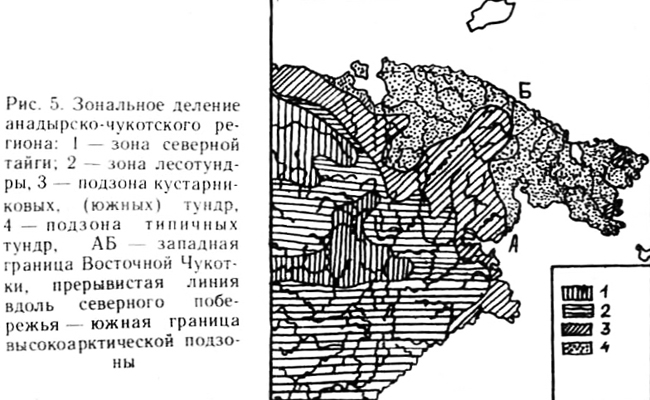
Зональное деление Анадырско-Чукотского региона
Азиатский северо-восток является территорией, которая очень перспективна для изучения ряда методических вопросов ботанического деления земной поверхности. На этой территории современные ботанико-географические особенности ее частей легко увязываются с относительно недавней историей ландшафтов по данным неботанических наук. И, напротив, эти особенности способствуют построению единой палеогеографической схемы, поскольку различные этапы флорогенеза оставили свои следы в современной флоре северо-востока Азии. Поэтому существенное значение приобретает специальное изучение вопроса о соотношении флористического, геоботанического и ботанико-географического деления этой территории. Каждое из названных делений основывается на особенностях своего собственного объекта, соответственно, флоры, структуры и роли сообществ, растительного покрова.
Как указал В. И. Грубов (1959, 1963), наиболее естественным является ботанико-географическое деление земной поверхности, а флористическое и геоботаническое районирования являются лишь двумя сторонами первого. Однако, как заметил В. М. Шмидт (1980), каждое районирование имеет свою специфику и отражает определенные свойства растительного покрова.

Озеро Эльгыгытгын окружено кольцом невысоких гор. Акварель Ю.П. Кожевникова.
Рассматривая отношение флористического районирования к ботанико-географическому, С. М. Разумовский отметил, что «сгущение границ ареалов, по Шаферу (1956) и Вальтеру (Вальтер, Алехин, 1936) свойственно ценофобным (не организованным в сообщества) видам; границы ценофильных (принадлежащих ценотическим системам) видов не сгущаются, а совпадают (по Толмачеву, 1960), образуя объективно существующие флористические линии. Эти линии, имеющие климаткчески-ривалитантный характер, являются единственно реальной основой объективного флористического районирования, совпадающего с единым ботанико-географическим районированием» (Разумовский, 1969). Совпадение границ обнаруживается лишь при работе с точечными (а не контурными) изображениями ареалов. Эти интересные соображения столь же подтверждаются в природе, сколь и не подтверждаются на территории анадырско-чукотского региона, где имеются многочисленные ценофильные и ценофобные виды и целый ряд границ и флористических линий. Ботанико-географическое деление более соответствует комплексному природному. Оно может быть осуществлено и для недостаточно исследованных в ботаническом отношении территорий, т. е. оно может быть проведено, с одной стороны, как предварительное, а с другой стороны, является основным, и при дальнейших исследованиях возможна его корректировка.
Геоботаническое районирование не всегда рассматривается как принципиально отличное от ботанико-географического, но они, конечно, очень близки. Геоботаническое районирование Арктики В. Д. Александровой (1977), по существу, представляет собой ботанико-географическое, так же, как и «Геоботаническое районирование СССР» (1947). Подлинное геоботаническое районирование, на наш взгляд, должно быть основано на картографическом материале, отражающем структуру растительного покрова. Остальные же схемы районирования, хотя и называемые геоботаническими, являются ботанико-географическими. Весьма аналогичный подход существует в ландшафтоведении, которое по существу очень тесно смыкается с ботаническим делением земной поверхности. Согласно Ф. Н. Милькову (1964), в природе существуют два, хотя и тесно взаимосвязанных, ряда ландшафтных комплексов: региональный и типологический. Региональный комплекс (район, подрайон, зона, страна) представляют единицы ландшафтного районирования, типологический — единицы ландшафтного картирования. Оба комплекса имеют самостоятельные системы таксономических единиц: тип урочища, тип местности, тип ландшафта. Д. 'Т. Арманд (1952) считал, что районирование должно начинаться «сверху». С этим согласуется представление А. И. Спиридонова (1959), который писал о том, что типологический и региональный подходы можно сочетать в единой системе, выделяя сначала типы регионов, а затем внутри каждого типа конкретные регионы. Районировать — это значит одновременно и делить, и объединять территории в соподчиненные единицы различного таксономического ранга (Ефремов, I960). К ошибкам, допускаемым в процессе районирования, Арманд (1952) относил выделение частей территории по разным признакам.
Можно сказать, что геоботаническое районирование в основном представляет статическое состояние современного растительного покрова, так как отражает физикогеографические условия, особенно на достаточно большой территории. Флористическое районирование возможно лишь для территорий с основательно выявленной флорой. Оно отражает динамическую тенденцию растительного покрова и должно опираться, на наш взгляд, на реликтовые и, наоборот, прогрессивные элементы флоры.
Еще в 1916 г. Б. Н. Городков писал, что при делении какой-либо страны на растительные области мы исходим из двух принципов: 1) проводить границы между областями, основываясь на распространении отдельных видов или их групп; 2) основываться при разграничении страны на растительные формации, пренебрегая до известного предела отдельными видами. Благодаря первому методу, при изучении карты мы не получаем достаточно ясного представления о географических особенностях территории, так как распределение по земной поверхности различных элементов флоры зависит в значительной степени от древних центров развития ее.
Таким образом, мысль о том, что флористическое районирование более тесно связано с историей флоры региона и, следовательно, должно отражать до известного предела эту историю, а ботанико-географическое деление земной поверхности призвано запечатлеть современное состояние растительного покрова, содержалась еще в данной работе Б. Н. Городкова, хотя он и не обозначил это как проведение сэмостоятельных районирований в духе нашего времени.
Флора более консервативна, а растительность более динамична. Этот феномен помогает установить не только направленность изменения растительного покрова, но и климата, хотя пока нам мало что известно о соотношении скоростей изменения флоры и растительности. Подобные же отношения существуют между растительным покровом и почвой, что было известно еще Б. Н. Городкову (1916), который писал: «...границы почвенных областей не всегда точно совпадают с границами ботанических в силу большей инертности почв по отношению к вековым изменениям климата». Вместе с тем, в каждом ландшафте формирование почв связано с сукцессиями растительности, поскольку они развиваются в одном направлении (Major, 1951).
|

Чукотский полуостров. Поселок Лаврентия. Фото А.А. Галанина.
|
Являющийся административной единицей Чукотский национальный округ охватывает несколько природных зон и подзон. Трактовка различающихся регионов у разных авторов часто очень несходна. Еще большие различия существуют в положении природных границ, поскольку разные авторы зачастую руководствуются неодинаковыми критериями. Существенно и то, что география избранных критериев нередко изучена недостаточно.
Физико-географы северную полярную область делят на две зоны: субарктическую и арктическую. Первая делится далее на две подзоны: южную (кустарничковые тундры на юге и мохово-лишайниковые на севере) и северную (арктические тундры близ побережий и на некоторых островах) (Григорьев, 1970; Горбацкий, 1964). В провинции чукотских горных тундр, относимой В. Б. Сочавой (1962) к северопритихоокеанской физико-географической области, доминируют ландшафты горных тундр берингийского типа. Наиболее повышенные и подверженные действию ветров местоположения заняты горными полярными пустынями. Река Амгуэма представляет внутрипровинциальный рубеж. На восток от нее наиболее резко сказывается влияние северотихоокеанских циклонов, которые умеряют зимние температуры, влияют на ветровой режим и осадки. Провинция чукотских горных тундр, по В. Б. Сочаве (цит. соч.), граничит на юге с Анадырско-Пенжинской физико-географической провинцией. Б. Н. Городков (1933) Анадырско-Пенжинскую депрессию (за исключением верховий р. Анадырь) рассматривал как лесотундру. Верховья Анадыря он относил к горнолесному району Колымского хребта.
Важнейшей природной границей на территории округа является южная граница Арктики, оконтуривающая собственно Чукотку (в физико-географическом отношении). За южную границу Арктики обычно принимают северную границу относительно сплошного распространения древесной растительности (Городков, 1939; Кэмпбелл, 1948; Толмачев, 1956, 1960; Polunin, 1960; Young, 197 I; Meusel et al., 1965; Александрова, 1971; Юрцев, 1974 и др.) или южную границу относительно сплошного распространения тундрового |
ландшафта (Горбацкий, 1964). В некоторых районах Земного шара северная граница леса совпадает с изотермой июля 10° С. Однако в континентальных районах Сев. Америки и в Скандинавии такого совпадения не наблюдается (Young, 1971), так же как на севере Якутии (Медведев, 1952) и Дальнего Востока. Из этого следует, что полярная граница леса определяется не только температурой. Положение изотермы 10° самого теплого месяца изменяется год от года. Так, в Канаде, в районе Гудзонова залива за более, чем 16 лет, ее положение варьировало на расстоянии около 1000 км (Tuhkanen, 1980).
Еще в начале нашего века Брокман-Ерош считал, что полярная и Субальпийская границы леса в разных частях Земного шара не связаны с действием одного какого либо «ведущего» фактора, т. е. связана с комплексом факторов (Будыко, 1971). Давитая и Мельник в 1962 г. показали, что на Кольском полуострове и в горах Кавказа граница леса «соответствует практически одинаковой сумме температур поверхности растительного покрова». Эта сумма является следствием взаимодействия комплекса биотических и абиотических факторов. М. И. Будыко и А. А. Григорьев (Григорьев, 1966) связывали безлесье с избыточным увлажнением, что вызывает недостаток газообразного кислорода в почве и, следовательно, препятствует развитию корневых систем деревьев. По другим данным, недостаточная солнечная инсоляция, зависящая от высокой широты местности, от большой облачности и туманов задерживает протаивание вечной и сезонной мерзлоты и препятствует росту и распространению деревьев. Отсюда вытекает связь древесных пород в высоких широтах с континентальностью климата и антагонизм их к морскому климату побережий (Медведев, 1952). По мнению С. Тюхканена (Tuhkanen, 1980), сопоставляя пределы распространения с климатическими показателями, мы имеем дело только с климатическими индикаторами, а не с действительно факторами, определяющими эти пределы. Н. Полунин (Pofunin, 1951) допускает, что за южную границу Арктики может приниматься одна из трех линий: 1) линия, проходящая в 80 км к северу от более-менее сомкнутых северных лесов (включая территории с редкостойным древостоем), 2) линия, ограничивающая северные пределы распространения деревьев 2—8-метровой высоты, но исключая кустарники; северные пределы «языков» и островов лесной растительности, отделенные друг от друга не более чем на 15° долготы, соединяются этой линией, 3) линия Норденшельда, точки которой определяются по формуле V = 9—0,1К, где V — средняя температура в градусах Цельсия самого теплого месяца, а К - самого холодного. Н. Полунин (Polunin, I 951) указывает, что линия Норденшельда более соответствует зональным изменениям растительности, чем изотерма июля 10 градусов. Формула Норденшельда представляет уравнение прямой линии, проведенной на рис. 4, на котором показано положение большинства пунктов Чукотского национального округа по соотношениям V и К (данные взяты из «Прикладного климатологического справочника...», 1960). На рис. 4 хорошо видно, что ниже линии Норденшельда находятся арктические пункты, а выше ее — бореальные. Однако станция Канчалан оказалась выше линии, т. е. примыкающей к бореальным пунктам, хотя по растительному покрову этот пункт относился к Арктике (Юрцев, 1973; 1974; Кожевников, 1978а). Отнесение это, впрочем, имело характер экстраполяции, так как в низовьях р. Канчалан ботаники не работали. Среднеиюльская температура в Канчалане 12° (Прил. 1, рис. 3) и, вероятно, этот пункт следует относить к бореальной области. Станция Угольная (63°04' с. ш., 179°23' в. д.) в точности соответствует формуле Норденшельда, т. е. находится на южной границе Арктики, что весьма согласуется с ботаническими данными. Линия Норденшельда в приложении к Чукотке показывает, что полоса редколесий и криволесий — лесотундра, в смысле Б. Н. Норина (1961, 1979), не относится к Арктике, а кустарниковые тундры относятся к ней.
|
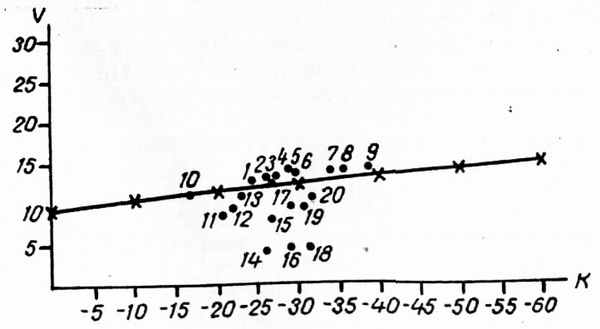
Рис. 4. Линия Норденшельда на территории Чукотского нац. округа. Неарктические пункты: 1 — Канчалан, 2 — Снежное, 3 — Березово, 4 — Танюрер, 5 — Маркове, 6—Сталине, 7 —Еропол, 8 — Островное, 9 — Усть-Олой, Переходные: Угольная. Арктические: 11— Уэлькаль, 12 — Эгвекинот, 13 —Анадырь, 1 мыс Шмидта, 15— Пенек, 16 —Айон, 17 — Перевальная, 18 — Рау-Чуа, 1 Чаун, 20 — Илирней
|
В. Ю. Визе в 1940 г. использовал формулу Норденшельда с
учетом среднегодовой температуры, которая в Арктике не превышает 0° С. Южная граница Арктики выведена им на побережье так же, как и на нашей схеме (Кожевников, 1978а), однако в глубине суши она проведена обобщенно, но сходно с нашей схемой (по Горбацкому, 1964).
Изолиния средних температур июля 10° на Чукотке совпадает не с границей леса, а с восточным пределом распространения Alnus fruticosa. В Эгвекиноте имеются лишь единичные кусты ольхи, а средняя температура июля ближайшей станции Залив Креста только 8°. Это свидетельствует о реликтовости ольхи в данном пункте. Ранее мы сделали этот вывод, основываясь на сочетании редкости ольхи и большей частоты встречаемости Lycopodium annotinum ssp . pungens, который на Чукотке тесно связан с ольхой (Кожевников, 1973а), Связь среднеиюльских температур с распространением ольхи свидетельствует также о снижении этих температур за последние столетия или даже десятилетия.
Согласно Э. Хультену, ольха предпочитает субконтинентальный климат (Bocher, 1949). В Америке ее северная граница хорошо совпадает с июльской изотермой 10°, но в Гренландии она встречается в районах, где средняя температура июля 8,5°. Т. Бёхер считает, что северный предел ольхи и древесной березы в Гренландии обусловлен сухостью климата.
Южная граница Арктики в Чукотском национальном округе с учетом расчетов по формуле Норденшельда соответствует с небольшими изменениями северной границе зоны лесотундры, которая проведена по форпостам разреженных зарослей Pinus pumila (Кожевников, 1978а). Б. Н. Городков (1933) уже давно предложил считать северный предел распространения кедрового стланика за границу горно- |
лесотундровой и горно-тундровой подзон. По мнению Н. К. Клюкина (1960), район с распространением кедрового стланика по климатическим условиям подходит к зоне тундры и лесотундры за пределами леса. Примечательно, что этот климатолог допускал существование лесотундры и тундры за пределами леса.
По сведениям А. П, Васьковского (1958), Pinus pumila достигает на правобережье Колымы 69°08' с. ш. Характерно, что на сопке у мыса Егоровича кедровый стланик поднимается выше лиственницы, но дальше к северу он не идет из-за отсутствия гор, так что граница леса на правобережье Колымы образована лиственницей (Кожевников, Андреева, 1980). В верховьях Анадыря граница распространения Pinus pumila пересекает эту реку при впадении в нее р. Пырканайваам, т. е. в 135 км от истока Анадыря (Васьковский, 1958). В долине р. Белой Pinus pumila распространен на север до ее начала (слияние рек Эньмуваам и Юрумкувеем), А. П. Васьковский,1958) указал наличие ольхи на восточном берегу зал. Креста, где она, вероятно, также представлена одиночными кустами,а близ ее впадения в Анадырь кедровый стланик образует еще сообщества лесного типа на надпойменных террасах р. Белой.
Внешние части гор южной окраины хр. Пекульней относятся еще к зоне лесотундры, поскольку в долинах встречаются заросли Pinus pumila (Сочава, 1980). Этот же автор указал заросли кедрового стланика в верховьях р. Танюрер, где их видела также Т. А. Комарова (1974). По р. Анадырь Pinus pumila достигает Кедровой кошки (Олсуфьев, 1896). В статье Т. Г. Дервиз-Соколовой (1967) о растительном покрове северного берега Анадырского лимана сообщается о находках Pinus pumila, однако определенно местонахождение не указывается; по-видимому, этот вид найден на р. Скорбутной в районе хр. Золотого. По сообщению Б. А. Юрцева (1978), Т. Г. Дервиз-Соколова встречала кедровый стланик близ пос. Шахтерского на плоской вершине холма с отметкой 150 м, где теперь находятся строения и живут люди, В настоящее время кедровый стланик в окрестностях пос. Шахтерского отсутствует. По сообщению местных жителей ближайшее его местонахождение находится в районе устья р. Великой. Еще Майдель (1868—1870) отметил, что древесная растительность здесь кончается лиственными породами и весьма не хилыми на своем пределе (Сочава, 1929). На р. Танюрер чозения и тополь указаны в его верховьях (на траверзе оз. Безымянного), а на р. Канчалан на 66° с. ш. и почти на 178° з. д. (Васьковский, 1958). Севернее тополь и чозения известна из юго-западной части Амгуэмо-Куветского массива (долина р. Юрумкувеем) и наиболее северо-восточное местонахождение чозении (уже без тополя), это — Телекайская роща на 67°45' с. ш. (Кожевников, 1974а, б, в). Здесь же находится предел распространения Betula platyphylla s . l., которая западнее встречается в долине р. Белой близ устья р. Чуванской, также на северном пределе распространения (Сочава, 1930).
|

Телекайская чозениевая роща. Вид с самолета. Фото Ю.П. Кожевникова.
|
В Верхоянском хребте древесная береза достигает 69° с. ш. (Васьковский, 1958). По данным того же автора, чозения и тополь на правобережье Колымы отстают в своем распространении к северу от лиственницы на 65 км, а от кедрового стланика на 45 км, хотя в низовьях р. Лена чозения и тополь достигают 71° с. ш. По мнению В. Б. Сочавы (1929), в Анадырском крае лесные островки из тополя и чозении по речным долинам представляют явление интразональное, а единично встречающиеся березовые рощицы — это остатки лесной зоны, отступившей в последнее время в этих краях к западу и югу в результате общего для всего северного полушария смещения на юг южной границы тундровой зоны. По нашему мнению, В. Б. Сочава не был прав, поскольку рассматривать тополевые и чозениевые леса в качестве интразональных нет никаких оснований. Они соответствуют в Анадырском крае по экологии таким же лесам в глубине лесной зоны и сочетаются со множеством бореальных формаций. Березовые рощицы представлены в басе. Анадыря (по его правобережью) слишком широко и занимают различные местоположения вплоть до склонов гор, чтобы их оценивать только как остаточные.
Таким образом, первые два критерия Н. Полунина (Polunin, 1951) для проведения южной границы Арктики в применении к Чукотке несостоятельны. Линия, проходящая в 80 км севернее более-менее сомкнутых лесов, находится в зоне лесотундры, а северные пределы распространения деревьев (высота 2—8 м) находятся глубоко в тундровой зоне. |
|

Чозения в Телекайской реликтовой роще. Фото Ю.П. Кожевникова.
|
Граница леса на севере Дальнего Востока определяется комплексным климатическим показателем — суровостью погоды, т. е. совместным охлаждающим влиянием отрицательной температуры и ветра (Клюкин, 1970). В бассейне Майна границу образуют леса из Larix gmetinii ssp. cajanderi, Betula platyphylla ssp. minutifolia, Populus suaveolens, а в районе Марково из Chosenia arbutifolia, Salix schwerinii, S. udensis. Хотя эти районы расположены всего в 50 км друг от друга климатические условия их весьма различаются, что индицируется не только различными породами деревьев, слагающих краевые леса, но и сроками вскрытия рек. Река Анадырь вскрывается близ пос. Марково около I июня (Прикладной климатологический справочник..., I960), а р. Майн, как это отметил еще Н. А. Гондатти (1897) — на 3—7 дней раньше. В это время течение в протоке Прорве, соединяющей Майн и Анадырь, происходит из первой реки во вторую. Когда р. Анадырь вскрывается, в протоке Прорве течение меняется на противоположное. В конце прошлого века пос. Марково заливался во время половодья (Гондатти, 1897), но с тех пор Анадырь углубил свое русло, и поселок не заливается. Несмотря на различие в характере лесов, оба района принадлежат лесному типу растительного покрова, и их нужно относить к подзоне северной тайги (Кожевников, 1978, 1981). В геоботаиическом отношении их, вероятно, следует включить в Колымский горный округ Якутской провинции по «Геоботаническому районированию СССР» (1947). В этих районах существует лесогольцовый тип вертикальной поясности. П. И. Полевой (1915) указал верхний край леса на Анадыре у р. Мангаля, а нижний ниже хр. Щучьего. Собственно лесной и гольцовый пояса разделены поясом горных редколесий, лиственницы или разреженных зарослей кедрового стланика. |
|

Чозения на северной границе ареала в бассейне Анадыря селится на речных галечниках. Фото Ю.П. Кожевникова.
|
По данным А. А. Кищинского (1970), пояс редколесий и стлани ков на юге Магаданской области населен типично таежной фау ной, лишь обедненной по сравнению с нижерасположенным поясом тайги. То же самое наблюдалось нами и в Анадырском крае, где пояс горных редколесий совсем беден фауной и, кроме того, здесь гнездится тундровый вид — мохноногий канюк Buteo lagopus.
Ю. П. Пармузин (1967) относит верховья Анадыря и низовья Майна к гольцово-лесотундровой Колымо-Анадырской ландшафт ной провинции, что согласуется с нашей точкой зрения, за исклю- чением эпитета «лесотундровый». В. В. Крючковым (1967) верхняя половина бассейна Анадыря (включая Майн) рассматри вается как восточная оконечность (в Азии) южной подзоны лесотундры, а вся Анадырская низменность как северная ее подзона, неограниченная автором из-за отсутствия данных на севере, в верховьях Канчалана. Разные ассоциации лиственничников не специфичны для Анадырского края, что раньше отметил В. Н. Васильев (1956). Они обычны и на севере Охотии и в Якутии.
Средняя температура июля на анадырской границе леса около 12° при среднегодовой около —9°. В Большеземельской тундре средняя температура июля на границе леса также составляет 12°, а среднегодовая —6,3° (Пьявченко, 1956). В Якутии П. М. Медведев (1952) установил, что: 1) граница леса и тундры хорошо совпадает с изотермой абсолютного максимума температуры 29°; 2) имеется хорошее совпадение этой границы с июльской изотер мой 12°; 3) некоторое общее сходство с границей леса имеет ли ния средней годовой силы ветра 4 м/сек. |
|

Береза плосколистная на склоне горы на северном пределе распространения. Фото Ю.П. Кожевникова.
|
В Канаде северная граница леса соответствует изотерме среднемесячных температур 11° самого теплого месяца в приморских районах и 12° — во внутренних континентальных районах (Tuhkanen, 1980). В последние десятилетия появилось много сообщений о том, что полярная граница леса продвигается к северу. В 1946 г. Б. А. Тихомиров отметил, что, по мнению многих, если не большинства фитогеографов, регрессия полярной границы древесной растительности, наступившая после периода послеледникового термического максимума, продолжается и в настоящее время. Однако данные ряда авторов, включая и самого Б. А. Тихомирова, свидетельствуют о «положительном продвижении древесной растительности в горы и на север, вызванном теплой климатической волной самого последнего времени». Данные такого рода продолжали накапливаться. В 1956 г. Н. И. Пьявченко сообщил, что в Большеземельской тундре граница леса наступает на тундру. В. Н. Андреев (1956) полагает, что этот процесс длится уже 500 лет, а за последние 100 лет граница леса продвигалась в глубину восточно-европейской тундры со скоростью 1 км за 5 лет. Он привел данные о повышении среднегодовых температур в период с 1942 г. по 1951 г. на 0,9° по,сравнению с периодом до 1935 гг.; при этом средняя температура января поднялась на 1,3°, а июля — понизилась на 0,9°. Климат европейского севера и Гренландии, по В. Н. Андрееву (цит. соч.), за одно десятилетие (1929—1938) потеплел в среднем на 2° в сравнении с периодом 1881 —1938. Надо отметить, что сравнение это неправомерно и поэтому в качестве аргумента о продвижении границы леса на север не годится. С 1881 по 1970 гг. во внетропической части северного полушария происходило волнообразное колебание температур (Гедеонов, 1973), поэтому в сравнении Андреева чисто случайно оказалось потепление климата. Взяв другой столь же короткий интервал времени, он мог бы обнаружить похолодание. Граница леса реагирует даже на кратковременные флуктуации климата, но в некоторых районах ее оценивают как реликтовое образование ( Tuhkanen , 1980). |
По данным В. Н. Андреева (1966), краевые островки леса в несколько десятков квадратных метров площадью из Picea obovata, Betula tortuosa, а в Западно-Сибирской низменности Larix sibirica, от п-ова Канин до Урала и восточнее представляют молодые образования. Более крупные острова леса (100—200 га) с елями 4—6 м выс. имеют возраст 80—100 лет и расположены они на южной окраине тундровой зоны; в них развиты почвы. На правом берегу Лены близ устья встречаются куртины площадью 500X100 м с сомкнутостью 0,3, выс. 2—3 м, с подлеском AInus fruticosa. Самые северные форпосты лиственницы представляют «латки» ползучих форм не более 1 м в длину, приуроченных к различного рода депрессиям микрорельефа. Самая северная граница лиственницы на правобережье Лены южнее самых северных кустов ольхи на 15—20 км, т. е. на 72° 15' с. ш. (Тихомиров, Штепа, 1956).
В. Н. Андреев (1966) отметил, что одним из доказательств заселения древесной растительностью площадей, недавно еще бывших безлесными, является довольно широкое развитие в крайнесеверных редколесьях и лесах тундровых форм микрорельефа. Этот аргумент можно рассматривать и в противположном смысле — как наступание тундры на лес. Позднее В. Н. Андреев (1970) отмечал два фронта наступления южной растительности на север — древесный и кустарниковый. Некоторое продвижение к северу кустарниковых ив мы отмечали и на северной Чукотке. По р. Кымынейвеем на самом юге Ванкаремской низменности были отмечены, например, единичные небольшие неплодоносящие кустики Salix alaxensis. Однако по их местопложению и по общему распространению этого вида на Чукотке мы более склонны расценивать краевые его местонахождения как указатели отступания Salix alaxensis на юг. В «лесном острове» Море-Ю на европейском севере ельники «проявляют признаки прогрессивного развития» (Толмачев, Токаревских, 1966). Однако то же самое можно сказать о «лесном острове» на Телекае, где прогрессивное развитие, выражающееся в наличии обильного разновозрастного подроста, характеризует чозениевый лес. Это совершенно не говорит о том, что чозения распространяется к северу.
Характеризуя островной лес Тит-Ары (на 72° с. ш.) в низовьях Лены, Б. А. Тихомиров и В. С. Штепа (1956) считали, что лесные элементы в низовьях Лены активизируют свои позиции, наступая на тундровые территории. Прежде всего,— писали они,— обращает на себя внимание поселение древесных пород на дренированных бровках тетрагональных болот, что совпадает с наблюдениями в других районах Севера СССР (бассейн р. Хатанги, восточно-европейская тундра). Авторы подчеркнули, что как ольха так и Larix в форме приземистых, прижатых к земле стлаников в настоящее время активно продвигаются в типично тундровые растительные сообщества на правом коренном берегу р. Лены. По их мнению, лесные сообщества обосновались здесь на тундровых местообитаниях 400—500 лет назад и поглотили тундровые элементы (указано, что даже Ledum palustre вымирает). По нашему мнению, вся аргументация, приведенная авторами, не является доказательством продвижения леса к северу. Стланиковую форму древесные породы принимают, не продвигаясь на безлесные ранее территории, а сохраняясь в таком обличье на территориях, где существование их нормальной биоморфы стало невозможным по условиям климата. Таким образом, те факты, которые обычно выдвигаются в пользу мнения о продвижении лесной границы к северу, можно рассматривать скорее в противоположном смысле или расценивать их как упорную тенденцию деревьев сохраниться на некогда достигнутых пределах.
Наиболее убедительные доводы о продвижении леса на прежде безлесные территории приведены для о-ва Кодьяк у южного побережья Аляски (Griggs, 1934, 1946; Bowman, 1934). Оба автора считали, что в послеледниковое время лес из ситхинской ели появился здесь впервые. По краю леса деревья имеют возраст 50 лет, а самые старые деревья в глубине леса 200 лет. Отсутствуют мертвые и упавшие деревья. Производство семян обильное, и всхожесть их хорошая. Лес оканчивается стеной. По палинологическим данным П. Баумана, лес на о-ве Кодьяк не существовал во время образования толщи торфа ниже 90 см (3 футов) верхнего слоя. Тем не менее пыльца ели (1%) обнаружена в одном болоте до глубины 220 см, в другом — до глубины 230 см, где пыльца березы составляет 8%, а вверх по разрезу ее обилие возрастает. Эти данные вынуждают относиться с осторожностью к выводам о совсем недавнем заселении острова елью. Кроме того, данные по о-ву Кодьяк вряд ли можно сопоставлять с азиатским северовостоком, учитывая то, что остров расположен значительно южнее лесов на Аляске, от которых он, правда, удален на 1000 миль (Griggs, 1946).
В Анадырском крае возраст наиболее старых деревьев превышает 200 лет. Овсянников (1930) получил следующие данные:
Возраст |
Высота |
Диаметр (в коре) |
Номер дерева |
230 |
17,5 |
27,8 |
1 |
226 |
18,5 |
25,2 |
2 |
222 |
18,3 |
19,8 |
3 |
240 |
19,3 |
25 |
4 |
206 |
17,1 |
29,5 |
5 |
212 |
16,1 |
24,8 |
6 |
210 |
15.6 |
24,8 |
7 |
По-видимому, старше лиственница здесь не встречается, поскольку отобранные Овсянниковым образцы, судя по диаметру, представляют значительную редкость. То, что лес оканчивается стеной, также не является безоговорочным аргументом в пользу точки зрения о продвижении его северной границы в тундру или даже в пределах лесотундры. Скорее наоборот, отсутствие подроста вдоль опушки может свидетельствовать о регрессивном состоянии леса. Известно, что в некоторых случаях граничные деревья не производят жизнеспособных семян или даже пыльцы (Tuhkanen, 1980).
Граничные леса, оканчивающиеся стеной, имеются и в северовосточной Азии, например, на междуречье Индигирка — Колыма и на правобережье последней реки. В бассейне р. Алазея у оз. Андрейкель их видел Г. Майдель (1894) в середине прошлого века. Он отметил такие леса и между низовьями Колымы и р. Рау-Чуа, указав, что «в большинстве случаев лес кончается сразу и недурными насаждениями». Однако для 1840 г. определен экстремум второй волны холодной фазы стадии фернау (Ловелиус, 1979). Правда, автор, видимо, считает, что этот экстремум проявился только в горных районах: «...начиная с 1820-х годов наступило резкое ухудшение условий произрастания лесов в горах Евразии, приведшее к частичной их гибели на верхнем пределе распространения» (Ловелиус, 1979. С. 52). Он иллюстрирует свои данные фотографией погибшего во время стадии фернау леса на Средне-Сибирском плоскогорье. Эта фотография очень напоминает некоторые ландшафты вдоль Майна, близ равнинной северной границы леса, но она напоминает также некоторые участки в бассейне средней Колымы, где лес погиб в силу изменившихся почвенно-гидрологических условий, а не из-за изменившихся климатических условий, поскольку район этот принадлежит северной тайге с нормально развитыми лесами. Надвигание леса на тундру в Анадырском крае отметила Л. Н. Тюлина (1936). Нам не удалось при полевых исследованиях обнаружить свидетельства, подтверждающие этот вывод.
Н. А. Гондатти (1897), собиравший сведения о природе Анадырского края, побывал в единственном тогда селении на р. Майн у Вакариной протоки. Он писал, что поселок здесь был прежде большой, «что видно по кладбищу, в последние годы совершенно размытому, но у жителей не сохранилось никаких сведений о бывшем селении». Самое удивительное, что он совсем не упоминал о лиственничных лесах в нескольких десятках километров выше по Майну, хотя отметил, что близ Вакариной протоки много наносного леса. Около пос. Маркове, по указаниям Н. А. Гондатти, «тополь и осинник» (чозения, Ю. К.) был вырублен; около села только ива по протокам образовывала лески выше 3 саженей; строевой лес имелся лишь «в верстах 10—15 от села». «Лес (лиственницу, Ю. К.) на церковь рубили повыше Еропола верст на 5, но до 50 бревен ниже Еропола, верстах в 60 от Маркова..., но лес здесь гораздо хуже» (Гондатти, 1897). Таким образом, чозенники около Маркове, как и тополевые колки,— детища нашего столетия, в то время как соотношения качества древостоев лиственницы в верхней части Анадыря и в конце прошлого века были такие же, как теперь. По данным Л. Н. Тюлиной (1936), одиночные деревца лиственницы встречаются недалеко от Маркова.
Возможно, что сама постановка вопроса о надвигании леса на тундру неверна, поскольку между этими зонами существует буферная полоса. Полоса редколесий и криволесий представляет по существу переходную зону между тундрой и тайгой. Поскольку и тундровый, и таежный ландшафты имеют глобальный масштаб, то и переход между ними характеризуется крупными самобытными признаками. Однако, принципиально, переходная полоса между тайгой и тундрой является экотоном, т. е. имеет характер градиента от типичной тайги к типичной тундре. Существует тесная аналогия между экотонами растительных сообществ и экотонами зон; различие сводится к порядку величины этих градиентов. Вместе с изменением порядка величины градиентов естественно допустить появление новых качеств в области градиента.
А. И. Толмачев (1939) подчеркнул, что существует «закономерная связь между наблюдаемыми на больших пространствах зональными изменениями и сменами растительных сообществ, происходящими на ограниченном протяжении в зависимости от чисто локальных изменений среды». По мнению А. Тэнсли (Tansley, 1935) «термин Клементса экотон может широко применяться ко всем промежуточным полосам — и климатическим, и физиографическим. Выделение экотонов не воспрещает нам использовать понятие формации в полосе экотона». Из этого видно, что А. Тэнсли имел в виду географический экотон, соизмеримый с природной зоной.
Первой по значимости величиной при сопоставлении различных экотонов (например, между сообществами и зонами) является площадь полосы экотона. Если между сообществами существуют резкие градиенты, то полоса экотона выражена слабо (довольно узкая), но даже если она выражена значительно в сравнении с площадью сообществ, она все таки остается только переходной полосой и обычно не выделяется в качестве самостоятельной единицы классификации. Аналогия между переходными полосами большого и малого масштаба усиливается тем, что и те, и другие полосы могут быть весьма различными (широкими, узкими, сплошными, прерывистыми и т. п.). В частности, как отмечал В. Б. Сочава (1929), полоса лесотундры на северо-востоке Азии вообще не выражена. Позднее он (Сочава, 1962) писал, что нередко выделяемый субарктический пояс не простирается сплошной полосой по континентам: он отсутствует в Европе и слабо выражен в Западной Сибири. Пространства, относимые к этому поясу, правильнее рассматривать как зоны внутри областей, где выражены субарктические ландшафты. Таким образом, принимается, что переходная полоса от таежной зоны к тундровой имеет значительные ботанико-географические различия в разных секторах, объясняемые не только четвертичной историей, но и современными условиями. Однако А. А. Григорьев (1970), ограничивавший Субарктику на юге границей леса, отметил, что она обладает по характеру строения и динамики географической среды столь глубокими отличиями от арктического и лесного поясов, что может быть выделена в особый географический пояс. Северной границей Субарктики может служить июльская изотерма 5° (Горбацкий, 1964).
|

|
История климатических изменений такова, что именно она обусловила характер растительного покрова Субарктики (полосы между июльскими изотермами 10°— 5° С) тем, что на этой территории происходила пульсация северной границы леса в голоцене (Тихомиров, 1944, 1947; Александрова, 1971). Было отмечено, что в современных условиях типы растительности, распространенные в Субарктике, в слабой степени зависят от радиации. Радиационный оптимум, хотя и закономерно, но весьма медленно повышается от лишайниково-моховой тундры до редкостойных болотистых хвойных лесов (Арманд, 1950). Очевидно, это объясняет тот факт, что микроклимат отдельных участков среди редколесий до некоторого предела остается тундровым. Лишь при уже весьма значительном участии древесных пород в растительном покрове, т. е. где участки с древесной растительностью преобладают над тундрой, вносится существенная фитоклиматическая поправка в околоземном слое воздуха. Следовательно, климатические показатели могут способствовать выяснению отношений между тундрой и лесотундрой, а также лесотундрой и тайгой. При этом какая то климатическая изолиния может индицировать степень связанности этих типов растительности. Индикаторные значения определяются по сугубо ботанико-географическим признакам, иначе они могут давать искаженное представление. Так, например, при исследованиях связи радиационного баланса и сумм эффективных температур было установлено, что изолиния суммы температур земной поверхности 1000° совпадает с южной границей зоны тундры, включая лесотундру (Григорьев, 1966; Будыко, 1971). Однако по ботанико-географическим и ландшафтным показателям лесотундра не может быть отнесена к тундровой зоне. Следовательно, показатель, найденный А. А. Григорьевым и М. И. Будыко, имеет не тот смысл, который они вкладывали в него. Выявленная ими изолиния совпадает с южной границей лесотундры и, по всей вероятности, сумма температур земной поверхности 1000° и определяет эту границу. |
Среди болот северной тайги и лесотундры выделяются бугристые болота с линзами льда. Они даже выделяются в особую зональную категорию «пальса», разделяющую более южные аапа-болота и более северные полигональные болота (Ruuhijarvi, 1969). По Б. А. Тихомирову (1967), лесотундра — это зональное явление, вызванное климатическими факторами. Согласно Б. Н. Норину (1961), северная граница лесотундры может быть проведена «по линии, соединяющей крайние северные участки редколесий и «лесные острова» на плакорах, т. е. включать значительную часть кустарниковой подзоны тундры». На отнесении кустарниковых тундр к лесотундре настаивают и некоторые зоологи. Например, С. М. Успенский (1969) считает, что «климат подзоны кустарниковых тундр с его особенностями инсоляции, температурного режима, режима снежного покрова, ветра и т. д. сближает их скорее с лесной, нежели с тундровой зоной. Поэтому вполне .закономерно, что доля таежных элементов в фауне кустарниковых тундр значительно выше, чем тундровых». Здесь, однако, нужно считаться с тем обстоятельством, что доля таежных элементов, приходящаяся на растения, большей частью является унаследованной со времени продвижений границы таежной зоны к северу, а доля, приходящаяся на элементы фауны, может быть и показателем современного продвижения на север птиц, насекомых и млекопитающих. Примеры такого рода приводятся самим Успенским.
Полоса кустарниковых тундр, на наш взгляд, не может быть отнесена к лесотундре, где существует специфический тип местности в понимании ландшафтоведов. Тип местности — это закономерное сочетание урочищ, «...границы типа местности определяются географией составляющих его характерных урочищ и урочищ доминантов. Подобно другим типологическим единицам, тип местности обладает разорванным ареалом, и его распространение не зависит от границ региональных единиц» (Мильков, 1964). Поэтому лесотундровый тип местности, включающий урочища болотных редколесий (или с одиночными деревьями), укрытых долинок с ивовыми лесками, горных склонов с парковыми зарослями кедрового стланика и др., могут быть встречены и в северной тайге. Как отметил Ф. Н. Мильков (1964): «В зависимости от характера региональных воздействий можно говорить о зональных, высотно-геоморфологических и литологических вариантах типа местности». В качестве особой растительной зоны, характеризующейся только ей свойственными признаками растительного покрова, претерпевшего автохтонную эволюцию параллельно с развитием таежного пояса с конца плиоцена, Б. А. Юрцевым (1966) выделен Гипоарктический пояс. Последний включает помимо субарктики полосу (подзону) северной тайги: «северная граница леса проходит в пределах Гипоарктического пояса, внутри него, подразделяя его на две основные полосы: гипоарктической тайги и гипоарктической тундры. Сама идея объединения заведомо неоднородных типов растительного покрова краевых частей широтных его подразделений не нова. И. М. Крашенинников (1939), характеризуя развитие растительного покрова Южного Урала, подобным же образом высказывался в отношении лесостепи. «Лесостепь в нашем толковании есть не просто переходная полоса от лесной к степной зоне, а вполне самостоятельная область, которая имела особую историю образования и развития своей флоры и растительности». Его понятие лесостепи было достаточно широко, включая как территории с преобладанием леса над лугово-степными группировками, так и, наоборот, территории, где луговые степи доминируют над лесом. Б. Н. Городков (1916) полагал, что «переход как тундры, так и степи в лесную зону весьма постепенен и совершается через посредство областей смешанного типа (лесотундра, лесостепь), и от вкусов автора зависит отнести эти области к той или другой зоне». На карте растительности, приводимой Е. Одумом ( Odum , 1959), переходные зоны между тундрой и тайгой и степью и тайгой названы экотонами.
Важнейшее значение Б. А. Юрцев (1966, С. 5) придал «массовому уходу с водоразделов одних и выходу на водоразделы других видов». Южная граница Гипоарктического пояса проведена Юрцевым с учетом того, что жизненный оптимум древесных пород на севере тайги лежит много южнее Гипоарктики. Все же нужно заметить, что и с учетом обоих критериев южная граница Гипоарктического пояса проведена в ряде районов (например, на европейском северо-востоке) с солидным запасом, так как сомкнутые северотаежные леса располагаются к северу от этой границы на плакорах не менее, чем на 400 км. В Западно- Сибирской низменности южная граница Гипоарктического пояса не согласована с районированием этой низменности Б. Н. Городковым (1916), который построил свое районирование на формационном принципе и по сей день оно относится к наиболее обоснованным (Крылов, Салатова, 1969). В пределах пояса имеется флористическое ядро, связанное своим происхождением с территорией пояса, однако по числу видов это ядро значительно уступает бореальной группе. По мнению Б. А. Юрцева, Гипоарктический пояс в настоящее время выполняет барьерную роль в отношении миграции растений в долготном направлении в силу специфических почвенных условий. По-видимому, это очень общий вывод, «соизмеримый» с масштабами пояса, так как при рассмотрении флор районов меньшего масштаба обнаруживаются миграции целых флористических элементов, например, на европейский северо-восток в пределы Гипоарктического пояса происходит вторжение видов центрально-европейского элемента (Кожевников, Плиева, 1976), а также видов восточно-европейского неморального элемента. И тот, и другой элементы включают серии видов, обитающих на различных местообитаниях, в том числе на местообитаниях с зональной растительностью.
Плато Путорана, целиком входящее в Гипоарктический пояс, имеет уже свои ботанико-географические особенности, одной из которых является обилие мигрантов из Южной Сибири. Наконец, гипоарктическая часть Анадырско-Чукотского региона, охватывающая большую его часть, настолько тесно связана исторически с более южными районами тайги, что нет никакого смысла говорить об общности ее развития с путоранской гипоарктикой или с аляскинской. Таежные секторы этого пояса, а равным образом и лесотундровые, можно рассматривать в единстве только в ландшафтном смысле, но не в ботанико-географическом. В этом имеется некоторая аналогия с зоной полярных пустынь, являющейся, однако, более компактной под давлением одних и тех же факторов неживой природы. Но единство таежных, лесотундро вых и тундровых подразделений одного сектора и с ландшафто-ведческих позиций совершенно нереально, поскольку несопоставимы зональные типологические характеристики, несмотря на наличие в пределах какой-либо зоны или подзоны нетипичных для нее ландшафтов.
По этому поводу Ф. Н. Мильков (1964), много работавший в лесостепи, отмечает: «...природа типологических единиц такова, что иногда они не считаются с границами региональных единиц и один и тот же тип местности может встречаться в разных ландшафтных зонах, так же, как и ареал типа ландшафта не повторяет область распространения какой либо определенной ландшафтной зоны». По нашему мнению, Гипоарктический пояс представляет собой явление такого масштаба, что, какие бы пертурбации не происходили в его пределах, он неизменно остается самим собой. Поэтому для целей исторической фитогеографии более целесообразен анализ зон и подзон, В.Д.Александрова (1971, 1977) выделяет в Арктике две зоны, совпадающие по рангу с областями: тундровую и область поляр ных пустынь. В пределах тундровой области ею выделены подобласть субарктических тундр и подобласть арктических тундр, совпадающие с подзонами, в пределах которых выделяются полосы. Основным отличительным признаком всех подразделений является характер растительности на плакорах: доминантные типы растительности, соотношение сообществ, климаксовые синузии; подобласть субарктических тундр отличается от подобласти арктических тундр также тем, что в первой корневые системы растений в сообществах сомкнуты в отличие от второй. Кроме того, все крупные подразделения имеют особенности во флористических комплексах. Однако, когда говорится, что «вариантами тундрового типа растительности являются петрофильные и псаммофильные сообщества большей частью с преобладанием лишайников», то отличие этих вариантов от зональных группировок полярных пустынь, где синузия накипных лишайников является климаксной, теряется. Очевидно, что если большие плакорные пространства будут заняты каменистыми и щебнистыми россыпями, то в пределах тундровой области, кроме горных тундр окажутся обычными фрагменты полярных пустынь.
Деление во всех случаях, перечисленных выше, основывается на комплексе признаков: флористических и ботанико-географических. По мнению Александровой (1971, 1977), «зональное деление тундр и полярных пустынь необходимо проводить по комплексу признаков, важнейшими из которых являются: состав флоры, особенности флорогенеза, набор жизненных форм, состав и структура плакорных и неплакорных сообществ, характерные типы комбинаций растительности разного ранга. В том, что такой подход правомерен в настоящее время, едва ли вызывает у кого сомнение. Однако, среди физико-географов не существует единства мнений относительно принципов районировании. Так, Ю. Ефремов (1960) настаивает на комплексе признаков, положенных в основу деления территорий, а Д. Арманд (1952) считает, что районировать следует по одному признаку, по крайней мере в пределах одного ранга единиц классификации; для различных рангов могут быть различные, но одиночные критерии. Одиночный критерий — богатство флоры — в зональном делении Арктики использован С. Янгом ( Young , 1971). На основе северных пределов циркумполярных и почти циркумполярных видов С. Янг выделяет 4 циркумполярные флористические зоны. Проделав зональное деление по северным пределам распространения растений, С. Янг затем сопоставляет это деление с климатическими факторами, именно с количеством осадков и ходом температуры. Им предложен критерий — сумма средних температур вегетативного периода — для сравнения разных во флористическом отношении территорий. Первая зона С. Янга совпадает с зоной полярных пустынь В. Д. Александровой, вторая зона весьма соответствует подзоне арктических тундр того же автора, третья зона охватывает узкую полосу суши вдоль северных побережий, исключает о-в Св. Лаврентия, четвертая зона включает всю субарктику, ограниченную с юга полярными пределами леса. Если сопоставить южные границы Арктики С. Янга и Визе, то обнаруживается значительное несоответствие. Так, у Визе Исландия, северо-западная приморская Аляска не попадают в пределы Арктики, в Северной Америке граница также проведена севернее, чем у С. Янга (между южным берегом Гудзонова залива и Большим Медвежьим озером). Во флористическом районировании Арктики Б. А. Юрцевым и др. (1978) ее южная граница проведена так же, как у Визе, но при ботанико-географическом выделении подзон бассейн Анадыря за исключением, лесных верховий, отнесен к тундровой зоне в качестве подзоны крупных стлаников. По сочетанию типов растительности, как было сказано, большая часть бассейна Анадыря относится к северной тайге и лесотундре.
При ботанико-географическом делении наблюдается небезинтересная психологическая особенность, когда исследователь, имея дело с совокупностью факторрв, не анализирует эту совокупность по составляющим, но, пользуясь в значительной мере интуицией (которая включает подспудным образом знание этой совокупности фактов), выводит результат исследования (т. е. проводит границы) и уж затем начинает анализ совокупности фактов, доказывая правильность результата, обосновывая его, и опираясь на него в ходе доказательства.
|

Лесотундра в континентальных районах Чукотки.
|
Проведение южной границы лесотундры также приводит к несогласиям разных авторов. Зачастую к лесотундре относят и часть подзоны северной тайги, если там на плакорах преобладают редколесья, не считаясь с тем, что прочая растительность является бореальной. А. А. Григорьев (1970. С. 257) писал: «На северо- востоке Азии самым южным из обширных равнинных низменных тундровых массивов, вкрапленных среди господствующих здесь горных поднятий, является Парапольский Дол, простирающийся к югу до 60,5° с. ш.». Л. Хамет-Ахти (1976) отнесла Парапольский дол к северобореальной растительности, несмотря на то, что там нет даже редколесий. Для многих ботанико-географов и ландшафтоведов такой взгляд явился неожиданным, однако его правильность не вызвает сомнений и помогает адекватным образом оценить и другие территории азиатского северо-востока. Мойзель и др. (Meusel et a!., 1965) отметил, что зона лесотундры, как ее показал Б. Н. Норин вслед за Ю. П. Пармузиным (1958), отчасти соответствует субарктической зоне в их понимании, отчасти суббореальной. То, что Стеффен в 1935 г. называл субарктической зоной, авторы отнесли к бореальной области. А. П. Васьковский (1958) считал, что тундра простирается на юг до берегов зал. Бабушкина и занимает южную часть п-ова Кони (58°55'15" с, ш.), а также острова Тауйской губы, т. е. до северного побережья Охотского моря. А. П. Хохряков (1971) отнесся весьма сдержанно к мнению Васьковского, пояснив, что «появление таких тундрочек можно поставить в связь не только с влиянием зональных арктических тундр, но и со снижением верхней границы леса в прибрежных районах». Действительно, типичные тундры в северной Охотии очень фрагментарны и приурочены исключительно к верхнему поясу гор. Что же касается горных тундр, то В. Б. Сочава (1980) обозначил их простирание на юг до Прибайкалья и Северной Кореи. |
Б. Н. Городков (1944), исследовавший различия между тундрой и альпийским поясом гор, писал, что «лишь северные горные страны, климатические особенности которых приближаются к арктическим, имеют альпийскую растительность, сходную с тундрой». Но в общем тундры отличаются от альпики тем, что корневые системы растений в них сосредоточены в верхних горизонтах, а альпийские растения простирают корни до 50 см вглубь. В альпийской растительности не образуется дерновина из мхов. Кроме того, альпийские луговые почвы отличаются от тундровых отсутствием глеевого горизонта (Daubenmire, 1947). Тундровая и альпийская растительность сходны лишь на местообитаниях с обильными снежными скоплениями или с оголенной зимой землей (Городков, 1938). При характеристике зональности в горных странах упор необходимо делать на типы растительного покрова нижних гипсометрических уровней.
Пределы распространения деревьев в укрытых местоположениях определяют границы между лесотундрой и кустарниковыми тундрами. В последних виды деревьев существуют лишь как кустарники или стланики. Эти критерии со всей очевидностью не подходят для Чукотки, где в континентальных районах деревья встречаются группами, хотя основной фон растительного покрова создают тундры. Некоторые древесные породы, например, Betula platyphylla, распространяются и в относительно океанические районы, где они, однако, принимают кустарниковую форму роста. В Гренландии Т. Бёхер (Bocher, 1949) также отметил, что усиление океаничности климата, имеющее результатом прохладное лето, ставит предел распространению древесных форм, но не видов, к которым принадлежат эти формы. Так, Betula pubescens достигает южной оконечности Гренландии в виде куста. Как отметил С. Тюхканен (Tuhkanen, 1980), при зонально- провинциальном делении иногда используют априорное допущение, что зональность растительности определяется количеством тепла и радиации, а провинциальность (секторы) основана на степени континентальности — океаничности.
Характеризуя климат Западной Гренландии и увязывая с ним особенности растительного покрова, Т. Бёхер (Bocher, 1951) подразделяет его на зональные и провинциальные категории. Так, климат южной Гренландии он оценивает как субарктический субокеанический с Betula pubescens и ее спутниками. Южная Гренландия по геоботаническим признакам является лесотундрой (Тихомиров, 1959). Ее населяют Betula tortuosa, Alnus ovata, Sorbus americana, Juniperus communis. Обширные ранее площади березняков теперь сократились из-за пастьбы овец. Уже с давних пор там ведутся посадки древесных пород. Однако Picea abies и Pinus silvestris, посаженные в 1899, частично выпали, хотя значительная часть деревьев сохраняется. Севернее (61°- 65° с. ш.) следует субарктический климат с массивами Alnus crispa, затем низкоарктический континентальный с Ranunculus lapponicus, Artemisia borealis, Ledum palustre ssp. decumbens (только севернее 64° с. ш., а южнее предпочитает внутренние районы). И на северо-востоке Гренландии он определяет климат как высокоарктический субокеанический с Arenaria pseudofrigida. Зональности растительности на севере Канады посвящена недавняя статья К. Аткинсона (Atkinson, 1981). Он отметил, что термин «субарктика» вызывает много проблем. Этот термин определен И. Хустихом (Hustich) в 1966 г. и Д. Лёве (Love) в 1970 г. как эквивалент «хемиарктики» и лесо-тундровой переходной полосы. Далее К. Аткинсон подчеркнул, что в отличие от взглядов Дж. Руссо (Rousseau), высказанных в 1952 г., лесо-тундровая полоса характеризуется прежде всего отсутствием деревьев, а не наличием представителей арктической флоры. Безлесные тундровые водоразделы и вершины холмов покрыты теми же видами лишайников и кустарничков, которые населяют северо-бореальные редкостойные леса. Большинство видов тундровых водоразделов субарктики являются скорее бореальными, чем арктическими. Поэтому лесотундру более целесообразно рассматривать как северный предел бореальной области и субарктики, чем в качестве переходной полосы к арктике. На северо-западе Канады установлены следующие зональные категории: арктика (тундра), высокая субарктика (кустарниковая тундра и лесотундра), низкая субарктика (редкостойные лишайниковые леса), высокий бореальный регион (хвойные леса), средний бореальный регион, (смешанные леса). Южная граница высокой субарктики проведена по форпостам леса на водоразделах. Следовательно, лесотундра начинается там, где кончаются леса на водоразделах.
Редкостойные лишайниковые леса низкой субарктики Канады образованы в основном Picea mariana. Южная граница этого региона проведена Аткинсоном там, где леса становятся сомкнутыми. Западнее Большого Невольничьего озера эта граница совпадает с линией края отступившего ледника 8000 лет назад. Кроме климатических факторов, определяющих границы субарктики, немалое значение имеют пожары, очень там обычные. Палеоэкологические условия также наложили свой отпечаток на современный облик субарктики. Во время голоценового климатического оптимума летние температуры были на 5° выше современных и леса продвинулись на несколько сотен км к северу. Похолодание началось 4000 лет назад, но происходили осцилляции; заметное потепление было 1200—1000 лет назад. Со времени оптимума в укрытых долинах сохранились изолированные хвойные криволесья.
Как показал наш опыт, ботанико-географические рубежи устанавливаются и получают убедительное обоснование при длинномаршрутных исследованиях, например, спускам но рекам, пересекающим эти рубежи, с остановками на 1—3 дня в местностях с какими-либо ландшафтными новшествами для коротких маршрутов (10—20 км в один конец). При таких действиях в поле зрения попадают многие признаки растительного покрова в их динамике, соответствующей климатическим изменениям. Правда, сами климатические изменения обычно могут быть охарактеризованы лишь по косвенным данным и по облику растительности. Однако при последовательном исследовании связь растительности с климатом обычно прослеживается в деталях. Эта связь дает нам возможность дать такую же семантическую характеристику климата по территориальным выделам с определенным сочетанием типов растительности и растительного покрова, какую дал Т. Бёхер в Гренландии. Для подзоны северной тайги характерными типами растительности являются лесной, лесотундровый, кустарниковый, болотный, горнотундровый, луговой; редкие типы: степоидный, степной. Основным условием существования этой подзоны является резко континентальный климат. Предельные значения континентальности климата для подзоны северной тайги приблизительно соответствуют району пос. Марково, где существуют такие формации лесного типа растительности в таежном типе растительного покрова, которые обнаруживаются и в других типах растительного покрова.
Следует отметить, что в приморских бореальных районах, где климат, хотя и океанический, но допускает существование лесной растительности, растительный покров весьма слабо дифференци рован. На Камчатке Н. В. Павлов (1936) установил, что растительный покров этой страны еще не имеет установившихся и резко отчлененных друг от друга растительных группировок даже в более крупном ранге формаций или типов растительности, а не только ассоциаций. Он считал, что березняки и высокотравье нужно рассматривать как одно целое, некоторый комплекс, возникающий в результате заселения высокотравных лугов деревьями березы. Э, Хультен (Hulten, 1960, 1972) даже считал, что растительный покров не установился на Камчатке и на Алеутских островах со времени ледникового периода. Однако это обусловлено только современными условиями Камчатки и Алеутских островов. В других районах, в том числе анадырском с резко континентальным климатом, со времени ледникового периода растительный покров неоднократно перестраивался в соответствии с изменением климата.
Для зоны лесотундры — характерные типы растительности — лесотундровый, кустарниковый, болотный, тундровый, горно тундровый, луговинный; редкие типы: лесной, степоидный. В. Н. Васильев (1956) выделил подзону кустарниковой тундры, которую он делит на полосу кедрового стланика и полосу кустарниковой ольхи. Его подзона кедрового стланика в нашем понимании является зоной лесотундры. Для этой зоны очень характерны заросли крупных приречных ивняков, которые исчезают при движении вниз по Анадырю в районе Утесиков (Тюлина, 1936). Согласно П. И. Полевому (1915), реку Анадырь можно разделить на три части: нижнюю — до Утесиков, характеризующуюся действием приливной волны, среднюю — до горы Опаленной выше Марково, верхнюю — горную. В районе Утесиков нужно проводить границу лесотундры при пересечении р. Анадырь. Далее вверх по Анадырю лесотундровый ландшафт тянется до протоки Прорва, соединяющей Анадырь с Майном. Климат в зоне лесотундры менее континентален, чем в подзоне северной тайги, о чем свидетельствует долгое залеживание снежников даже близ устья Майна. Он может быть определен как резкоконтинентальный субарктический с переменной жест костью. С Анадырского залива сюда поступают массы морского воздуха. Древесная растительность приурочена в основном к укрытым местоположениям или к надпойменным террасам, но редкие деревья встречаются н на равнинных участках (лиственницы, березы).
Основная причина безлесья климатическая, но, кроме того, действуют и другие причины. Как отметил П. М. Медведев (1952), безлесье тундры связано не только с наличием вечной, но и долго задерживающейся весной сезонной мерзлоты. Особое влияние на древесную растительность, очевидно, имеет изменение почвенно-гидрологических условий. В частности упоминавшийся мертвый лиственничник погиб вследствие поднятия вечной мерзлоты. Характерно, что вдоль его окраины сохранились редкие живые деревья, а некоторые одиночные лиственницы были отмечены по днищу лощины рядом с мертвым лесом. Следовательно, происходит физиологический отбор деревьев, и отдельные индивиды его выдерживают. На спиле одного из них не было обнаружено каких-либо отклонений в характере распределения годичных колец. По данным В. Ф. Овсянникова (1930), во всех лиственничниках мерзлота находится не менее 47 см от поверхности почвы. Исследовав майнские лиственничники, этот автор подтвердил представления Б. Н. Городкова о «решающем значении почвенно-грунтовых условий по отношению к продвижению границы древесной растительности к северу». Для роста уже существующих деревьев на пределе наиболее критическое время года — весна; для семенной продуктивности и, следовательно, для продвижения к северу очень важно и летнее тепло, отметил П. М. Медведев (1952), указав также, что в тундру попадают низкокачественные семена с окраины леса. Таким образом, безлесье обусловлено целым комплексом причин, каждая из которых оказывает неблагоприятное действие и сама по себе, так что в разных местоположениях безлесье может быть обусловлено разными причинами.
Горные системы в зоне лесотундры подвержены влиянию морских воздушных масс, но кедровый стланик на них весьма обычен, хотя обычно образует лишь разреженные заросли. В этой зоне он уже почти не встречается на болотах как в северной тайге. Континентальность, а вместе с тем жесткость климата в пределах зоны очень варьирует. Она снижается по направлению к востоку (к Анадырскому заливу) и к северу. Очень резкие ее градиенты существуют по внешним окраинам горных систем, например, в районе пос. Отрожного, что связано с постепенным увеличением высоты местности. На маршруте от р. Анадырь к горам можно встретить лесной тип растительности (вдоль проток близ реки), затем кустарниковый, болотный и, наконец, типично тундровый, а у подножий гор отмечались нивальные луговины, свидетельствующие о наличии здесь в первой половине лета снежников. «Фактор мезоклиматических и местных эдафических условий в распределении лесных островов играет в лесотундре большую роль, чем в северной тайге» (Лукичева, 1967). Переход лесотундровой зоны в подзону кустарниковых тундр очень постепенный, и линия на карте отображает этот переход только очень обобщенно. Он имеет мозаичное строение, т. е. на одних местообитаниях существует лесотундровая растительность, а на других — тундровая.
Для подзоны кустарниковых тундр характерные типы растительности - это кустарниковый, болотный, тундровый, горнотундровый, луговинный (причем нивальный подтип довольно редок); к редким типам относятся лесной, степоидный и степной. Южнотундровая растительность межгорных впадин почти достигает р. Амгуэмы на ее долготном отрезке. В этой части долины Амгуэмы, отделяющей Чукотский п-ов от материковой Чукотки, еще встречаются склоновые заросли ольхи и ив и низкокустарниковые ерники. Ботанико-географическая граница, отделяющая берингийскую Чукотку от ее континентальной части, является в то же время границей подзон кустарниковых и типичных тундр. Лишь близ северного побережья (рис. 5) она разделяет западный и восточный участки Чукотки, относимые к «настоящей Арктике» в смысле Н. Полунина (Polunin, I 951). Изолированно южнотундровая растительность встречается на востоке Чукотского полуострова (Юрцев, 1973, 1974; Секретарева, 1979, 1982), южнее Полярного круга. Амгуэмо-Куветский массив и бассейн Амгуэмы расположены севернее Полярного круга. Подзона кустарниковых тундр относится уже к Чукотке, тогда как лесотундра — к Анадырскому краю. Таким образом, граница между Чукоткой и Анадырским краем весьма расплывчатая, тогда как граница между подзоной северной тайги и зоной лесотундры в пределах Анадырского края довольно резкая. Западная граница Чукотки весьма условно проводится в низовьях Колымы. В. Б. Сочава еще в 1933 г. показал, что самые низовья Колымы разделяют Восточно-равнинно-тундровый (к западу от реки) и Восточно-горно-тундровый (к востоку от реки) районы.
|
|
К северу от пос. Черский вниз по Колыме лиственничные леса, в основном горелые, тянутся по правобережью до мыса Егоровича, а затем выклиниваются. Остаются лишь отдельные деревья, индицирующие узкую (порядка 20 км) полосу лесотундры. На левобережье Колымы вдоль края надпойменной террасы выс. Зм близ села Михалкино (в углу между протокой и рекой) имеются обширные парковые ивняки высотой до 1,5—2 м с Salix alaxensis, S. krylovii, S. glauca, S. boganidensis, S. hastata, S. lanata ssp. richardsonii, S. schwerinii, S. udensis, бедные видами разнотравья, а местами с голубикой, морошкой, березкой тощей и багульником. Ивняки переходят в густые заросли ольховника до 2 м высотой (крупный массив), которые изреживаются по мере удаления от дренированной части террасы.
На правом, более возвышенном, берегу р. Колыма кустарниковая растительность более угнетена и сосредоточена близ ручьев между увалами или на участках надпойменной террасы Колымы. Отдельные ивняки имеются также на склонах в ложбины и к Колыме. Следует заметить, что выше по Колыме (в пределах таежной зоны) возвышенный берег способствует большему облесению, тогда как в низовьях такой берег способствует большему отундровению ландшафта. |
Вдоль Колымы против Михалкино тянется высокий и крутой склон увала, выходящего к реке и обращенного к северу. Его нижняя половина высотой 8 м с крутизной 30° занята кустарничково-моховой сплошной тундрой с Cassiope tetragona, Dryas octopetala ssp. viscida, Betula nana ssp. exilis, Ledum palistre ssp. decumbens, Parrya nudicaulis, Saxifraga nelsoniana, Hedysarum hedysaroides ssp. arcticum, Polygonum tripterocarpum, Poa malacantha, Vaccinium vitis-idaea ssp. minus, V. uliginosum ssp. microphylla, Luzula unalaschkensis, L. nivalis, Minuartia macrocarpa, Astragalus alpinus, Salix glauca, S. pulchra, Stellaria ciliatosepala, Claytonia acutifolia, Hierochloe alpina, Valeriana capitata, Pedicularis lapponica, Oxyria digyna.
В верхней части этого же склона больше щебнистых (из углистого сланца), чем дерновых, участков. Здесь растут: Myosotis asiatica, Arnica frigida, Artemisia arctica, Erysimum pallasii, Saxifraga cernua, S. firma, Cerastium beeringianum, Polygonum viviparum, P. ellipticum, Minuartia rubella, Draba iactea, D. hirta, Rhodiola rosea, Papaver radicatum s. I., Cystopteris fragilis, Trisetum spicatum, Delphinium chamissonis. Осыпные участки в верхней части склона чередуются со скальными, не отличающимися заметно по набору видов. Здесь можно выде лить кустарничковые, редкотравные щебнистые сообщества и группировки. На плоских увалах правобережья распространены сухие кустарничково-моховые тундры с морозобойными пятнами сухого суглинка без щебня с бугорковатым микрорельефом. Местами на увалах бугорковатый, преимущественно абиогенный микрорельеф переходит в кочковатый биогенный с Сагех lugens.
В межувальных лощинах с ручьями вполне обычны и разнотравные разреженные ивнячки высотой 30—50 см из Salix lanata ssp. richardsonii c Nardosmia frigida, Aconitum delphinifolium, Poa arctica, Arctagrostts arundinacea, Alopecurus alpinus s. str., Salix reticulata, Valeriana capitata, Polygonum ellipticum, Vaccinium uiiginosum. На пологих скатах в лощины встре чаются низкие куртины Salix krylovii (отсутствующей в других местоположениях) в составе луговинной растительности.
На щебнистых южных участках склонов изредка встречаются редкотравные группировки, напоминающие обликом степоиды с Potentilla stipularls, Arenaria tschuktschorum, Arnica iljinii. На таких же склончиках противоположной экспозиции, очевидно, проявляется нивальный процесс, так как пустоты щебня наполнены нивальным наилком, и здесь растут в обилии нивальные виды, например, Saxifraga hyperborea. Нивальные признаки в ландшафте правобережья Колымы в данном районе выражены достаточно разнообразно, тогда как на левобережье они практически отсутствуют. Учитывая еще и то, что кустарниковая растительность на левобережье выражена как в подзоне кустарниковых тундр, а на другой стороне Колымы — как в подзоне типичных тундр, следует признать, что Колыма разделяет здесь два типа ландшафтов, но оба они характерны для Чукотки, поэтому ограничение Чукотки на западе устьем Колымы абсолютно условно. Возможно, что оно менее естественно, чем граница по водоразделу бассейнов Колымы и Анадыря.
Степоиды обнаружены в районе Походска (Андреев и др., 1978, Андреев, Галактионова, 1981). Цитируемые авторы называют их «реликтовыми степными фрагментами» (стр. 45), однако, судя по их описанию (стр. 50) — это именно степоиды, и вовсе не следует думать, что под давлением тундровой растительности они исчезнут в ближайшие десятилетия. Они исчезнут на тех местах, где существуют теперь, но появятся в другом месте, где в силу каких-либо причин произойдет эрозия растительного покрова. Авторы сами указывают на значение этого фактора, отмечая, что для «сообществ» Сагех spaniocarpa благоприятна деятельность роющих животных, нарушающих целостность покрова мхов и кустарничков. Причинами эрозии могут быть также оползание грунта, разрушение дернины ветром в зимнее время и т. д.
Давая обшую ботанико-географическую оценку района с. Михалкино в сравнении с другими районами северо-востока Азии, мы сразу можем обратить внимание на значительную общность этого района и низовьев Анадыря (Кожевников, 1981). Точно так же там имеются смежные ландшафты, один из которых без натяжки относится к подзоне кустарниковых тундр, а другой - ко вполне типичным тундрам. Точно так же ольховник встречается там в болотах и имеется комплекс тундровых и бореальных птиц. Отмечалось, что в полосе контакта двух флористических областей близ устья Колымы наблюдается значительное обеднение арктическими и бореальными видами (Петровский, Заславская, 1981). Было бы, конечно, странно ожидать противоположную ситуацию в этой полосе, тем более, что обеднение это не характеризует собственно полосу контакта. Оно наблюдается непрерывно от среднего течения Колымы при движении на север.
В подзоне кустарниковых тундр климат в целом умеренно континентальный субарктический, но в ряде урочищ он резко континентальный. Климат Субарктики, как указал А. А. Григорьев (1970), зависит от степени континентальности, а в приморских районах от степени его суровости. К этому можно добавить, что в южной Субарктике климат особенно сильно дифференцирован на систему местных, мезо- и микроклиматов, вследствие различного влияния морских воздушных масс из-за условий рельефа и формирования локальных или местных физико- географических процессов, препятствующих адвекции воздушных масс в околоземном слое воздуха. Однако общий характер климата в пределах подзоны изменяется мало, несмотря на то, что одни районы подзоны являются приморскими, а другие внутренними.
Согласно С. П. Хромову (1968), континентальность-океаничность климата есть не экологическая, а географическая характеристика, так как «величина годовой амплитуды температуры зависит не просто от характера подстилающей поверхности или от близости данного места к береговой линии. Она зависит от повторяемости в данном месте воздушных масс морского и континентального происхождения, т. е. от условий общей циркуляции атмосферы». Мнение климатолога в этом вопросе, разумеется, авторитетно, однако следует заметить, что вторгающиеся воздушные массы по-разному взаимодействуют с элементами ландшафта, и поэтому можно оценивать относительную континентальность-океаничность мезо- и даже микроклиматов. Локальные условия для растительности часто важнее, чем общий климат. Это учитывают специалисты по интродукции растений. Так в руководстве лесоводам о принципах выбора пород деревьев для посадок в разных местах Англии в зависимости от среды сказано, что локальные факторы значительно важнее общих факторов (Anderson, 1961). Исследование интродуцированной из северо-западной Америки Picea sitchensis на северо-востоке Шотландии показало, что рост деревьев положительно коррелирует с температурой почв в течение вегетационного периода и с количеством зимних осадков. Рост деревьев тесно связан также с высотой, положением на склоне, с наличием геоморфологических укрытий, но не связан с экспозицией и крутизной склонов. Наибольшая корреляция роста деревьев обнаружена с количеством почвенной органики и, следовательно, содержанием фосфора. Обнаружено отрицательное влияние на рост деревьев калия. На максимальных высотах климатическое влияние на рост деревьев превосходит влияние дренажа.
Для подзоны типичных тундр в качестве характерных типов растительности нужно назвать тундровый, горнотундровый, луговинно-нивальный, болотный; редкие типы; кустарниковый, степоидный, высокоарктичёский. Эта подзона на Чукотке имеет как бы два крыла: океаническое и материковое. Соответственно климаты различаются на субарктический океанический и субарктический умеренно континентальный. В обоих «крыльях» подзоны хорошо представлены реликтовые элементы континентальной растительности, которые обильнее в материковом крыле. Наличие этой растительности обусловлено прежде всего дифференциацией общего климата на мезо- и микроклиматы. Существование местных климатов, т. е. климатов местности (Сапожникова, 1950; Хромов, 1952), резко уклоняющихся от общего климата в подзоне типичных тундр практически не выражено, хотя нельзя исключить, что это проявляется кое-где в материковом крыле.
Лучшая представленность реликтовой континентальной растительности вовсе не свидетельствует о том, что и климат здесь является более континентальным. Так, при аналитическом сравнении районов оз. Эльгыгытгын на Анадырском плоскогорье и г. Анадырь нами было установлено, что оба этих района относятся к континентальной ботанико-географической подпровинции Чукотки, но район оз. Эльгыгытгын расположен в подзоне типичных тундр, а г. Анадырь — в подзоне кустарниковых тундр, причем близ границы с лесотундрой. По соотношению континентальных и океанических признаков флоры и растительности оказалось, что в районе оз. Эльгыгытгын, несмотря на его континентальное положение, усиливаются океанические признаки растительного покрова. Это подтверждают и другие наблюдения: находки в поверхностных слоях грунта стволов крупных ив, отсутствующих здесь в настоящее время, орнитологические находки (Кожевников, 1977в), геоморфологические изменения в течение последних 100 лет по описаниям ранних исследователей. Усиление океаничности климата в районе оз. Эльгыгытгын и соответствующее изменение растительного покрова обусловлено поднятием местности над уровнем моря.
В районе г. Анадыря, несмотря на его приморское положение, континентальные элементы флоры и растительности находятся в большей сохранности, чем на оз. Эльгыгытгын. Объясняется это низким положением суши в данном районе и более южным местонахождением. Поэтому реликтовая континентальная растительность более представлена в условиях менее континентального, хотя и менее сурового климата.
В «ботаническом содержании понятия «Южная Чукотка» (Юрцев, 1978) не приведено почти никаких специфических особенностей, отличающих эту часть Чукотки от континентальной ее части. Переходный характер флоры Южной Чукотки обусловлен, прежде всего, ее геосистемной структурой, часто на субстратном уровне, т. е. на одних участках (с одним субстратом) существуют, скажем, группировки, более характерные для континентальных условий, а по соседству (на другом субстрате) — для океанических условий. Идея выделения Южной Чукотки в том же ранге, что и западная, и восточная ее части, основана скорее на ее географическом положении, чем на достаточности ботанических показателей.
В пределах подзон кустарниковых и типичных тундр особенно броско отличаются орографические геосистемы, что нередко приравнивается к зональным отличиям и получает весьма неопределенные оценки. Такие сентенции, как «по характеру растительности участок (в среднем течении р. Амгуэмы, Ю. К.) отнесен к южному варианту подзоны северных гипоарктических тундр» (Катенин, 1974), являются типичным словоблудием, выдаваемым за достижение ботанической географии. Этот участок расположен на 200 км севернее пос. Конергино, растительность в окрестностях которого отнесена (Катенин, 1984) к «приморскому аналогу северного варианта подзоны северных гипоарктических тундр». Это район увалистой равнины. Севернее и восточнее его, в условиях горного ландшафта, вдоль рек существуют крупные массивы ивняков и, следовательно, представляют какойто иной зональный вариант (придумывать название которого мы не решаемся, т. к. для нас ясно, что дело тут не в зональности, а в различии орографических геосистем). По сравнению с подзоной кустарниковых тундр в типичных тундрах увеличивается мозаичность растительного покрова и, как следствие, возрастает количество видов в местных флорах. В пределах обеих подзон количество видов возрастает с запада на восток, достигая максимума в приморских районах.
|

Анадырский лиман. Начало чукотского лета.
|
Увеличение мозаичности растительного покрова обеспечивает плавный переход к подзоне арктических тундр, в которой исчезают ортотропные кустарники и становятся редкостью или также исчезают гипоарктические кустарнички (голубика, брусника, багульник, тощая березка). Главенство среди кустарничков приобретают аркто-альпийцы, главным образом дриада и Кассиопея. Степень развития растительности, отражающая мезо- и микроклиматические особенности, некоторые авторы рассматривают как признак зональных экосистем. Так, в Канадском арктическом архипелаге, на о. Виктория к низкоарктической экосистеме относятся сообщества с многочисленными низкими кустарниками и кустарничками (Dryas, Cassiope, Vaccinium, Rhododendron, Arctostaphylos), обилием трав; среднеарктическая экосистема менее разнообразна, особенно в отношении кустарников, доминирует дриада; в высокоарктической экосистеме — Dryas integrifolia, Salix arctica, Saxifraga oppositifolia (Edlund, 1983).
В подзоне арктических тундр основные типы растительности это — высокоарктический, горнотундровый, болотный, луговинно- нивальный; редкие типы (на о-ве Врангеля) — кустарниковый, степоидный. Климат подзоны полярноокеанический арктический с существенной дифференциацией на мезо- и микроклиматы лишь на о-ве Врангеля, природе которого посвящена целая серия публикаций (Городков, 1938; Сватков, 1958; Петровский, 1967, 1973 и др.). По мнению Б. Н. Городкова (1938), на границе аркти ческих тундр кустарники исчезают из-за растянутого периода стаивания снежных наносов, но это не главная причина. Кустарники не способны здесь существовать из-за температурных условий. Это доказывается состоянием кустарников на самой границе их существования, где они размещаются в наиболее теплых местах, но уже с пониженной жизненностью.
|
К. Брукс (1952) указывал, что Северный Ледовитый океан, будучи покрыт льдами, оказывает на температуру почти такое же влияние, как если бы он представлял собой продолжение Азиатского и Американского материков, а не водную поверхность. Однако В. Стефансон (1948) весьма убедительно доказал обратное, т. е. что океан влияет на температуру ближайшей суши и будучи скован льдом. Поэтому очень низких зимних температур, таких как в Центральной Якутии или в глубине бассейна Юкона, на северных побережьях не случается.
Когда высокоарктический тип растительности перестает сочетаться с другими типами (на арктических островах), оставаясь практически единственным, то эти территории некоторые исследователи выделяют в качестве зоны полярных пустынь. Согласно наиболее авторитетному автору, придерживающемуся этой точки зрения, В. Д. Александровой (1971), подземная часть фитомассы в тундровых сообществах (62—83%) больше надземной массы, а в сообществах полярных пустынь надземная фитомасса (78%) больше подземной (18%). Фитомасса цветковых растений в разных тундровых сообществах (78—91%) превышает фитомассу споровых (6—21% — мхи, лишайники — 1—-6%). В полярных пустынях обратные соотношения: цветковые — 23% всей фитомассы, 25% — мхи, 51% —лишайники. Эти данные являются яркой иллюстрацией различий двух типов растительности, но вряд ли их следует оценивать как зональный критерий, тем более что полярные пустыни представлены лишь в виде фрагментов суши — арктических островов или крайних выступов суши, т. е. территории, на которых экологический режим находится под контролем островного или полуостровного эффекта — общего нивелирования среды в неблагоприятном для развития растительности направлении. Там же, где площадь, приходящаяся на полярные пустыни, увеличена, например, на Земле Пири, на ней появляются весьма сомкнутые группировки растений, в которых соотношение надземной и подземной фитомассы не изучено.
В этой ситуации проявляется связь экологического потенциала территории с ее площадью. С увеличением площади при прочих равных условиях экологический потенциал увеличивается. Это следует уже из того, что большая площадь получает больше тепла и влаги. Соответственно на большей однотипной поверхности всегда более разнообразная растительность и богаче набор видов. Подобную же связь растительного покрова с площадью выделов можно наблюдать при высотной дифференциации растительного покрова. Если на одной и той' же не слишком большой высоте находятся две вершины, из которых одна конусовидная и имеет малую площадь, а другая плоская и площадь ее значительно больше первой, то первая будет относиться по растительности к холодным каменным пустыням в понимании В. Б. Куваева (1985), а вторая к типичным тундрам.
Методик количественного определения экологического потенциала местообитаний до сих пор не существует, хотя некоторые агрофизические приемы и приборы могли бы быть усовершенствованными для этой цели. Определение экологического потенциала способствовало бы решению многих проблем в изучении растительного покрова, в особенности причин его изменения и дифференциации на стыке зон и подзон. Простые наблюдения убеждают в том, что растительность одной зоны нередко встречается в виде островов глубоко в пределах другой зоны. Следовательно, в пределах этих островов среда не слишком существенно отличается от той, в которой эта растительность является фоновой. Это подобие складывается из свойств геосистем разных рангов. Например, существование Телекайской рощи, представляющей лесной остров в подзоне кустарниковых тундр, обязано свойствам орографической геосистемы (межгорной котловины с низовьями Левого Телекая), геоморфологической геосистемы (многорукавной системе самой реки), субстратной геосистемы (опесчаненные галечники, местами задерновелые). Решающее значение в данном случае имеет орографическая геосистема, однако без комплекса ее с двумя другими рощи в этом месте не было бы. Таким образом, экологический фон существования рощи слагается из нескольких составляющих, но фактически она расположена в пределах только определенной субстратной геосистемы и к тому же не заполняет ее полностью.
Субстратные геосистемы в тундровой зоне имеют огромное значение из-за низкорослости растений. Некоторые из них зависят от геосистем более высокого ранга, другие автономны. Поэтому сначала остановимся на принципах выделения и характеристике субстратных геосистем, а уж затем приведем примеры других геосистем, которые, хотя и составлены субстратными, не являются арифметической суммой последних, а приобретают специфические качества, которые, в свою очередь, способны оказывать влияние на субстратные геосистемы.

