкции суши берингийских районов Д. Хопкинса (1976) показано, что Анадырский залив не осушался в период максимального (иллинойского) оледенения. Этот залив обеспечивал влагой для образования материковых льдов Чукотку и северную Аляску. В пользу этой точки зрения имеется датировка морских отложений на мысе Дионисия близ Анадыря 184±22 тыс. лет (Свиточ, 1976). Существование обширной акватории на юге Берингии свидетельствует о весьма океаническом климате, по крайней мере, в южной ее части, что несовместимо с представлением о здешней тундростепи. Имеются достаточные основания для суждения о том, что в период максимального оледенения на Чукотке современных очертаний могли существовать открытые участки склонов гор (КДЛЭ), но из-за суровых условий они были необитаемы.
Последовавшее за этим оледенением казанцевское межледниковье было теплым. Мост суши (Берингия), по мнению В. Н. Сакса, сохранялся и после деградации максимального оледенения. Но в первую половину последовавшего межледниковья мост суши исчез и наступила трансгрессия, воды которой достигали Марковской впадины. Представление о теплом периоде бореальной трансгрессии оказало влияние на мышление фитогеографов. В частности, Б. А. Тихомиров (1946) писал: «Сравнение растительного покрова арктических районов Восточной Сибири с растительностью современной зоны |
полярных пустынь показывает, что межледниковый (или межледниковые) период и связанная с ним бореальная трансгрессия оказали существенное влияние на развитие расти тельного покрова Арктики».
С казанцевского времени началась эволюция КДЛЭ, КЭЛЭ, ТЛЭ, ГЛЭ. Другими словами, имелись горные склоны, местами задернованные, а также нагорные террасы, тоже местами задернованные (может быть, большей частью) или заболоченные. Возможно, что в глубине горных массивов была развита гидросеть и имелись озера, т. е. существовали также ПАЛЭ и ОЛЭ, но мало вероятно, что были скалы (СЛЭ) и совсем невероятно, что были снежники-перелетки, создающие НЛЭ. Береговая линия, проходившая в глубине современной суши, создавала условия для ЛЛЭ и ПГЛЭ. Трансгрессия происходила из-за низкого стояния суши (в результате гляциозостатической просадки) и из-за повышения уровня Мирового океана. Однако освобождение суши от нагрузки льдов вызвало ее поднятие, которое продолжалось до конца казанцевского этапа.
По мнению В. Н. Сакса (1948), во второй половине казанцевского межледниковья произошло общее воздымание суши. Очевидно, с этим поднятием можно связывать образование скалистых верховий в некоторых фьордах. Нужно сказать, что представления Сакса хорошо соответствуют гляциоизостатической концепции, которую сам он принимал лишь как малозначимый фактор. По-видимому, со времени казанцевского межледниковья только отдельные участки склонов в верхних частях гор существуют непрерывно вплоть до нашего времени, учитывая, разумеется, их физическое обновление. Если такие участки и существуют ныне, то их экофон за весь период менялся неоднократно из-за изменений климата. Менялся также и субстрат вследствие оползания, выветривания и т. п. Несмотря на все сделанные допущения, сами по себе каменистые участки горных склонов, вероятно, представляют наиболее древний тип субстрата, определяющего современные ЛЭ. Таких участков, возможно, было немного, так как последовавшее оледенение (зырянское, ранний вюрм или висконсин) было весьма значительным. Впрочем, имеются указания, что во время зырянского оледенения ледники, хотя и были крупными, не выходили за пределы горных систем (Сакс, 1948). Относительно мощный ледовый покров, возможно, развивался в районе оз. Элыыгытгын, тогда как в низовьях Анадыря имелись лишь локальные ледники. Но некоторые горные системы оледенение охватывало сверху донизу. Так, например, Экугский массив на севере Чукотки был покрыт льдом почти целиком, и конечные морены были вынесены одним из языков ледника далеко в Ванкаремскую низменность. С юго-западной оконечности хр. Искатень языки льда спускались в Анадырскую низменность до среднего течения р. Канчалан, где они также оставили морены. Участки низменности, не покрывавшиеся льдом, по свидетельству ряда авторов (Гитерман и др., 1968; Гасанов, 1969; и др.), не были безжизненными. На них развилась типичная кустарничково-моховая тундра, создавая условия ГЛЭ. Нет никакого сомнения, что в летнее время, когда льды подтаивали, существовали обширные площади, занятые НЛЭ. Причем, вдоль краев ледников НЛЭ тянулся широкой полосой непрерывно, как это имеет место вдоль края современного ледника Гренландии. Анадырская низменность в целом была свободна ото льда и к ней существовали проходы по незанятым льдом пространствам из более глубинных районов от Яны и Колымы. В. Н. Сакс считал, что севернее устьев Лены и Колымы располагался крупный ледовый щит.
Это противоречит палеонтологическим данным А. В. Шера (1971). Сакс не показывает существование берингийского моста в зырянское время, а считает, что он образовался после этого оледенения. По его мнению, мост был значительно уже, чем в эпоху максимального оледенения. Появилась чукотская депрессия, представлявшая крупный залив на осушенном шельфе. Время появления чукотской депрессии — очень важная веха в истории Берингии,'поскольку означает существенное изменение физико-географических условий северной Берингии и возникновение крупной топографической преграды как раз на основном миграционном пути животных и растений. Представления Сакса о существовании Берингии после зырянского оледенения в настоящее время не могут считаться достоверными. Их следует, очевидно, относить ко времени зырянского оледенения.
В это время Анадырский залив осушился и район окрестностей г. Анадыря был удален от побережья почти на 300 км. Между устьями рек Танюрер и Канчалан и в долинах этих рек, а также в междуречье Великой и Туманской на юге Анадырской низменности существуют пояса конечных морен, которые выражены очень четко (Васьковский, 1970). Эти пояса, по всей вероятности, нужно рассматривать как зырянские. Весьма примечательно, что языки зырянских ледников не достигали современных фьордов Чукотки, сильно обработанных языками льдов самаровскрго периода. Во фьордах, особенно в их верховьях, обычно имеются скалистые участки с наиболее благоприятным режимом условий. В зырянское время верховья фьордов находились далеко от морских побережий и, несмотря на суровый в целом климат, их тепловые условия могли быть достаточными для быстрого таяния снега, накапливавшегося зимой. Верховьям фьордов, как убежищу для относительно термофильных видов в эпоху оледенения, придавал большое значение Э. Эбби (Abby, 1936) критически рассмотревший представление М. Фернальда (Fernald, 1925) о роли горных вершин в качестве нунатаков в зал. Св. Лаврентия (п-ов Лабрадор). Свои догадки он подкреплял мнениями геологов. Так, он писал, что геолог Оделл ( Odell ) считал, что нунатаков не существовало, и округлые вершины с массивами останцов на них представляют послеледниковый эффект выветривания. Он сравнивал ситуацию со Шпицбергеном, где ледники были недавно.

Шпицберген. Нунатаки над ледником. Фото взято с сайта Googl eth.
|
К представлению о нунатаках на вершинах гор с сочувствием относился К. Остенфельд (Ostenfeld, 1926), исследуя историю флоры Гренландии. X. Симмонс (Simmons, 1913), рассматривая вопрос о переживании растений на горных нунатаках ледниковой эпохи, указывал, что нам следует искать такие растения только среди высокоарктических наиболее устойчивых видов, но все они являются в основном циркумполярными и поэтому являются плохим свидетельством гипотезы о переживании ими ледниковых периодов на нунатаках. Учитывая все «за» и «против», Э. Эбби приходит к заключению, что присутствие кордильерских видов в зал. Св. Лаврентия не связано с существованием нунатаков, так как если они и существовали, то их флора должна быть подобна таковой северной Гренландии больше современной флоры этого залива. Присутствие кордильерских видов на полуострове Лабрадор требует иного объяснения: либо их послеледниковой миграцией сюда, либо их переживанием в более низких широтах в укрытых фьордах ниже уровня льдов, что весьма соответствует современной ситуации в Гренландии. В качестве «хорошей рабочей гипотезы» Эбби принял концепцию о климатическом оптимуме, во время которого происходили миграции (в том числе, и с Кордильер), а во время послеледникового похолодания некоторые виды «укрылись» в благоприятные места, в частности, где их обнаружил впервые М. Фернальд. Комплекс факторов в верховьях фьордов, по Э. Эбби, включает защиту от ветров, обогрев фенами и прямой инсоляцией, влагу от подтаивающих выше ледников. При определенном сочетании значений этих факторов некоторые виды могли пережить висконское оледенение в горах Торнгат на Лабрадоре, однако были ли среди них кордильерские виды, остается загадкой. |
Впоследствии исследователи еще не однажды обращали внимание на флористическое богатство фьордов и связывали этот феномен с историей. В частности, Б. Н. Городков писал: «Там, где было больше скал и невысоких горных хребтов, выступавших из льдов, должна была лучше сохраниться и растительность.
|

Шпицберген. Свалбард. Троговая долина недавно освободилась от ледника, об этом говорит почти полное отсутствие обломочного материала. Фото взято с сайта Googl eth.
|
Такие участки особенно свойственны внутренним, континентальным частям фьордов. Этим объясняется относительно богатая арктическая флора горных районов Гренландии и Шпицбергена с их фьордовыми побережьями» (Городков, 1952).
Таким образом, имеются достаточные основания считать, что в те периоды оледенения, когда ледники не достигали верховий фьордов, там сохранялась весьма богатая флора благодаря соответствующим мезоклиматическим условиям, а на их фоне условиям СЛЭ, чередующимся с условиями КДЛЭ. После зырянского оледенения наступил теплый, каргинский период. Анадырская низменность была покрыта лесотундрой (Петров, 1965, 1967; Гасанов, 1969; Васьковский и Терехова, 1970). В каргинский период море прогрессировало по долине Анадыря почти до устья р. Белая. Прибрежные части Анадырской низменности затоплялись, хотя на северном побережье Сибири каргинская трансгрессия проявилась слабо. Ляховские острова вовсе не подвергались затоплению, а, возможно, попрежнему входили в материк (Сакс, 1948). Однако о-в Врангеля, по мнению этого автора, окончательно отделился от материка в данный период. Каргинский период в целом был теплый, что доказано радиоуглеродными датировками по соответствующей древесине (Кинд,1973; Ложкин, 1973 и др.). Отдельные отрезки этого периода были теплее, чем современный климат. Со времени каргинского периода общий относительный уровень поверхности, по крайней мере, некоторых низменностей северо-востока понизился на 50—80 м. Нижние уровни ландшафтов в это время были заняты ЛЭ со значительной ролью биотической составляющей, а роль КДЛЭ, КЭЛЭ, СЛЭ существенно снизилась. В период последнего (сартанского) оледенения, наоборот, КДЛЭ, КЭЛЭ и СЛЭ широко распространились на нижних уровнях, а ЛЭ со значительной ролью биотической составляющей среды исчезли. В этот период образовалось много тонких минеральных частиц, впоследствии переработанных в покровные суглинки, способствовавшие заболачиванию ландшафтов. Последнее началось сразу с началом потепления в раннем голоцене и явилось основной причиной вымирания крупных плейстоценовых животных.
Голоценовое потепление вновь обусловило увеличение роли биотической составляющей среды на нижних гипсометрических уровнях, максимально проявившееся в оптимальное время. Климатический оптимум сменился похолоданием, во время которого началось формирование булгунняхов 2000—3000 лет назад (Васьковский, Терехова, 1970), В. Н. Сакс (1948) писал, что «следует считать бесспорным, что такие бугры (булгунняхи, Ю. К.) образуются в определенной сравнительно узкой ландшафтной зоне, а именно в северной части зоны лесотундр и в южной части зоны типичных тундр». С этого времени миграции бореальных видов на Чукотку прекратились и флора в целом начала приобретать современный субарктический облик в соответствии с реконструкцией совокупностей ЛЭ. |
Гляциоизостатическое поднятие привело к расширению площадей КДЛЭ, КЭЛЭ и СЛЭ и усложнению их структур. Из приморских районов вглубь начал проникать НЛЭ. Обновился ПАЛЭ, так как увеличился сток, в результате чего многие реки развили многорукавные системы. Перестройка растительного покрова происходила на всех геосистемных уровнях. Сдвинулись к югу границы зональных геосистем, но этот процесс в значительной мере был связан с изменением в орографических геосистемах. В одних он происходил быстрее в других — медленнее, что покажем при сравнительной характеристике двух параллельных долин на средней Чукотке.

