Экофон
Действие среды на растения
Экология изучает взаимодействие живых организмов и среды их обитания, которая может быть представлена в виде двух составляющих: биогенной и абиогенной (или абиотической). В умеренных широтах и отчасти в субарктике биогенная и абиогенная составляющие представляют настолько цельный комплекс, что их раздельное рассмотрение едва ли возможно без ущерба для выяснения существующих корреляций между растениями и средой. В лесах попытка такого разграничения вообще теряет смысл. Здесь мы имеем взаимопроникновение двух сущностей примерно такого же порядка, как, скажем, в растворе поваренной соли. Частицы этой соли в водном растворе существуют лишь в потенциальной связи. Раствор в нормальных условиях представляет целостную динамическую систему с равновесным взаимодействием трех видов частиц. Убирая среду (выпаривая раствор), мы изменяем свойства ингредиентов — образуется кристаллическая решетка. Система «раствор» теряет свою целостность; мы не можем рассматривать кристаллы соли по отношению к раствору, поскольку он уже не существует. Образовалась новая система.
Взаимодействие растительности с косной средой весьма аналогично. Поэтому совершенно правы Н. Д. Ярошенко (1953) и Е. П. Гусельников (1955), считая, что не стоит разделять понятия «местообитание» и «экотоп», поскольку местообитание является довольно целостной системой, обладающей спецификой обмена веществом и энергией. Живая среда пронизывает неживую, вызывая в ней изменения, которые могут быть значительными, хотя и не улавливаемыми нашими ощущениями. С такой обстановкой мы сталкиваемся в лесных зонах, отчасти в Субарктике и даже в Арктике. Но в Арктике биогенная составляющая в значительной мере утрачивает свое значение в общем балансе среды и главная роль отводится абиогенной составляющей. Эта мысль была высказана еще Ч. Дарвином (1859, по изд. 1909), а в дальнейшем развивалась целым рядом исследователей. В частности, Б. Н. Городков (1946) был полностью согласен с мнением Дарвина: «В наиболее суровых климатах возникли упрощенные растительные сообщества, в которых борьба за существование стала протекать в форме непосредственного противостояния стихиям». Н. Полунин (Polunin, 1960) обратил внимание на то, что подобная ситуация должна рассматриваться как частный случай, хотя и очень распространенный. Действительно, оценка конкурентных отношений в Арктике должна быть дифференцирована, в частности, по крупным формам рельефа, для каждого из которых наиболее характерен какойто один растительный тип. Моховые тундры на равнинах Чукотки доходят до северного побережья, а в такого типа тундрах биогенной составляющей среды принадлежит существенная роль, которая еще более усиливается в южной Субарктике. Однако в горных условиях Субарктики, так же, как и в высокой Арктике (полярных пустынях), биогенная составляющая среды местами сходит на нет. Недаром Н. Полунин (Polunin, 1960) отметил, что «in the Arctic the vegetation is relativety impotent».

Трасса Эгвекинот - Иультин на перевале хр. Искатень. Акварель Ю.П. Кожевникова.
Взаимодействие биогенной и абиогенной составляющих среды (или в предельных случаях только абиогенная составляющая) определяют условия произрастания в данном районе некоторого числа видов растений, наличие которых обусловлено особенностями флорогенеза и современными условиями обитания (Salisbury, 1939; Mason, 1946, 1954; Толмачев, 1959). Согласно Толмачеву (1959), «историческая обусловленность состава флоры — это зависимость его от географического происхождения и возраста определенных групп растений и конкретных видов, от хода их расселения в минувшие эпохи, часто под влиянием условий, существенно отличающихся от современных, имеет первостепенное значение. Состав флоры страны не представляет прямого отражения влияний современных условий, но до известной степени автономен от них» (подчеркнуто мной.— Ю. К-). В настоящее время это представление является общепринятым. Однако правомерно может быть поставлен вопрос — до какой степени состав современной флоры автономен от современных условий.
Квалифицированный географ растений может весьма правдоподобно представить себе климатическую обстановку в непосещавшемся им районе на основе изученного списка растений, обитающих в этом районе (Boyko, 1955). И действительно, соотношение географо-генетических элементов флоры весьма коррелирует с современной обстановкой. Это подтвердил А. В. Галанин (1974) при анализе географических элементов двух КФ (континентальной и океанической) на Чукотке с применением метода хи-квадрат. Уже в поле флорист получает информацию о том, что влияние современных условий на флору велико. Так, при исследовании района, переходного в отношении климатов, мы нередко сталкиваемся с единичными находками дифференциальных (нахо дящихся на пределе географического распространения в данном районе и занимающем специфические стации) видов, к тому же в угнетенном состоянии. Можно лишь догадываться, что часть подобных видов выпала из состава флоры совсем недавно. Параллельное суждение касается «свежих» иммигрантов, которые обладают противоположной жизненностью. Переживание реликтов такой же результат современных условий, как и активизация иммиграций с территорий смежных областей. Количество реликтовых видов по флоре, очевидно, зависит от быстроты и качества изменяющейся среды в ходе истории. Такой взгляд, однако, не отрицает некоторого консерватизма флоры по сравнению с растительностью, перестраивающейся с изменением условий более быстрыми темпами. Не следует лишь преувеличивать тот временный интервал, который разделяет изменения во флоре и растительности адекватно изменениям среды. По наблюдениям в основании Чукотского полуострова количество реликтовых видов в местных флорах, по-видимому, зависит от соотношения биогенной и абиогенной составляющих среды на данной территории. Чем больше в исследуемом районе элементарных ландшафтов с преобладанием абиогенной составляющей, тем больше в нем реликтовых видов. Это связано, очевидно, с отсутствием конкуренции на таких элементарных ландшафтах, на что указывал Григгс (Griggs, 1940). Исследуя особенности поведения, современных реликтов, нельзя не сделать вывод, что, только проецируя экологическое распределение видов на их географическое распространение, мы имеем основания для надежных исторических интерпретаций.
В основании Чукотского полуострова установлено, что роль биогенной составляющей среды обитания усиливается параллельно возрастанию континентальности климата. В континентальных районах заметно увеличивается фитопродуктивность ландшафтов, возрастает стабильность в строении растительного покрова. В океанических районах растительный покров обычно не имеет столь четкой структуры, как в континентальных. К аналогичному выводу пришел Э. Хультен (Hulten, 1960), сравнивая растительность Алеутских островов и Камчатки. Растительность Алеутских островов представляет несколько обедненный дериват растительности горной Камчатки. Но на Камчатке горная растительность имеет стабильную структуру, тогда как на Алеутских островах она «расплывчата». Это объясняется большей океаничностью климата Алеутских островов.
Взаимодействие абиогенной и биогенной составляющей должно изучаться в географическом и экологическом отношениях. При экологических исследованиях должны быть предприняты попытки отыскания количественных критериев для оценки этого взаимодействия. Хорошим объектом для исследования взаимо действия абиогенной и биогенной составляющей среды являются пятнистые тундры. При стационарных исследованиях в них легко могут быть проведены статистические исследования по этому взаимодействию. Как указывал Б. А. Тихомиров (1957), голые тундровые пятна представляют собой абиогенную среду. Вокруг пятен, в условиях сплошной дернины уже велика роль биогенной составляющей среды. Пятна в разной степени зарастания характеризуются промежуточными отношениями абиогенной и биогенной составляющих среды.
В литературе существуют данные, что фитомасса положительно коррелирует с продуктивностью растительности. А продуктивность является показателем интенсивности ландшафтных процессов (Хомченко и др., 1974). Фитомасса тесно связана с размерами растений, т. е. с формами роста видов; создающих фитомассу. Этим объясняется то, что низкая фитомасса образуется при обилии мелких растений (трав), а высокая — немногими крупными (деревьями) ( Bell, 1982). Имея даже качественные сравнительные данные о фитомассе (здесь она больше, там меньше), мы можем говорить о сравнительной фитопродуктивности и интенсивности ландшафтных процессов.
|

Палаточный лагерь ботаников на Чукотке в 70-е годы ХХ века. Фото А.А. Коробкова.
|
На основе взаимоотношений растений и среды создано уже очень много спекулятивных теорий, нередко взаимоисключающих и основанных притом на одних и тех же фактах. Такое положение обязывает к более пристальному исследованию свойств современной среды. Нельзя не согласиться с мнением Л. Н. Со болева (1961), что «поскольку растительный покров представляет собой сочетание растений, обусловленное факторами среды (как современными, так и воздействовавшими в прошлом), его разделение на экологически обусловленные категории имеет первостепенное значение». Приведенная цитата совершенно не случайно согласуется с нашими представлениями, т. к. и в том и в другом случае объектами ботанического исследования были открытые ландшафты. Поэтому, несмотря на различия, обусловленные региональными климатами, могут быть использованы одни и те же методы исследования, или, по крайней мере, общие идеи, касающиеся понимания связи растений со средой. Еще в 1929 году Дю Рие (De Rietz) писал, что настало время исследования степени корреляции между местообитаниями и их растительностью.
Гористая Арктика имеет особые преимущества для исследования экологических режимов в разных геосистемах, поскольку биогенная составляющая в одних случаях сведена к нулю, в других играет существенную роль. Изучение среды обитания следует проводить независимо от наличия индикаторных видов. Н. Полунин (Polunin, 1960) писал, что «вместо физической аппаратуры мы можем использовать |
«фитометры» — определенные растения или куски дернины, переносимые в другие условия, которые интегрируют все действующие факторы среды и выражают результат своими реакциями». В другом месте этой же книги Н. Полунин уже не склонен к обобщениям широкого плана: «Эти «индикаторные» растения, сообщества и другие признаки склонны варьировать от одного района к другому, иногда даже с сопоставимым (сравнимым) климатом, так что для использования принципа индикации необходимо солидное знание поведения видов, единственный выход из такого положения - детальные местные флористические и экологические исследования». Это высказывание Полунина не отрицает существование видов с определенной индикаторной ролью, но является предостережением против чрезмерно широкого использования растений в качестве индикаторов. Равным образом, роль растительных сообществ как индикаторов определенного комплекса физических условий не должна переоцениваться, особенно при сравнении разных географических районов с несходным ландшафтом. Так, остепненные группировки в континентальных районах Чукотки занимают, как правило, строго определенные местообитания на южных склонах, но случаются и исключения, и тогда предположения о главенствующей роли температурного режима становятся зыбкими, поскольку отсутствуют конкретные данные о температуре. Место такого «выбившегося из правила» сообщества в ландшафте отнюдь не располагает к предположению о большем количестве тепла на данном местообитании в сравнении с другими без остепненных сообществ. Так, Б. А. Юрцев (1967) обратил внимание на «остепнение сухих лугов на высоких песчано-галечниковых гривах» в поймах близ п. Баранихи (Западная Чукотка). «Остепнение» заключается в присутствии Сагех spaniocarpa, С. obtusata, Potentilla arenosa, Alyssum biovulatum и др. видов. Однако никаких данных о собственно остепнении, представляющем собой физико-географический процесс, этот автор не приводит. Поскольку подобных примеров существует множество, то следует сделать заключение, что индикаторная роль как отдельных видов, так и их сообществ имеет некоторую амплитуду колебаний по приуроченности к условиям среды, пределы которой не могут быть установлены по показаниям самих растений.
Любой участок земной поверхности может быть представлен в виде совокупности множества физических и химических факторов, взаимодействующих между собой на основе прямых и обратных связей и являющихся открытой системой. Взаимодействие факторов таково, что при изменении одного из них происходит изменение и других, причем направленное на устранение нарушения общего энергооборота, вызванного изменением первого фактора, т. е. происходит компенсаторный эффект ( Rubel, 1935; Daubenmire, 1947; Odum, 1959; Polunin, 1960 и др.). Компенсаторный эффект на участке земной (или водной) поверхности является функцией поддержания динамического равновесия (в отличие от живых систем с динамическим неравновесием) в соответствии с принципом Ле Шателье. Можно также сказать, что каждый участок земной поверхности обладает гомеостатическими свойствами, хотя и в несравненно меньшей степени, чем это имеет место в живых организмах. Уровень гомеостаза повышается с увеличением рассматриваемой площади.
Из этих соображений вытекает вывод, что комплекс абиогенных факторов действует на живые организмы как единое целое или холоценотически (Cain, 1944; Майр, 1947; Dansereau, 1957; Good, 1964). В западной, в том числе американской, литературе точка зрения о холоценотическом действии среды на организмы приобрела широкую популярность. Недаром Ф. Эглер весьма едко критиковал книгу Добенмайра «Растения и среда». «Этот однофакторный усредненный каузализм среды является спекулятивной философией, на которой аутэкология (environmental autecology ) ненадежно сбалансирована» (Egler, 1951). По поводу того, что Р. Добенмайр (Daubenmire, 1947) сводит действие комплекса факторов в единичном примере (против множества «однофакторных» примеров) опять к эдафическому фактору ф. Эглер восклицает: «Однофакторная экология психологически столь удовлетворительна!». Дю Рие. ( Du Rietz , 1929) уже давно высказал мнение, что традиционное разделение факторов местообитания на эдафические, климатические, биотические, исторические и т. д. является нелогичным, т. к. эти группы не исключают друг друга при изучении местообитания, а взаимосвязаны и, более того, взаимопроникающи.
Надо сказать, что вопрос о комплексном действии среды заострялся еще в начале нашего века. Так, Брокман-Ерош, исследовав пределы распространения деревьев и черты климата у этих пределов, пришел к выводу, что лесные границы зависят от комплекса климатических факторов (Tansley, 1920; Griggs, 1934), а не от какого-либо одного, так называемого ведущего, фактора. Несмотря на этот вывод Брокман-Ероша, еще несколько десятилетий, вплоть до наших дней, делались попытки увязать естественные границы распространения древесных пород с каким-либо одним главенствующим фактором. Это с необходимостью приводило к допущению, что в разных районах Земного шара распространение деревьев лимитируют разные ведущие факторы.
В 1962 г. Ф. Ф. Давитая и Ю. С. Мельник сообщили, что полярная граница леса на Кольском полуострове и верхняя граница леса на Кавказе «соответствуют .практически одинаковой сумме температур поверхности растительного покрова» (Будыко, 1971). Очевидно, главенствующим фактором природных лесных границ является количество радиации, хотя с холоценотической точки зрения это вновь означает «однофакторный каузализм». Однако сказанное о холоценотическом действии среды на организм вовсе не отрицает и того, что в ряде случаев в комплексе факторов среды имеется один основной фактор, необходимый для существования некоторых организмов (например, реликтов на известняках, близ выходов горячих вод и т. п.) или ответственный за узкую экологическую специализацию, связанную с адекватными среде морфологическими изменениями организмов (например, в случае приморских галофитов или шиловиднолистных криоксерофитов). Однако среди любых хорошо объяснимых явлений подобного рода мы находим, что «однофакторное» объяснение недостаточно. В случае реликтов весьма правдоподобным будет объяснение с использованием понятия компенсаторного эффекта или гомеостаза, который включает взаимодействие факторов. В случае узкой экологической специализации к одному фактору также потребуются дополнительные объяснения. Например, приморские галофиты приспособлены не только к засолению, но и к сильным ветрам и к рыхлому субстрату (на галечниках и песках) или, напротив, к тяжелому (в лайдах), а также к вмерзанию в лед, действию нагонных волн и т. п., т. е. всему комплексу условий обитания в приморской полосе.
Только полный учет особенностей среды может дать четкое понимание различной экологии специализированных групп растений, т. к. два выше приведенных наугад примера имеют то общее, что обе группы растений являются ксерофитами. П. Данзеро (Dansereau, 1957) указывает на этот счет, что «во всех случаях морфология растений и среда обитания не могут быть без риска выведены одно из другого и должны различаться и изучаться раздельно». Е. Рюбель (Riibel, 1935} считал, что значение «фактора в минимуме» для организмов всегда ниже в совокупности прочих факторов за счет их компенсации того критического значения «фактора в минимуме», который был бы гибелен для организма, например, в неестественной среде (скажем, в культуре).
Е. Сэлисбэри, изучавший явление кальцефилии писал: «Проблема - что определяет предпочтение карбонатных почв кальцефилами — отчетливо сложная, и неправильно допускать объяснение на основе одного единственного фактора» (Salisbury, 1920). Далее этот автор указывает, что в одной местности встречаются кальцефилы, которые растут не только на карбонатных почвах, но и на таких, которые лишь напоминают карбонатные, будучи хорошо аэрированными и сухими, но не содержащими кальция. В данном случае для растений более важны физические свойства субстрата, чем химические. Об этом же писал Е. Рюбель (Rubel, 1935). Кальций является одним из наиболее широко распространенных элементов, необходимых для растений. Он есть всюду. По данным Б. В. Польтнова (1956), в области распространения гранитов химический состав сухого остатка вод содержит: 17,33 мг % SiOs и 11,89 мг % Са, а в области распространения базальтов эти же составляющие представлены соотвественно 7,67 мг % и 20,07 мг %. Однако и в том, и в другом случае, как указал Б. В. Полынов, карбонаты преобладают над кремнеземом (в области гранитов — 30,49 мг %, в области базальтов — 46,85 мг %). Но и на гранитах, и на базальтах растительность не является кальцефильной, следовательно, для последней требуется более высокое содержание карбонатов и притом кальция. Поскольку содержание кальция в почвенных растворах связано с величиной рН, то определение последней, хотя и неоднозначно, свидетельствует о степени тяготения растений к кальцию. На юго-востоке Чукотского полуострова было выделено 6 групп растений по их отношению к рН (Баландин, 1982). Фактически растения образуют непрерывный ряд этого отношения и число групп является произвольным. Большинство растений предпочитает нейтральную реакцию почво-грунтов (Small, 1954), т. е. умеренное содержание в почве кальция.
Р. Гуд (Good, 1964) приводит случай, когда влияние минеральных составляющих почв производит морфологические изменения одних растений и является фактором реликтовости других. Это — явление серпентиноморфоза, проявляющееся среди некоторых растений, обитающих на скалах серпентина. В числе морфологических изменений: отсутствие опушения, наличие воскового налета и карликовые размеры. Имеется три типа растений: облигатные серпентинофиты, предпочтительные серпентинофиты и серпентинофиты реликты. На Чукотке эти группы выделяются по отношению к известняку, морфогенетическое влияние которого также прослеживается, например, подушковидная биоморфа Oxytropis nigrescens ssp. bryophila (O. gorodkovii) скорее всего сформировалась именно на известняках; не исключено, что на них же возникла Artemisia senjavinensis. Случаи, подобные только что приведенному, относятся к категории явлений, названных Дансеро (1966) законом факториального контроля по Del Villar: «Хотя все живые существа реаги руют холоценотически (на все факторы среды в их специфической совокупности), часто имеется особый фактор, который имеет силу контроля при его избытке или недостатке».
Комплементарнооть биогенной и абиогенной составляющих среды
Если рассматривать среду обитания растений вообще, то можно прийти к кажущейся тривиальной истине, заключающейся в том, что подавляющее большинство растений имеет контакт со средой в двух направлениях: их надземные части находятся в воздушной среде, а подземные в почве, так что факторы среды могут быть разделены на климатические и эдафические (Good, 1964). В том же духе высказывался Л. Г. Раменский (1971) в отношении биоценоза: «биоценоз находится в непрерывном взаимодействии с почвенной средой (эдатопом) и воздушной (климатопом)». Дю Рие (Du Rietz, 1929) также различал «местообитание образующие факторы» двух категорий — климатические и эдафические. При этом он указывал, что дождевая влага и снег являются климатическим фактором, пока находятся в воздухе; упав на землю, они становятся эдафическим фактором. Развивая мысль об этих двух средах и отношении к ним растений, можно высказать суждение, что надземные и подземные части растений до некоторой степени развиваются автономно (но по принципу комплементарности), поскольку для каждой из них, в сущности, неважно, каким образом устроена комплементарная бисистема, лишь бы она функционировала согласно требованиям целого организма. В отношении такого подхода вообще к живым организмам сошлемся на мнение Р. Л. Берга (1964), с которой мы полностью согласны: «Живой организм рисуется как система, стоящая из отдельных подсистем, обладающих своими показателями устойчивости (нормой реагирования). Каждая из би- или эдсистем взаимодействует с определенным комплексом внеших условий». Весьма характерно, что для одних видов первостепенным является климатоп, для других эдатоп и для третьих биотическое окружение. В соответствии с этим различают климатические, эдафические и биотические экотипы (Розанова, 1946).
|

Гусеничный вездеход в чукотской тундре, начало 70-х годов ХХ века.
Фото А.А. Коробкова. |
Конечно, речь идет лишь об определенном уровне противопоставления, т. е. климатоп может быть противопоставлен эдатопу лишь до некоторого предела, поскольку между обоими происходит тесное взаимодействие. Еще большая степень интеграции свойственна растению (его подземной и надземной частям). И в том и в другом случае имеем комплементарность двух подсистем, разграниченных земной поверхностью. Однако понимание комплементарности неотделимо от целостности комплементирующих частей (дополняющих друг друга). Поэтому для анализа среды как целого необходимо учитывать и климатические, и эдафические показатели.
Поскольку факторов среды существует огромное число, рационально определить наиболее ответственные из них, т. е. те, через которые выражаются все другие показатели (которые, так сказать, «вмещают» в себя остальные показатели, оказывающие влияние на учитываемые). Такие интегральные показатели могут быть определены и для климатопа, и для эдатопа. Представляется, что отношение интегральных показателей условий климатопа и эдатопа (их индекс) может использоваться в качестве относительной оценки абиогенной среды, а этот показатель может служить для изучения соотношения абиогенной и биогенной составляющих среды. В нашем случае такой индекс был бы полезен для изучения распределения видов и флористических групп по местообитаниям на территории КФ, особенно в тех случаях, когда |
биогенная и абиогенная составляющие среды могут безболезненно рассматриваться раздельно, а это, как уже говорилось, весьма распространенное явление в Арктике. Такие же возможности имеются и в других районах Земли — всюду, где нет сомкнутой многоярусной растительности.
Сравнение территории конкретных флор (КФ) значительно выиграло бы, если бы мы могли сопоставить индексы среды на сходных местообитаниях разных территорий (пусть не на всех, а лишь наиболее характерных), а затем уже оценивать различия в растительном покрове на тех же местообитаниях. Трудность заключается в отыскании удачных выражений интегральных показателей среды, но, главное, в практическом отсутствии необходимых данных по режимам отдельных факторов на разных местообитаниях. Разумеется, такая работа может быть выполнена только на стационарах, и мы говорим здесь об этом лишь как о принципиальной возможности. Если рассматриваемая проблема будет разрешена даже для какого-либо одного района, это послужит стимулом для дальнейших исследований в этом направлении. Интегральные индексы могут быть получены эмпирическим путем, когда будет оценена амплитуда колебаний отдельных факторов среды и характер их связи между собой. По всей вероятности, значения индексов могут иметь только региональное значение (возможно, в пределах физико-географической провинции), поскольку сходные климаты различных регионов имеют различное экологическое значение (Сочава, 1971).
Принцип комплементарности был использован А. Л. Тахтаджяном для определения «связи между топами в рамках надфациальных структур» (Крауклис, 1974). Однако в этом смысле применение данного принципа неточно, поскольку он касается лишь тех структур, которые неделимы без утраты целостности, но в то же время их целостность создается благодаря взаимодополняемости некоторых составляющих. Так, ДНК существует только при условии взаимодополняемости двух тяжей нуклеотидов. Отношение двух смежных топов — это отношения взаимодействующего соседства, но не комплементарности. Напротив, принципу комплементарности отвечают надземная и подземная части растений, а также эдатоп и климатоп, поскольку фация не существует без этих составляющих.
Некоторые данные о температурном факторе на западе Чукотского полуострова
При географическом сравнении разных пунктов берутся данные метеостанций и сопоставляются цельные флоры или структуры растительного покрова. Эти данные относятся к местному климату того типа местности, в котором располагается метеоплощадка. Они могут быть репрезентативными, если метеоплощадка заложена в типических условиях местности (Хромов, 1952), но последнее не является обязательным. Данные метеостанции могут существенно не соответствовать уже ближайшим орографически различным территориям. Однако при географическом сопоставлении участков растительного покрова такие данные оказывают существенную услугу. Подчеркиваем, что использование данных метеостанций является в сущности географическим методом, а не экологическим, как это, например, расценивает С. Янг (Young, 1971). При экологическом сравнении различных местообитаний по микроклимату и растительности или набору видов могут использоваться те же приемы, что и при географическом сравнении. Здесь имеет место перекрытие географических и эколог ических аспектов изучения распространения и распределения растений. Это же перекрытие проявляется в использовании метода климадиаграмм, где на оси ординат откладываются интервалы температур, а на оси абсцисс — интервалы количества осадков, в поле графа показывается область гидротермических отношений, освоенная каким-либо видом (Odum, 1959). Метод изоплет Л. Г. Раменского является экологическим вариантом географического метода климадиаграмм.
|

Провиденские горы. Кар и троговая долина разработаны ледником. Фото А.А. Галанина.
|
Таким образом, и в глобальном масштабе, и в пределах постепенно уменьшаемых территорий (вплоть до физико-географической фации) распределение растений наитеснейше связано с распределением тепла и влаги. Коснемся несколько экологического аспекта температурного фактора, поскольку в любое время температура участка земной поверхности определяет климат близ ее в наивысшей степени (Geiger, 1957). В настоящее время имеется мало опубликованных данных по температуре растений обитаемого слоя биосферы в Арктике, а по Чукотке их почти нет. Поэтому любые дополнительные данные представляют большой интерес, даже в том случае, если это несистематические, в значительной мере случайные, данные (Хромов, 1968). Такие данные, на необходимость которых указывал и А. А. Григорьев (1966), имеют не только познавательную ценность, но позволяют также сделать, некоторые экологические обобщения, ориентировать дальнейшее исследование термального режима, в частности, при нестационарных исследова ниях.
Здесь приводятся данные, полученные по ходу маршрутных исследований растительного покрова в некоторых пунктах на западе Чукотского полуострова. Наряду с измерениями обычным ртутным термометром (почвенным), использовались также самописцы, помещаемые на контрастные местообитания. В литературе не раз отмечалось существование резкого температурного градиента близ деятельной поверхности (например, Bocher, 1949; Шамурин и Тихменев, 1971). М. И. Будьгко (1971) писал, что различия в температуре по вертикали в околоземном слое воздуха соответствуют различию средних температур пунков, удаленных друг от друга на тысячи километров. В тундрах это различие особенно велико благодаря свободной циркуляции воздушных масс в открытых ландшафтах. Околоземные и почвенные температуры наиболее тесно связаны с мезоклиматом отдельных подурочищ, который, в свою |
очередь, определяется условиями формирования климата урочища. В каждом конкрет ном случае эти условия весьма своеобразны, хотя известны и некоторые общие закономерности. Так, например, температурная инверсия на восточных склонах всегда больше, чем на западных. Микроклимат более высоких уровней характеризуется большим количеством дневного тепла и меньшей потерей этого тепла ночью (Geiger, 1957), В отношении почвенного микроклимата известно, что даже «экспозиция нанорельефа оказывает существенное влияние на микроклимат верхнего, наиболее корнеобитаемого горизонта почвы» (Александрова, 1960). Этим же автором установлено на о-ве Большом Ляховском, что «различия в почвенном микроклимате сказались не только на ритме и степени развития отдельных растений, но и на составе видов, произрастающих на юго-западном и северо-восточном склонах одной и той же кочки».
В каждой данной местности температуры прежде всего зависят от погоды и топографии. Как и многие другие факторы среды, температура является очень динамичным показателем, связанным очень тесно с влиянием других факторов, в частности ветра. В солнечную погоду с порывистым ветром в начале июля на южном склоне сопки близ зал. Свободный отмечено, что температура колеблется от 19° (затишье) до 16° (порыв) за 1 мин. О нестабильности максимальных дневных температур говорят «дрожащие» максимумы на лентах термографов и минимумы на лентах гигрографов. При приближении к 40° кривая термографа «пляшет» с амплитудой до 10°, совершая до трех таких колебаний за два часа. В пасмурную или ясную, но с сильным холодным ветром, температуры выравниваются и в горизонтальной, и в вертикальной плоскостях. Согласно Зиегенталеру (Geiger, 1957), коэффициент корреляции между температурами почвы на глубине 10 см и приземного воздуха равен +0,87.
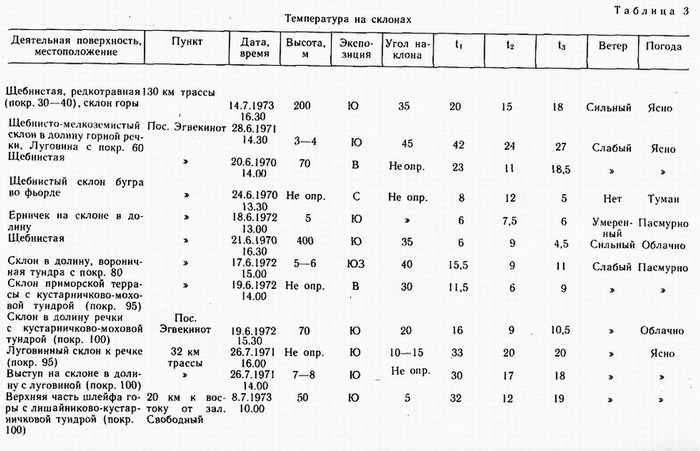
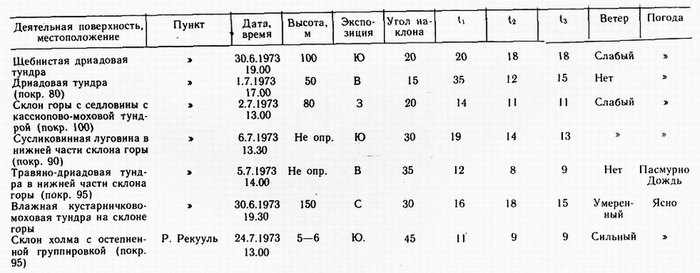
В солнечную погоду степень нагрева деятельной поверхности зависит от экспозиции и угла наклона и практически не зависит от наличия или отсутствия растительности. Но растительность весьма чутко реагирует на нагрев, т. е. наблюдается практически односторонняя связь. В укрытых местечках t достигает 42° (табл. 3), a t свыше 30° не является редкостью в июне и июле. В Эгвекиноте t 35° была зафиксирована 20.06.1970 на вершине сопки 550—600 м в 15 ч на щебнистом субстрате с дриадовой тундрой. Однако в подобном местопложении сильный нагрев поверхности не оказывает существенного влияния на характер растительности. Во-первых, это очень спорадическое явление на вершинах; во-вторых, здесь действует комплекс факторов, в котором
Для краткости в таблице приняты следующие обозначения: ti — температура деятельной поверхности; t 2 — температура субстрата на глубине 3—5 см; t 3 — температура воздуха на высоте 1 м.
|
|
температура не является решающим фактором. Летом верхние части гор обычно обдуваются сильными ветрами, и температура поверхности часто низкая (3—8°) или умеренная (10 —15°), поднимающаяся на несколько градусов в малейшем укрытии. Напротив, внизу в укрытых местоположениях влияние нагрева проявляется: 1) во флористической обогащенности, 2) в отличной жизненности отдельных растений, 3) в сравнительно раннем наступлении фенофаз, 4) в физиономии растительности, отличающейся пышностью. В районах субконтинентального климата запада Чукотского полуострова для подобных местоположений характерны также низкокустарниковые (а не кустарничковые) заросли березки, голубики, багульника. Такие сообщества играют существенную роль в резко континентальных районах (например, в верховьях Телекая), с чем ассоциируются многие другие ботанико-географические различия этих районов и субконтинентальных.
Растительность оказывает влияние на почвенный температур ный режим, что отчетливо прослеживается в пятнистых тундрах. Замеры в разных местах одной подушки диапенсии показывают некоторые различия (15—16°). Здесь же t 2 голого суглинистого сухого пятна колеблется в разных точках на 1° ниже (14 —15°), В пятнистой дриадовой тундре на Ю—3-склоне, 25° близ Рекууля (Ванкаремская низменность) 24.07. 1973 в 14.30 отмечены для дриадовой полосы ti 25,5°, 1 2 10,5°; для голого пятна t ,'25°, t 2 14,5° при t 3 20°. В этом же районе, на поверхности высокой надпойменной террасы с кустарничково-моховой тундрой с криогенными голыми пятнами (в 13 ч. 30 мин. 25.07.1973): для дернины t , 12, t2 8,5; для голого пятна t , 12, t 2 13 при ts 13. Можно привести еще ряд примеров, но все они свидетельствуют о том, что при неизменной микротопографии температурные различия почв в комплексных тундрах обусловлены растительностью. Однако достоверное различие заключается лишь в том, что растительность задерживает тепло от проникновения вглубь. Эта задержка обусловлена в ряде случаев расходом тепла на испарение в моховых вариантах тундр. При t |
торфянистой дернины 15,3°, а голого пятна здесь же 15,6°, температура гальки под дерниной мощностью 15 см (на рыхлой поверхностной толще близ зал. Свободный) была всего 3°, при t 12°, a t2 8,8°. При невысоких t3 (8—12°), в болотах лишь незначительно выше температуры воздуха, но с повышением последней ti в болотах резко повышается. На влажных речных аллювиях это не наблюдается. Даже при t3 20—23° ti влажного аллювия превышает эту величину только на 3—4° или вовсе не превышает.
Р. Гайгер писал: «Что касается микроклимата близ поверхности земли, то эта поверхность сама действует как регуляторный резурвуар тепла» (Geiger, 1957). Далее Р. Гайгер указывает, что чем больше термальная проводимость почвы (грунта), тем она более эффективна в качестве резервуара тепла. Различие температур поверхности голых пятен и окружающей их дернины является весьма постоянным. Оно приводит к более глубокому протаиванию мерзлоты под пятнами по сравнению с дерниной (Тихомиров, 1957). Нагрев торфяной поверхности также зависит от экспозиции и угла наклона, не уступая количественно нагреву аналогичных по местоположению щебнистых субстратов. В парковом кустарничково-моховом ольшатнике в среднем течение Амгуэмы (130 км трассы) в 14.30 14.07. 1973 было: t , — 23°, t 2 —15°, t 3 — 20°.
В последнем примере ярко проявляется влияние на температурный режим микротопографии. В ясную погоду это влияние охватывает весь ландшафт, т. е. любые формы макрорельефа и растительные формации. В кочкарниках при ti кочки 11 —16°, между кочек t4—8°, а t3 10—16°, т. е. температура поверхности кочки часто незначительно превышает t3 , а между кочек t составляет половину t кочки. Одиночные замеры в Баранихе и Певеке свидетельствуют, повидимому, о том, что в континентальных районах Чукотки отношения иные. Разница между t , и t2 не столь, велика и, напротив, tj существенно превышает t3. Устойчивость: таких отношений в связи с преобладанием ясной погоды и повышенных tg приводит к пересыханию многих болот на шлейфах гор и на равнинах и их легкому воспламенению (табл. 4). В субконтинентальных и приморских районах подобные болота не пересыхают, т. к. значительный нагрев поверхности болот случается не часто. С моря регулярно приходит низовой туман. На Чукотском п-ове отмечена более сложная вертикальная структура микрогруппировок на вершинах пушицевых кочек по сравнению с междукочиями (Кононова, 1978). Немалое значение в этом различии имеет дифференциация температур.
В восточноевропейской лесотундре на поверхности сухого мохового покрова А. В. Друзин (1967) отмечал максимальную температуру 46°, а на влажной поверхности — 35—36°. По данным этого автора в пятнистой тундре амплитуда колебаний температур за лето больше в западине (52°), чем на минеральном пятне (36°), но в кустарниковой тундре, в березовом и еловом редколесьях эта амплитуда больше на бугорках, чем в западинках. На Чукотке такие высокие температуры поверхности почвы и такие значительные их амплитуды не достигаются.
На сухих горизонтальных поверхностях, например, надпойменных террасах выс. 7—8 м, с горными вариантами тундр и волнистым микрорельефом отношения ti к ta в солнечную погоду таковы. Близ зал. Свободного в начале июля ta 11 —13—18°. При этом ti на взлобке с лишайниковой тундрой 14—17—25°, а в ложбинке с дриадовой или арктоусовой растительностью (с покрытием 80—100%) 13—19—26°. Соответственно t2 в первом, случае 13—14(16) —17°, во втором—10—12—9°. Флористическое различие взлобка и ложбинки несущественно, но геоботаническое огромно. Однако вызвано оно не различиями в температурах поверхностей, а целым комплексом условий, включающим сдувание снега зимой со взлобков и «законопачивание» им ложбинок, накапливанием мелкозема и лучшим увлажнением в ложбинках и пр. Следует сказать несколько слов о термальной роли снежников. Как хорошо видно в резко континентальных районах, само существование снежников зависит от особенностей местного климата, в частности, от температурного режима. В этих районах снежники в нижних частях рельефа нехарактерны (Bocher, 1949; Кожевников, 1974а, б, в). Субконтинентальный и морской климаты характеризуются обилием снежников по всему вертикальному ландшафтному профилю. Многие из них сохраняются до второй половины лета, а часть перелетовывает.
Было установлено, что снежник вносит очень незначительное изменение в температуру околоземного слоя воздуха. Наши наблюдения соответствуют данным Р. Гайгера (Geiger, 1957), который отметил, что на высоте нескольких сантиметров над тающим снегом температура может достигать 10° в полдень. Наиболее интересным является тот факт, что самый верхний слой снега тающего снежника может иметь температуру 1°. Очевидно, это успевает нагреться талая вода, прежде чем она просочится в глубь снежника. В Норвегии Н. Kevin (I 982), изучая температуры нивальной ниши, сделал предположение, что замерзание происходит не при переходе через 0°, а при более низких температурах.
В особенно теплые дни, когда близ поверхности кочкарника на южном шлейфе сопки близ зал. Свободного достигала 34°, а на высоте 5 см над снежником вдоль склона высокой надпой менной террасы — 17° (08.07.1973). Близ Ванкаремской сопки термограф (закрытый белой бумагой) и гигрограф стояли на большом снежнике с 26.07 по 11.08.1973 (рис. 6д2). В середине этого промежутка снежник стаял почти полностью под действием установившегося сильного лобового ветра, и приборы оказались на сыром илистом субстрате. На показаниях это изменение никак не сказалось. С падением влажности до 62% температура поднималась в начале второй половины дня до 15 . Эта температура соответствовала температурам: 18° на холме с куртинной дриадовой тундрой, 20° на уступе над долинкой ручья со злаково-кустарничково-моховой тундрой и 19° на галечнике ручья в глубокой долинке. Влияние снежников на жизненную среду ограничивается областью стока талых вод. Здесь устанавливается специфический нивальный режим среды, влекущий за собой формирование нивальной растительности, состоящей из хионофильных видов. Температура талых вод уже вблизи края снежника обычно 4°, а у самого его края 1°. Такие же величины температур имеют выходящие на поверхность ручейки от подтаивания вечной мерзлоты на склонах сопок. Эти выходы всегда связаны с ti до 10°. После стаивания снежника подсохшая поверхность его ложа может нагреваться до 30° даже в августе.
Для сопоставления температурных особенностей контрастных местообитаний построены рис. 6, а—д, на которых верхние кривые показывают суточные максимумы, нижние — суточные минимумы. Таким образом, между кривыми имеем суточные амплитуды для разных местообитаний. Сопоставляться между собой рисунки не могут, так как одновременность данных распространяется только на каждый рисунок в отдельности (см. даты).
Кроме суточных экстремумов на этих рисунках можно видеть,
что изменения температур на разных местообитаниях имеют один
и тот же знак, но разный сдвиг. При ухудшении погоды на одних
местообитаниях температура понижается относительно предшествующей более, чем на других. При резком похолодании с выпадением снега (рис 6, г, 22 июля) температуры близ поверхности
остаются все же положительными, хотя в глубоких долинках
ручьев они могут быть менее 1°. В Ванкаремской низменности такие погоды связаны с холодными морскими туманами, часто
сопровождаемыми ветрами. В результате этого сочетания образуется изморозь.
В еще большей мере различие температурных сдвигов на разных местообитаниях наблюдается при повышениях температур с наступлением солнечной штилевой погоды (рис. 6, в). При этом крупные различия температур необязательны для горного макроландшафта и весьма слабо связаны с характером макроклимата, т. е. отдельные участки деятельной поверхности могут нагреваться до температуры, казалось бы, несопоставимой с данным макроклиматом. Так, в Ванкаремской низменности, близ устья Рекууля приземный слой воздуха нагревается до 30° (рис. 6, г}, но и температура воздуха при этом 25°. Для Ванкаремской низменности это, конечно, очень высокая температура, даже если она и завышена на 2—3°. Различие температурных сдвигов на разных субстратах в одно и то же время обусловлено теплопроводностью и теплоемкостью субстрата, что подчинено физическим законам и имеет всеобщее значение. Р. Гайгер (Geiger, 1957) привел очень наглядные графики, на которых видно, сколь существенно различается теплопроводность гранитной породы, песка и болотистой дернины. Наиболее глубоко прогреваются скалы, наименее — торфяная дернина. Глубина прогревания связана и с интенсивностью нагрева поверхности. В одно и то же время сильнее нагреваются граниты, затем песок и наименее — торфяная дернина, В этой же последовательности наблюдается уменьшение амплитуды колебаний температур. Среднегодовая амплитуда температуры на гумусном, субстрате на 3° больше, чем на суглинке, и на 1,5° больше, чем на песке. На температуру близ поверхности почвы оказывает существенное влияние ближайшее окружение. Так, на рис. 6, б, температуры понижены потому, что выше места, где стоял самописец, на склоне находился большой снежник, с которого «скатывался» вниз холодный воздух. На этом же рисунке видно, что на каменной осыпи при солнечной безветренной погоде температура подскочила больше, чем на южном склончике в долину, хотя поверхность осыпи представляет неблагоприятное местообитание и растительность на нем крайне разрежена, а на южном склончике находится одна из замечательных луговин в районе. На рис. 6в, г также можно видеть максимум нагрева на горизонтальной поверхности и могильном бугорке на высокой террасе, с очень разреженной растительностью. Здесь суточная амплитуда 13 июля была 35°, а на берегу озера под склоном террасы 26°, тогда как на болоте около озера 27°. Однако, поверхность болота нагрелась сильнее, чем песчаный берег озера, на 3°, а ночью остыла меньше.
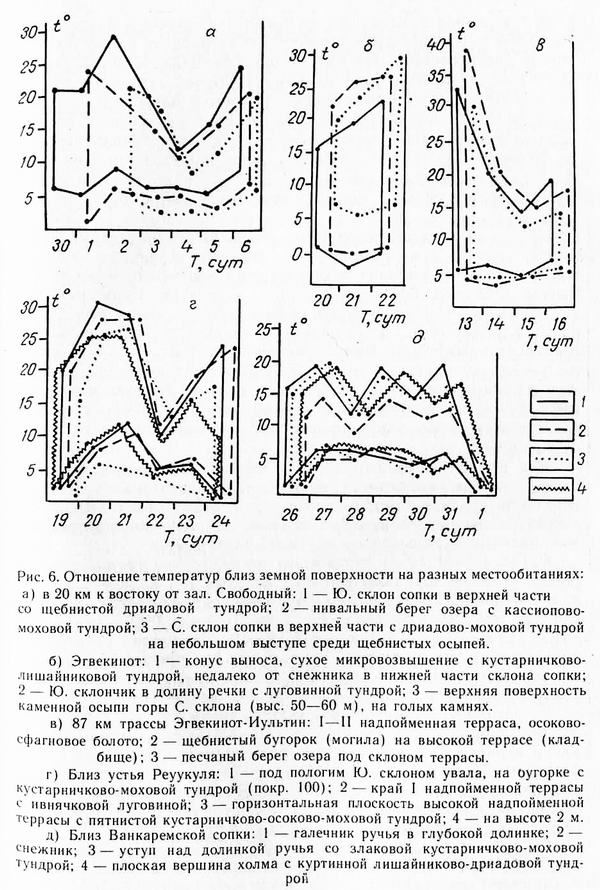 |
На основе всех температурных изменений, проведенных нами, можно сделать пока еще предварительный вывод, что в более континентальных районах температурная дифференциация по элементам рельефа более резка, чем в океанических районах. Эта дифференциация связана прежде всего с большими суточными максимумами, а последние связаны с более высокими среднесуточными. Близ Ванкаремской сопки (рис. 6, д) температуры весьма контрастных местообитаний имеют довольно близкие максимумы и минимумы, так как температура воздуха низка. На мысе Ванкарем в последней декаде августа 1973 г. суточный ход температур был одинаков на голой гальке и в осоковом болоте неподалеку. Как данные самописцев, так и многочисленные несистематические замеры температур позволяют сделать вывод, что поверхность сухих местообитаний с самой разной растительностью может нагреваться очень значительно, но действие нагрева может целиком скрадываться другими факторами.
Р. Гуд (Good, 1964) считает, что «в географии растений температура более фундаментальный фактор, чем количество осадков, а в экологии растений количество осадков более важный фактор, чем температура». Е. Одум (Odum, 1959) указывает, «что взаимодействие температуры и осадков, как и взаимодействие большинства факторов, зависит как от относительной, так и от абсолютной величины каждого фактора. Таким образом, температура оказывает более лимитирующее действие на организмы, когда условия увлажнения экстремальны, т. е. либо очень велики, либо очень малы, чем когда они умеренны, точно так же влага играет более критическую роль при экстремальных температурах». Такие отношения говорят в пользу применения гидротермических индексов при сопоставлении территорий по особенностям их растительного покрова; территорий любого размера - от физикогеографической страны до местообитания. Е. Одум (Odum, 1959) считает, что поскольку любой организм чувствителен к изменениям температуры, а последняя легко измеряется, то ее значение, как лимитирующего фактора, иногда переоценивается. Одум дает дружеский совет экологам: при исследованиях особенностей распределения организмов непременно изучать температурный режим их местообитаний, но не останавливаться на этом.
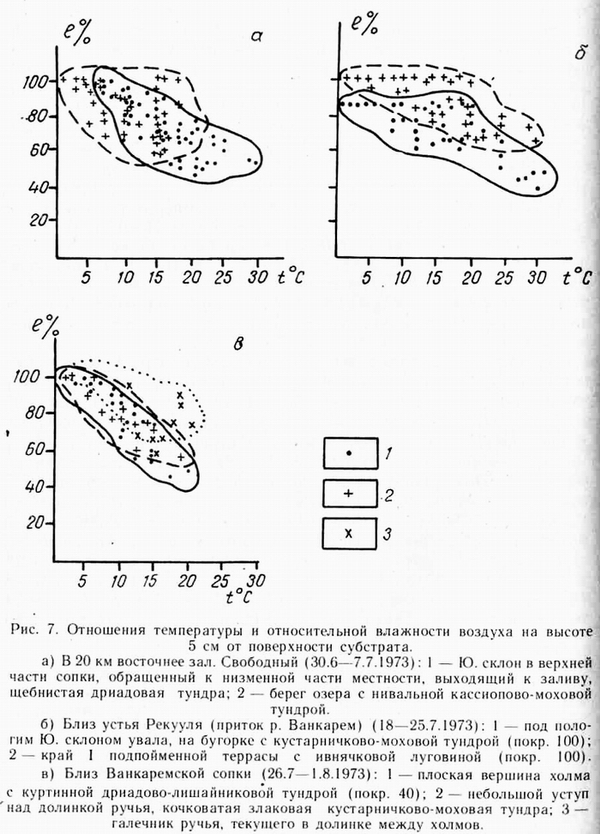
Следуя совету Одума, попытаемся выяснить связь между температурой и влажностью. Чтобы получить исходные данные, мы ставили рядом термограф и гигрограф на контрастные местообитания. Выяснилось, что связь температуры и влажности носит обратно пропорциональный статистический характер, т. е. одному значению каждого показателя может соответствовать несколько значений другого (рис. 7, а, б, в). На этих рисунках можно видеть, что при существенном налегании, области значений t ° — e % несходных местообитаний все же заметно различаются. Это различие заключается в том, что: 1} на одних местообитаниях не достигается температура других местообитаний, 2) на одних местообитаниях не случаются некоторые сочетания t °-— е%, характерные для других. Для сравниваемых таким образом местообитаний характерна несходная растительность. Как показывает |
сопоставление лент термографа и гигрографа, стоявших рядом, изменение температуры влечет за собой изменение влажности, а не наоборот. Некоторый сдвиг пиков вправо на кривой влажности по сравнению с кривой температур может быть связан с различной инертностью приборов, однако указанная связь температуры и влажности является выражением физических законов. Количество испаряющейся влаги зависит, в соответствии с законом Дальтона, от температуры испаряющей поверхности (Geiger, 1957).
Пользуясь приведенными отношениями температуры и влажности воздуха в околоземном слое, можно определить ночную уходящую радиацию с помощью графика Р. Гайгера (Geiger, 1957. С. 17). Например, если берем данные рис. 7б (эти данные берутся не с рисунка, а с лент самописцев, соответственно времени суток), то получим следующие значения отраженной радиации. Под пологим склоном увала со сплошным растительным покровом отраженная радиация колеблется от 3,8 до 6,3 кал/см кв. в мин. На краю надпойменной террасы с ивнячковой луговиной (этот же рисунок) отраженная радиация колеблется от 6,5 до 7,3 кал/см кв в мин. Таким образом, радиационное различие этих местообитаний весьма показательное. Оно объясняется свойствами самих местообитаний и их взаимоотношением с окружением. Под пологим склоном накапливается влажный воздух, который стекает со склона, в результате чего ночью влажность воздуха здесь 100%, С края террасы влажный возду стекает в пойму, поэтому максимальная влажность здесь ночью — 83% - несколько ниже, чем под склоном, но в холодную погоду они были несколько выше. В целом микроклиматические условия на краю террасы оказываются более благоприятными для растительности, поэтому здесь растительность — луговинная, тогда как под склоном — типичная тундровая.
График действителен лишь для ночной эффективной отраженной радиации, поскольку построен на использовании закона Стефана-Больцмана, который в строгом смысле применим только к абсолютно черному телу. Однако, для расчета тепловой радиации этот закон, по-видимому, применим и для дневного времени, так как для инфракрасной (тепловой) части спектра «большинство поверхностей являются абсолютно черными телами, т. е. они поглощают почти всю приходящую Радиацию» (Falckenberg, цит. по: Geiger, 1957). Известен парадокс Фалькенберга, дающийся инфракрасной радиации — «свежевыпавший снег является «наичернейшим телом», поскольку его альбедо составляет только 0,5% инфракрасной Радиации.
|
|
Из отношений, показанных на рис. 7а, по графику Р. Гайгера следует, что ночная отраженная радиация на щебнистом склоне сопки, в верхней части с дриадовой тундрой весьма постоянна (5,7 кал/см кв. в мин) несмотря на изменения влажности и температуры. На берегу озера с нивальной кассиопово-моховой тундрой в этот же период уходящая радиация колеблется от 5 до 6,3 кал/см кв. в мин. Среднее из этих колебаний соответствует уходящей радиации на щебнистом склоне сопки, т. е. в совершенно иных условиях и с другой растительностью. Гайгер считает, что отраженная ночная радиация в первую очередь зависит от относительной влажности воздуха, т. к. на его графике кривые влажности почти перпендикулярны оси радиации.
Связь температур и влажности носит обратнопропорциональный характер, хотя и статистический. На западе Чукотского полуострова, где нередка низовая облачность, относительная влажность 100% может держаться по несколько суток подряд. Температуры в это время, конечно, низкие. В ясную, а иногда и в пасмурную погоду, влажность воздуха снижается до разных величин, в зависимости от местопложения и его окружения. На рис. 7в заметно, что за один и тот же период влажность на вершине холма была постоянно ниже, чем на галечнике небольшого ручья (надо думать, что наличие ручья несущественно отражалось на уровне влажности), а на уступе над долинкой этого ручья уровень влажности в среднем был промежуточным между поверхностью холма и галечников ручья, так как сам уступ занимал промежуточное высотное положение. Анализируя и другие подобные случаи, нельзя не сделать вывод, что на положительных элементах рельефа суммарная относительная влажность ниже, а суммарная температура выше. Таков экологический вывод, верный до какого-то, пока неопределенного, предела. Этот вывод действителен для не очень крупных форм рельефа, ибо мы знаем, что в географическом плане отношения меняются. Верхние части гор Чукотки имеют большую суммарную влажность и более низкую температуру, чем их нижние части.
Как известно, величины температурных амплитуд и количество осадков являются показателями континентальности-океаничности макроклимата. Анализируя данные (рис. 6, 7) и привлекая при этом эколого-географические черты растительности приводимых местообитаний, следует сделать заключение, что комплекс местообитаний одного ландшафтного района также может анализироваться по принципу оценок континентальности-океаничности микроклиматов. Возможно, что показатель влажности при таком подходе, как наиболее легкий для измерения, может заменить показатель количества осадков, различия которых в одном пункте в летнее время трудно уловимы. |
Отмечалось, что распределение растений по земной поверхности подчинено одним и тем же закономерностям, независимо от размеров исследуемой территории (Boyko, 1947; Dansereau, 1957, 1966). Это означает, что в делении любой территории, начиная от поверхности Земного шара (Good, 1964) до территории площадью около 100 кв. км, по флористическим, геоботаническим и климатическим признакам всегда обнаруживается тесная связь, которая может быть сопоставлена как по величинам отдельных факторов среды, так и по отношениям этих факторов. Для исследования теплового режима различных местообитаний следует разрабатывать калориметрические способы, включая самопишущую портативную аппаратуру.
Необходимо также отметить, что частные, даже систематические данные по элементам физико-географической среды, по-видимому, будут заменены в будущем интегральными элементами, методика получения которых находится в стадии разработки (Бойчук, Марченко, 1968). Даже располагая методиками получения интегральных данных по отдельным элементам среды, нельзя оценить ее холоценотическое влияние на организмы, и, вероятно, потребуется разработка интеграции интегральных отдельных факторов среды.
В Онон-Аргунской степи В. Б. Сочава и др. (1970) использовали для сопряженного изучения экологических режимов метод комплексной ординации, заключающийся в «систематизации и количественной оценке главнейших связей и соотношений внутри геосистемы». Связи между факторами среды оценивались авторами с помощью корреляционного и регрессионно-дисперсионного анализов. Это очень сложный путь, и при экологических исследованиях в тундре требуется найти более простые, но адекватные природным изменениям пути решения проблемы холоценотического воздействия среды на тундровые организмы.
Экофон особи и экофон вида
На каждое растение действует комплекс внешних по отношению к нему факторов, представляющий величины по крайней мере трех порядков: общий климат (местный) и макроландшафт; мезоклимат и ландшафт урочища; ближайшие непосредственно действующие на растение факторы. Эти комплексы не противопоставляются друг другу, а входят друг в друга по степени уменьшения порядка по принципу русских матрешек. Поскольку обитаемый растениями слой в Арктике очень невелик, то огромное средообразующее значение имеет подстилающая (деятельная) поверхность, т. е, характер субстрата. Комплекс взаимодействующих факторов среды, их суммарный (интегральный) эффект, обусловленный в значительной мере свойствами субстрата, в дальнейшем будет называться в этой работе экофоном. Иными словами экофон — это приземный слой атмосферы и верхний слой педосферы, экологический режим которых определяется свойствами подстилающей поверхности. Согласно Б. А. Тихомирову (1971), это ниша «наивысшего напря жения жизненных процессов и максимального накопления органических веществ «растениями».
Наша концепция соответствует представлениям А. А. Крауклиса (1974), который отмечает,что «геосистему можно рассматривать как природный эффект, связанный с земной поверхностью». В 1969 г. А. А. Крауклис предложил вводить в характеристику ландшафтной структуры понятие о физико-географическом фоне, которому подчинены локальные географические системы фации (Сочава, 1974). В. Б. Сочава (1974) считает, что «для каждой фации нужно иметь в виду несколько соподчиненных фонов», соответственно с иерархическим подразделением географической среды. В своих построениях мы исходим из заведомого предположения, что суммарный эффект среды на растение (и вообще живые организмы) не является просто арифметической суммой отдельных факторов, а в той или иной степени отличен от нее.
Климат местности (местный климат) определяет характер не только флоры и растительности, но и характер эрозии, нивальность и другие абиотические процессы, являясь одним из основных моментов рельефообразования (Бондарчук, 1949). В первую очередь местный климат влияет на процесс образования поверхностных отложений, что связано также с топографией местности, соответственно которой местный климат дифференцируется на систему мезоклиматов урочищ и более дробных микроклиматов местообитаний.
Каждая ступень климата, являясь частью общего климата, имеет собственную суточную цикличность, которая определяет внутриландшафтное перемещение воздушных масс и которая, складываясь, определяет сезонную и годовую ритмику ландшафтов (Солнцев, 1960а). Микроклимат некоторых местообитаний совершенно не соответствует макроклимату района (Geiger, 1957).
Геоморфологические процессы приводят к быстрым изменениям влажности грунтов, глубины протаивания и, следовательно, почвенного покрова и растительности (Peterson, Billings, 1978). Авторы исследовали изменения растительности около песчаных обрывов и установили зависимость этих изменений не только от влажности, но и от преобладающих ветров, обусловливающих ту или иную развеваемость песков. Они отметили меньшее значение автогенной сукцессии, сопровождающейся повышением уровня мерзлоты и стабилизацией растительности. Вопрос об автогенных сукцессиях в Арктике должен быть поставлен со всей остротой, т. е. существуют ли они вообще в этом Регионе?!
Экофон в Арктике является преимущественно абиотической частью экосистемы, но тогда, когда поверхностный покров полностью или большей частью образован биогенной составляющей, например, на моховых коврах, суммарный эффект среды включает, конечно, и ее. В таких случаях биогенную составляющую невозможно отделить от абиогенной, не нарушив комплексность условий обитания. Принято называть микросреду обитания растительной особи биотопом, если она практически полностью определяется растительностью (Dansereau, 1957}, например, в условиях темнохвойно-таежных лесов с мощным наземным моховым покровом, где обычны Lycopodium complanatum, Goodiera repens, Trientalis europaea и др. Однако и в этом случае, по отношению к отдельным растениям и даже видам, можно говорить об экофоне, принимая условно растительность за фактор среды. Принимал же Майджер (Major, 1951) флору за фактор среды, определяющий растительность; обратное допущение не менее естественно.
Роль биогенной составляющей экофона Чукотки подчинена зональным условиям и вертикальной поясности. От практически нулевых значений, т. е. когда биогенная составляющая не оказывает никакого влияния на комплекс абиотических факторов, она постепенно увеличивает свою роль до образования неделимого комплекса с абиогенной составляющей. Это изменение прослеживается параллельно и взаимосвязано с усилением континентальности климата. Средообразующее влияние подстилающей поверхности также зависит от географического положения местности (Константинов и Сакали, 1967).
Теоретически экофон существует вокруг любой точки на подстилающей поверхности. Но поскольку наименьшей целостной единицей является растительная особь (система с весьма высоким уровнем целостности), то ее экофоном будет тот суммарный эффект среды, который испытывают все части растения; он может быть назван экофоном особи (ЭО), который представляет своего рода «экологический футляр» растения. Границы этого «футляра», возможно, и существуют, но пока мы не можем их определить. Существенно то, что для арктических растений этот «футляр» невелик. Это дает методические преимущества при сравнительном исследовании его свойств.
Н. Полунин (Polunin, 1960) справедливо считает, что «очень важно при экологических исследованиях иметь в виду концепцию «микроместообитания» (microhabitat ), которая является фундаментальной». Этой концепцией и является ЭО. Алгебраическая сумма экофонов особей одного вида будет представлять экофон вида (ЭВ). Это понятие распространяется на все пространство, занятое видом, и этим отличается от понятия широты местной экологической амплитуды. Кроме того, представление об экофоне вида, как и вообще об экофоне, связано в конечном счете с количественной оценкой среды, облекающей все особи данного вида, что пока имеет только теоретическое значение.Представление об ЭО и ЭВ связано с представлением о соподчиненности элементов одного и того же явления — экологической оболочки. Большое пространство делится на все более мелкие части, пока не получаем то пространство, которое заключает одно растение. Поэтому распространение понятия экофон на особь и на вид не должно казаться искусственным; фактически речь идет об одном и том же — функциональных особенностях среды, связанных с деятельной поверхностью.
Таким образом, экофон вида не может выходить за пределы толерантности этого вида к суммарному разнообразию среды на протяжении его ареала. Широта местной экологической амплитуды вида (или местный ЭВ) точно так же ограничена амплитудой толерантности вида в ландшафтных условиях небольшой территории. Толерантность является биологической характеристикой вида и обусловлена его генофондом, от которого зависит возможность существования данного вида в том или другом комплексе внешних условий. Р. Гуд (Good, 1964) использует понятие толерантность как в отношении действия отдельных факторов среды (температурная толерантность, толерантность к переувлажнению или, напротив, к засухе), так и в отношении их интегрального комплекса. Р. Гудом введено понятие потенциального ареала, т. е. пространства, на котором какой-либо вид может существовать по своей толерантности, но по различным причинам не существует. Понятие потенциального ареала является теоретическим, так как доказать действительную пригодность данного участка земной поверхности для заселения каким-либо видом, отсутствующим здесь, можно, лишь определив количественно среду или путем массовых искусственных посадок (фитомеров).
Наши наблюдения показы вают, что любой вид может произрастать в каком-либо районе лишь при наличии достаточно выраженного в ландшафте этого района потенциального экологического ареала, соответствующего толерантности вида. Этот ареал включает некоторую совокупность сходных по режиму среды местоположений со сходной по экологическим требованиям (но не обязательно по систематическому составу) растительностью; единичные или малые по площади местоположения (с комплексом однотипных условий) часто не способны сохрнить популяцию какого-либо вида с узкой экологической амплитудой в данном районе (Кожевников, 1973 6). Такие местоположения с большим трудом заселяются и иммигрантами со смежных территорий в том случае, если только на этих е диничных местоположениях условия среды отвечают толерантности иммигрантов. Это суждение, безусловно, нуждающееся в проверке путем количественнных измерений среды, настраивает на ту мысль, что поотенциальный ареал имеет некий пространственный предел, ниже которого участки земной поверхности, даже если их условия и находятся в пределах толерантности некоторых видов растений соседних территорий, не относятся уже к потенциальному ареалу, этих видов. Такая ситуация возникает, например, в случаях «островного» отсутствия вида в пределах его сплошного в общемто ареала. «Островные» дизъюнкции (дырки) часто обусловлены, особенностями ландшафтов на их протяжении. Однако подобные «дырки» нередко окружены областью распространения вида, отсутствующего в данном месте. Таким образом, физикогеографические условия вокруг «дырки» не препятствуют произрастанию вида, а в пределах «дырки» имеются местообитания (пусть их даже очень мало), на которых отсутствующий вид мог бы произрастать. Отсюда высказанное суждение представляется справедливым. Приведенные представления в корне противоречат высказываниям А. И. Толмачева (1932—1935): «наличие и степень развитости определенных стаций, имея решающее значение в определении степени обычности отдельных видов, оказывается совершенно второстепенным фактором в определении; собственно состава флоры» ... «Изменение характера субстрата оказывается само по себе недостаточным для определения каких либо абсолютных черт состава флоры».
|

Чукотский полуостров, Лоринские горячие источники.
Фото А.А. Галанина. |
Экофон растительности особи и экофон вида являются проявлением толерантности вида к определенным условиям среды толерантность же — это наследственно обусловленная характеристика вида, являющаяся выражением специфики обмена вещества и энергией. X . Мезон (Mason, 1946 a , б; Mason, Stont, 1954 и др.) показал, что толерантность особи меньше толерантности вида, и это имеет как географическое, так и экологическое значение. Экофон как среда отдельной особи и, далее, как сумма ЭО, т. е. ЭВ могут быть названы экофитноменом у среды, поскольку среда оценивается в связи с пристутсвием растения. Однако оценка среды, качественная или количественная, может быть произведена и без присутствия определенного растения, но с отношением к его возможному присутствию здесь, т. е. с учетом его потенциального экологического ареала. Сраведливость этого суждения может быть понята из очевидной возможности произрастания представителей какого-либо вида в любой точке конкретного достаточно однородного местообитания, где этот вид имеется. Существуют местообитания (например, очень ровные нагорныа террасы) с практичеси одинаковыми условиями среды в любом его части, обширные по площади, с сильно разреженной растительностью (фитоценотические связи не существуют), слагаемом некоторой совокупностью видов, индивиды которых могут произрастать в пределах этого местообитания в любой точке, но произрастают в тех точках, куда волею случая попали их диаспоры.
Конкретное место произрастания растения в пределах однородного по условиям обитания топографического контура большей частью случайно, тогда как произрастание его именно в условиях этого контура (или другого, аналогичного) определяется биологией вида, и случайность здесь уже не имеет превалирующей роли. Если диаспоры какого-либо растения, толерантного к условиям данного местообитания, |
попадают на него, то они развиваются в нормальные особи, чего не происходит в условиях, к которым данный вид не толерантен. Следовательно, имеется основание говорить об экофоне топоэлемента, в данном случае, экофоне местообитания (ЭМ). Л. Г. Раменским в 1938 г. была выдвинута концепция экотопологии (Раменский, 1971), которую мы в данном случае интерпретируем в соответствии с попыткой представлять среду как единое целое, распадающееся на несколько субординационных порядков.
Типологический экофон представляет орографическую поверхность, ограниченную контуром элемента мезорельефа или микрорельефа с однородным мезо или микроклиматом. Бессмысленно говорить о климате слишком дробных подразделений ландшафта, исходя из определения климата как многолетнего устойчивого режима погоды (Хромов, 1952). Из этого определения климата может быть выведено заключение об объеме местообитания, а именно, что оно не может быть слишком мелкой единицей поверхности, поскольку не существует климата кочки (Хромов, цит. соч.). Относительная устойчивость мезо- и мик роклимата местообитаний является их основным гомеостатическим фактором в течение отрезков времени, превышающих смену, по крайней мере, нескольких поколений одного вида и групп видов, особенно, узкоспециализированных по отношению к местообитаниям в данном районе. Есть основания полагать, что устойчивость мезоклимата подурочища (включающего обычно несколько различных местообитаний) выше, чем устойчивость макро- и микроклимата, характерного для ландшафта как типа местности.
Следует учитывать, конечно, что экофон данной точки подстилающей поверхности есть динамическая совокупность факторов, протекающая во времени с определенной периодичностью (суточные и сезонные изменения, многолетние и вековые циклы) и некоторой амплитудой отклонений (изменения погоды) от средних значений в каждый момент цикла или периода. Особи растений, как и все живые организмы, существуют с определенной наследственной нормой реакции на изменения среды обитания, но видам и популяциям, к которым они относятся, свойственна, кроме того, генетическая изменчивость этой нормы, что отличает надорганизменный уровень организации от организменного.
Временная изменчивость среды, которая неизмененно сопровождала существование живого с самого начала его появления, подразумевает и изменчивость пространственную, поскольку четырехмерное пространство является минимальным относительно любой точки выбранной за начало координат. Поскольку растения испытывают временные изменения среды обитания, они существуют и на фоне пространственных ее вариаций. Можно сказать, что сумма прошлых временных изменений нашла отражение в пространственной изменчивости среды в каждый настоящий момент, как бы спроецировалась в настоящее, «разливаясь» в пространстве. Результатом больших временных интервалов является органическая эволюция, но нас сейчас занимают сравнительно меньшие промежутки времени, сопоставимые со «средним» временем существования современных видов. Представив себе историю развития вида в пространстве и времени до наших дней, мы снова придем через адаптивную норму реакции или селекцию неприспособительных модификаций (Шмальгаузен, 1968) к идее толерантности. Толерантность же связана с понятием экофона вида, по отношению к которому она проявляется, т. е. имеется в виду экофитномен (присутствие растения в такойто среде).
Всякий однородный по условиям среды участок поверхности обнаруживает в силу космических и иных причин постоянную временную изменчивость, а следовательно, и пространственную, заключающуюся в том, что разные точки этого участка могут различаться между собой по интегральной совокупности факторов в каждый момент времени. Однако их режим остается одним и тем же (общим), но лишь «сдвинутым по фазе». Постоянство среды на однородном участке есть постоянство комплекса изменений, но подчиненное гомеостатической целостности, т. е. комплекс изменений инвариантен. Каждое конкретное местообитание обладает собственной пространственно-временной инвариантностью.
На языке ландшафтоведов местообитание представляет физико-географическую фацию — часть урочища, характеризующуюся однородным микроклиматом, обшей литогенной основой (Солнцев, 1949). Согласно Солнцеву (цит. соч.) и Арманду (1952), физико-географическая фация имеет площадь от 1 га до 1 км 2 ; при этом признаком фации является то, что на ней в девственных районах развивается один фитоценоз. С точки зрения геоботаника решительно невозможно представить себе фитоценоз, имеющий подобную площадь даже на равнине в лесной зоне. В тундрах — это и вовсе невероятное событие даже для безупречно девственных районов. Отсюда формулировка В. Б. Сочавы (1959), что «биогеоценоз — это физико-географическая фация, но понимаемая как энергетическая система»,— является лишь частным и редким случаем. Тем не менее, например, на о-ве Лохид на севере Канады отмечено, что для каждого типа породы, содержащего определенное количество влаги, имеется свой тип растительного сообщества (Edlund, 1980). В Арктике одна физико-географическая фация несет, как правило, несколько фитоценозов в понимании геоботаника. Однако это не специфическая черта растительности Арктики. То же самое характеризует растительность лесной и степной зон. В. Н. Сукачев еще в 1949 г. писал, что фация значительно шире, чем фитоценоз; фация включает, как правило, несколько фитоценозов. Но и после этого ландшафтоведы снова не раз утверждали (и в том числе Крауклис, 1974), что площадь фитоценоза соответствует площади фации. В ряде работ эта установка опровергается на фактическом материале. Так, в статье М. Я. Лайвиньша (1967), посвященной картографированию внутрифациальных различий, можно видеть, что к одной и той же фации относятся различные фитоценозы, рассматриваемые автором как парцеллы в пределах фации.
Из сопоставления понятий «физико-географическая фация» и «местообитание» можно вывести заключение, что целесообразно в случаях сложных поверхностей, например, бугристо-мочажинных болот, с голыми пятнами суглинка рассматривать их как одно местообитание. В его пределах могут быть выделены более дробные категории (в полевых этикетках обычно называемые конкретным элементом микрорельефа) подобно тому как в фации выделяются подфации и участки. С биогеоценотической точки зрения такое болото вполне отвечает одному биогеоценозу. Согласно авторскому определению «биогеоценоз представляет собой всякий участок земной поверхности, где на известном протяжении биоценоз и отвечающие ему части атмосферы, литосферы, гидросферы и педосферы остаются одинаковыми, имеющими однородный характер взаимодействия между ними, и поэтому в совокупности образующие единый, внутренне взаимообусловленный комплекс» (Сукачев, 1949). По площади биогеоценоз совпадает с фитоценозом. Биогеоценоз, таким образом, представляет элементарную экосистему в смысле А. Тэнсли (Tansley, I 935).
А. Тэнсли развивал концепцию экосистем отчасти потому, что рассматривал биотическое сообщество как «неестественную концепцию» (Sjors, 1955). Очевидно, понятие «биогеоценоз» основано на этом же представлении. Современная экология ландшафта пополнилась недавно чрезвычайными важными представлениями, в частности следующими» «...взаимообусловленность между компонентами природной среды, рассматриваемая на уровне географических фаций, имеет не абсолютный характер, а прослеживается лишь как более или менее выраженная тенденция. Соответственно и площадь, занятая одной и той же фацией, обнаруживает большую или меньшую неоднородность по признакам главнейших компонентов. Отклонение от однородности — не исключение, а норма, что объясняется динамическими факторами» (Крауклис, 1974).
Экофон ландшафта
С целью ботанико-географических сопоставлений многих районов, кроме местообитаний, в качестве более крупных экотопологических единиц можно использовать комплексы местообитаний, сходные в известной мере по условиям среды. Комплексы включают разные серии местообитаний, достаточно близкие по экологическому режиму. Такие комплексы называются в этой работе ландшафтным экофоном или ландофоном (ЛЭ). Необходимость выделения этих единиц вызвана преимущественно гористым характером изученной территории. Основной особенностью ЛЭ, на которые подразделяются ландшафты, является та, что это по-видимому, минимальные хорологические единицы, на основе которых может осуществляться связный историко-флористический и эколого-флористический анализы.
Мы полностью разделяем мнение Н. В. Тимофеева-Ресовского (1961), что «основной низшей систематической единицей в географии (элементарный ландшафт или фация по Л. С. Бергу, 1945}, не обязательно должен быть элементарный биогеоценоз, может быть элементарным ландшафтом следует считать физико-географически (ландшафтоведчески или геоморфологически) объединенную группу территориально смежных элементарных биогеоценозов». Такая единица была впоследствии названа элементарной геохорой (Сочава и др., 1970). Мы исходим из тех же соображений, что и упомянутые авторы, а именно при сравнительно-географическом анализе более рационально иметь дело с заведомо комплексными единицами, чем с предположительно элементарными. Объем комплекса определяется сообразно целям и возможностям исследования.
Местообитания объединяются в один ЛЭ по их режиму среды. Группировка местообитаний в один ЛЭ теоретически должна производиться на основании близости их интегральных индексов среды. Но поскольку таких комплексных оценок среды пока не существует, то следует выбрать какой-либо фундаментальный признак, дающий возможность приблизительно оценить среду. Этот признак должен иметь крупное нивелирующее значение (приведение к одному уровню), так как служит для группировки подчас мало сходных по флористическому составу и по растительности местопложений. Нами была предпринята попытка использовать в качестве такого фундаментального признака современный тип поверхностных отложений, реже близость тех природных образований, которые комплексно определяют режим среды (снежники, озера, море), или тип биогенного субстрата (торф, богатство почвы гумусом). Характер субстрата может, считаться наиболее естественным фундаментальным (для крупных объединений) признаком, так как поверхность определяет многие черты местообитаний, в том числе тепловой баланс (Сапожникова, 1950; Солнцев, 19606; Хромов, 1968; Русин, Флит, 1971; Константинов, Сакали, 1967 и др.), с которым связан радиационный баланс. Согласно М. И. Будыко (1971) «радиационный баланс довольно существенно различается для разных типов подстилающей поверхности в одном и том же районе. Большая «микроклиматическая» изменчивость радиационного баланса объясняется значительным влиянием на его величину альбедо и температуры подстилающей поверхности».
Каждый тип поверхностных отложений по-своему влияет на режим околоземной среды, где сосредоточены растения, и, таким образом, определяет их состав и численность. Температура и количество осадков в раздельном рассмотрении не связаны с распределением растений, поскольку одновременно действуют многочисленные локальные факторы ( Abbe , 1936). Между собой условия увлажнения и температуры также не связаны какой- либо зависимостью, т. е. при одних значениях температуры условия увлажнения могут быть разными и наоборот (Good, 1964). По отношению к температуре и влажности освещенность и ветер являются вторичными факторами, как указал Р. Гуд, потому что они отражаются на значениях первичных факторов. При изменении каждого из многочисленных факторов происходит перестройка всех других таким образом, чтобы общее изменение среды было наименьшим, т. е. факторы ее связаны компенсаторно (Rubel, 1935; Dansereau, 1957; Odum, 1959; Good, 1964; и др.). Однако постепенные изменения происходят на каждом типе отложений, эта постепенность отражена многоликими группировками растений, которые постоянно перестраиваются и по структуре, и по видовому составу, как это видно при посещении одних и тех же участков в течение нескольких сезонов.
|

Кустарниковая (Salix alaxensis) ивняковая и кустарничковая ивнячковая (Salix schamissonis) растительность сформировались на одном ЛЭ. Фото А.В. Галанина.
|
Согласно Дю Рие (Du Rietz, 1929), следующие факторы влияют на заселение видами любого местообитания в определенный период: 1) среда местообитания в принятое за начальное время, 2) распространенность видов в это время поблизости, 3) количество диаспор, 4) конкурентоспособность видов, 5) влияние животных, человека и паразитов растений, 6) интервал времени. Число сочетаний по степени значимости перечисленных факторов чрезвычайно велико и оно в значительной степени объясняет пестроту растительного покрова. «Если допустить многофакторную обусловленность растительности,— писал Л. Г. Раменский еще в 1925 г,— то наличность безграничного разнообразия растительных группировок станет вполне объяснимой». Но однофакторная ее обусловленность, очевидно, во всех случаях является только результатом упрощенного мышления ботаников (Egler, 1961). Пока не существует способов определения интегрального эффекта факторов среды и, по-видимому, наибольшее приближение к такому определению дает расчет радиационного баланса.
Поверхностные отложения являются только внешним, одним из коренных признаков ЛЭ; но своеобразие последнего определяется всей совокупностью факторов среды. Отсюда следует, что визуально ЛЭ могут сопоставляться по их растительности и наборам видов лишь в пределах территории более или менее однород ной по макроклимату, т. е. порядка физико-географической подпровинции или округа. Сказанное надо понимать так, что в разных подпровинциях значения ЛЭ, связанные с одним и тем же фундаментальным признаком, будут существенно различны, что отразится и в растительном покрове. В предпринятом Н. А. Солнцевым (19606) анализе |
взаимоотношений живой и неживой природы были сформулированы три принципа: «во взаимоотношениях живой и неживой природы имеет место принцип неравнозначности взаимодействующих факторов; все факторы разбиваются на три группы: литогенные, гидроклиматические, биогенные; ведущие факторы — литогенные. О последнем свидетельствует весь ход эволюции организмов, в котором существенный момент — способность организмов приспосабливаться к изменениям среды».
Литогенные факторы являются признаком крупного разряда при анализе экологии ландшафтов. Характер их действия может быть представлен как результат взаимодействия геологического строения местности и ее гидроклиматических факторов при определяющей роли рельефа. Литогенньгй фон является материальной основой для почвообразовательных процессов. Но последние еще теснее связаны с климатом. В разных климатических условиях происходят и разные почвообразовательные процессы, однако их влияние на растительность может быть одним и тем же (конвергентным). Слабые почвообразовательные процессы являются причиной отсутствия древесной растительности в некоторых районах среднего Урала (Сочава, 1945) и на Гудзоновом заливе (Маrr, 1948). В подобных случаях литогенные факторы, очевидно, не являются ведущими во взаимодействии «живой» и «неживой» природы. Они не являются ведущими и в зоне вечной мерзлоты, на тех местах, где развиваются болота, поскольку образование последних связано с условиями климата и рельефа. Но в большом числе случаев литогенные факторы действительно оказываются ведущими, и в Арктике это особенно хорошо прослеживается.
Поверхностные отложения в определенной мере представляют результат влияния современных литогенных факторов, поэтому говоря о них мы называем аллювий, делювий и пр., но не флювиогляциал, древние морские отложения и пр., имеющие солидный геологический возраст, изменившиеся вследствие выветривания, выщелачивания и т. д. Современные поверхностные отложения являются результатом процессов разрушения и перемещения горных пород, которым свойственна суточная цикличность в соответствии с таковой климата. Вместе с тем, поверхностные отложения оказывают действие на суточные амплитуды температур (Хромов, 1968), что является очень важным экологическим показателем.
Выделение ЛЭ соответствует принципам типологии земель с целью хозяйственного использования (Раменский, 1971; Odum, 1959; Polunin, 1960), основанном на оценке экологического режима территорий. Возможно, что ЛЭ соответствует понятию «геом», введенному В. Б. Сочавой (1970, 1974 и др.}, поскольку геом также является категорией промежуточной между топологическим и региональным уровнями исследования или, точнее говоря, высшей категории топологического подразделения. Однако соотношение этих понятий не совсем ясно в силу того, что понятие «геом», по-видимому, недостаточно разработано. Согласно В. Б. Сочаве (1974. С. 67), растительный компонент геома представляет формацию. Но в пределах одного ЛЭ может быть несколько типов растительности.
Соотнесение понятий в данном случае момент очень ответственный, поскольку понятия вырабатываются применительно к в разным природным зонам. Уже сейчас видно, что, если по одному условию понятия соответствуют друг другу, то по другим условиям не соответствуют. Очевидно, немалую роль в этом несоответствии играет различное соотношение биогенной и абиогенной составляющих среды в южной Сибири и на Чукотке. ЛЭ является высшей категорией Геосистем топологического порядка, граничащей с категориями геосистем регионального порядка. В отличие от понятия «геом», которое разработано еще недостаточно, мы более подчеркиваем гетерогенность ЛЭ, чем гомогенность, которая считается свойством геома. По всей вероятности, «гетерогенность ЛЭ» и «гомогенность геома» однокачественны, поскольку геом является весьма крупной площадной и структурной единицей. Гетерогенность ЛЭ и является основной причиной его инвариантности.
Геом соответствует, по В. Б. Сочаве (1974), макрогеохоре или округу, в пределах которого он существует. Подобным образом и ЛЭ распространен на соответствующих поверхностных отложениях в пределах физико-географического округа, а уже в соседнем округе на тех же отложениях существует иной, но аналогичный ЛЭ. Это представление должно способствовать определению природного выдела ранга округа: относительное постоянство структур каждого ЛЭ от одного пункта к другому означает нахождение в пределах одного округа. Соответственно, перестройка структур ЛЭ одного типа и растительности в нем означает переход в иной округ. Такой подход отвечает принципу к омплементарности абиотической и биотической составляющих среды и устраняет произвольное трактование ранга округа. Заметим, что с этой точки зрения материковую (западную) и океаническую (восточную) Чукотку следует рассматривать в ранге округов, а не подпровинций. С этим согласуется и представление об Арктике как о провинции Голарктиса (Тахтаджян, 1978) или подобласти (Кожевников, 1978а).
С учетом всех вариаций каждого ЛЭ и их взаимных переходов в каждом районе и округе совокупность ЛЭ в модельном отношении можно представить в виде некоей синусоиды, отображающей континуальные переходы между разными ЛЭ. Такая же синусоида характеризует смежный район и отношения аналогичных ЛЭ между районами. Модель подразумевает сходное развитие разных ЛЭ, чего в действительности никогда не бывает. Поэтому более реалистическая модель, включающая точное знание площадей ЛЭ и их структур, очевидно, не будет периодической. Выяснение этих отношений представляет увлекательную задачу. Кроме того, стык различных ЛЭ одного типа на границе округов может не строго соответствовать этой границе, т. е. «концы» одних ЛЭ не соответствуют «концам» других, что создает переходную полосу. Пространственные взаимоотношения ЛЭ и растительности в переходных полосах позволяют определить современную динамику растительного покрова в соответствии с направленностью изменений среды.
Растительность, характеризуемая по собственным признакам для целей фитоценологической классификации без привлечения факторов, обусловливающих ее, является слабым основанием для динамической интерпретации растительного покрова. Это в особенности касается растительности Чукотки, в которой в большинстве случаев отсутствуют ценотические связи, распределение и численность видов преимущественно не зависят друг от друга, а определяются абиотическими факторами. Согласно правилу экологической индивидуальности видов Л. Г Раменского (1925), нет таких двух растений, которые бы распределялись по достаточно большой выборке описаний идентичным образом. Но, как заметил Раменский, правило это не противоречит факту существования связанных ценотическими отношениями закономерных группировок. Оно свидетельствует лишь о действительно наблюдаемом в природе безграничном разнообразии количественных сочетаний растений. На Чукотке это правило в высшей степени действенно. На ее территории, как и во всей Арктике, растительность очень тесно связана со средой, абиотическая составляющая которой в большинстве случаев превосходит биотическую. Было показано, что даже условия нанорельефа оказывают на растительность заметное влияние (Александрова, 1960). Однако и на неарктических территориях установлено, что фитотолологический подход к изучению растительности имеет определенные преимущества перед фитоценологическим.
|

Комплексная растительность из сообществ с доминированием кедрового стланика (Pinus pumila) и сообществ кустарничков (Betula exilis + Vaccinium uliginosum + V. vitis-idaea + Empetrum nigrum) на одном ЛЭ. Фото А.В. Галанина.
|
При классификации растительности выделяют ее гомогенное и гетерогенное горизонтальное строение. В последнем случае за единицу классификации принимают тип комплекса (Кустова, 1979; Холод, 1985). При этом гомогенный фитоценоз расценивается как система с наиболее сильными внутренними связями, а макропоясный экологический ряд — с наиболее слабыми (Холод, цит. соч.). Авторы, однако, не заостряют внимание на том, что, собственно, гомогенность и гетерогенность растительности обусловлена прежде всего внешними причинами, чаще геоморфологическими. То же самое касается и комплексности растительного покрова. При статистической классификации растительности неучет причин ее гомо- и гетерогенности приводит к нелогичному использованию критериев классификации. Так, Н. В. Кустова (цит. соч.) при классификации гомогенного растительного покрова использовала общепринятые фитоценотические единицы - ассоциации, группы ассоциаций, классы ассоциаций и т. д. При этом относительно гомогенные и мозаичные сообщества в силу их полидоминантности и мозаичности объединялись в ассоциации «по характерному ядру сопряженных видов (Александрова, 1977), а для мозаичных сообществ по однотипности мозаичной структуры. Для гетерогенного растительного покрова основной единицей классификации принят тип комплекса в соответствии с классификацией комплексов, разработанной 3. В. Карамышевой и Е. И. Рачковской (1973). Комплексные сообщества систематизируются по форме, количеству и составу компонентов фрагментов слагающих их ценозов (гривчато-ложбинный комплекс ерниково-кустарничково-лишайниковых и травяно-моховых сообществ ). |
Ставшие традиционными принципы описания растительности без выяснения причин, ее обусловливающих, легли в основу огромного количества геоботанических работ, имеющих очень ограниченную полезность. Противопоставляя генетический подход к классификации растительности феноменологическому, В. Б. Куваев пишет: «пойменные, аласные, суходольные и т. п. луга вычленяются из пойменного, аласного и т. п. комплексов и объединяются в одной рубрике «луга». Но ландшафтный характер элементарных единиц требует иного подхода к их объединению. Принимая во внимание не только растительность, как «ботаническое лицо» типа, но и его местоположение в рельефе, материнскую породу, почву, ее мерзлотный, гидрологический режимы — словом, все то, что обусловливает облик этого «лица» — мы убедимся, что пойменные луга генетически гораздо ближе к находящимся в той же пойме степям, а не суходольным и т. п. лугам» (Куваев, 1968. С. 5). Характеризуя растительность, он выделял эколого-генетические ряды, объединяемые по ландшафтным признакам в 10 типов. Л. Г. Раменский (1925) отметил, что все списки растений по местообитаниям могут быть сгруппированы в плавные и разнообразные экологические ряды, связывающие их в единую сложную систему. Система эта будет иметь ландшафтную обусловленность со скрытыми причинными связями из-за того, что мы подходим к ней «не с того конца».
Экологи с ботаническим уклоном предпочитают основываться более на флористике, т. е. на наборах видов (Goldsmith, 1976). Очевидно, для экологического анализа необходимо изучение самой среды (ойкос). С этой точки зрения Л. Г. Раменский (1925) не был прав, полагая, что, только суммируя эпизоды взаимной борьбы растений путем массового статистического учета значительной площади с выводом итоговых средних обилий, мы обнаруживаем экологически закономерный состав растительности, отражающий условия обитания. Следует различать индикационную роль местного растительного покрова, отдельных сообществ и группировок, отдельных видов. Несомненная, «безошибочная» роль индикатора на Чукотке принадлежит только растительному покрову (включая местную флору в целом). Нам уже приходилось отмечать (Кожевников, 19736, 19746, 19756, 1979а}, что индикационная роль сообществ и группировок не всегда очевидна, несмотря на популярность противоположного взгляда. Это особенно касается группировок растений со слабыми или отсутствующими ценотическими отношениями. В еще большей мере это присуще отдельным видам. Такое положение с необходимостью ставит задачу исследования действительной индикационной роли растительных сообществ и группировок, а также отдельных видов, для чего требуется углубленное изучение среды обитания. Должен решаться вопрос: что, собственно, индицируется и с какой степенью достоверности?
Ландшафтная обусловленность растительного покрова убеждает в необходимости исторического анализа самих ландшафтов с тем, чтобы более явственно проступила история растительного покрова. Мозаичный характер современного растительного покрова Чукотки получит более убедительную историческую трактовку на фоне анализа динамики среды обитания в зависимости от рельефа, удаленности береговой линии, похолоданий и потеплений и т. д. Для рационального использования принципа актуализма также возникла необходимость выбора такой современной ландшафтноэкологической единицы, которая давала бы возможность более обоснованно представлять эволюцию среды, основываясь в значительной мере на современных экологических связях в привязке их к установленным палеогеографическим фактам. Полевые исследования показали, что в качестве такой единицы целесообразно использовать ЛЭ. Историческая сущность этого образования заключается в том, что каждый ЛЭ представляет особый канал миграции. По совокупности ЛЭ одного района мигрируют на новые территории виды с различной экологией и биологией. Если на новой территории имеются благоприятные макроклиматические условия, то из «исходного» ЛЭ виды расселяются по другим ЛЭ, одновременно увеличивая численность в «исходном» ЛЭ. Конечно, это происходит лишь в том случае, если генотипическая структура вида способна формировать новые биотипы (Turesson, 1925). Однако надо полагать, что все мигрирующие виды обладают этой способностью, иначе они не мигрировали бы. Новые биотипы формируются в популяциях с высокой модификационной изменчивостью, поскольку это увеличивает возможности отбора (Sorensen, 1954).
При ухудшении климата происходит обратный процесс. Прежде всего сокращается экологическая амплитуда вида, пока он не остается только в «исходном» ЛЭ, т. е. по которому он мигрировал в данный район. При анализе современных условий обитания ЛЭ уже не может быть назван элементарным звеном, роль последнего выполняет тип (или серия) местообитаний. Но и при современном ботанико-географическом анализе роль понятия ЛЭ имеет существенное значение, особенно в переходных климатических полосах Арктики, где, благодаря существованию континентально-океанической мозаики мезо- и микроклиматов на небольшой территории, уживаются, можно сказать бок о бок, виды и континентальные, и океанические по своей природе.
Каждый ЛЭ занимает достаточно протяженную, но не цельную площадь; например, торфяный ЛЭ в континентальных районах охватывает большую часть шлейфов сопок, нижние части самих сопок, поверхности шлейфо-террас и бессточные равнинные днища межгорных котловин. Кроме того, каждый ЛЭ заходит в пределы общего контура смежных ему ЛЭ языками различной формы и размеров и кроме того, представлен обособленными фрагментами в окружении других ЛЭ. Каждый ЛЭ обычно нерезко переходит в смежный; как правило, имеется широкая или узкая полоса экотона.
В резко континентальных районах изученной территории тип поверхностных отложений, как признак ЛЭ, более удобен, чем в океанических районах. Это связано с резкой контрастностью условий на различных элементах рельефа и, соответственно, с резкими различиями в растительном покрове и крупными площадями однотипных конкретных местообитаний. В районах с относительно океаническим типом климата возрастает степень мозаичности растительного покрова в пределах одного ЛЭ, однако это не может служить препятствием для использования понятия, охватывающего крупные топографические контуры и учитывающего только экологические характеристики растительности, в основном же, ориентирующегося на абиогенную составляющую среды. Л. Г. Раменским (1971) было указано, что «естественная мозаичность покрова приводит к необходимости учета значительных его площадей». Разумеется, что элементы мозаики и степень мозаичности увеличиваются соответственно увеличению размеров выбираемого топографического контура сообразно цели изучения.
Достаточно сказать, что на территории 100—200 кв. км выделяется 5—6 ЛЭ, чтобы представить, что это крупные объединения участков поверхностей со сходным режимом среды, имеющие гетерогенный состав растительности и флористических групп, но что еще более существенно, гетерогенность среды обитания. Но степень гетерогенности в одном ЛЭ является меньшей в сравнении с гетерогенностью между разными ЛЭ. В ряде случаев воздействие поверхностных отложений на растительный покров превосходит влияние микроклимата, который сформировался на этих же отложениях. В таких случаях главенствует эдафическая компонента среды обитания, о чем было сказано и выше. Например, грядовые микроповышения на равнинах с поверхностным слоем элювия существенно не отличаются по растительности от элювия плоских вершин невысоких сопок или внешних частей нагорных террас, хотя микроклимат этих местообитаний весьма различен, в частности по температурному режиму, по характеру увлажнения. Эти различия следует расценивать как различия в пределах одного ЛЭ.
В пределах территории одной местной флоры ЛЭ включает несколько близких типов местообитаний, каждый из которых включает группу сходных по режиму конкретных местообитаний. Понятно, что уровень сходства конкретных местообитаний и типов местообитаний будет неодинаковым. Местообитания, являющиеся частями одного урочища, будут, как правило, относиться к разным ЛЭ, хотя мезоклимат урочища будет для них общим. Ландшафтный экофон характеризуется действием одного или двух определяющих (или ведущих) комплексных факторов среды, которым подчинен суммарный эффект среды. Элювиальный экофон, например, характеризуется действием сильных ветров, определяющих микроклиматические и эдафические (выдув мелкозема) условия. Торфяный экофон — это большая или меньшая заболоченность и т. д. Несмотря на различные ведущие факторы, интегральный климатический показатель может быть выражен через температуру и количество осадков, которое включает в себя и зимнее перераспределение снега.
Существенным недостатком использования поверхностных отложений, как индикатора ЛЭ, является то, что здесь на микроклимат нередко оказывает сильное влияние положение участка в ландшафте (влияние окружения). Участок с горизонтальной щебнистой поверхностью на равнине будет отличаться по условиям среды от такого же участка, закрытого, хотя бы частично, окружающими его возвышениями. Поэтому и требуется разработка методов определения интегрального режима среды. Один ЛЭ может включать несколько растительных типов. Обратимся к следующему примеру. Ольховники в истоках Канчалана являются краевыми подзональными массивами. В нижней части склонов гор ольховники обычно моховые, т. е. это один ЛЭ для мелких растений. В верхней части ольховники не имеют моховой подушки, а растут на голом щебне, флористический набор, соответственно, совершенно иной по сравнению с нижней частью того же ольховника. По соседству с нижней частью ольховника располагаются моховые тундры, среди которых имеются ивняки, также моховые. Однако по всей нижней части такого склона флористический набор разных мест определяется только моховым ковром и почти не связан с кустарниками ивы или ольхи. Единственное исключение составляет Lycopodium pungens, который является спутником ольховника, и в некоторых районах Boschniakia rossica, которая на нем паразитирует. Точно также флористические наборы в верхней части (щебнистой) ольховников, ивняков и открытых участков имеют различия не большие, чем соответствующие ценозы между собой.
Примеров такого рода можно привести множество, и все они говорят лишь о том, что не следует переоценивать индикаторную роль не только отдельных видов, но и систематического состава целых сообществ, пока таковая не будет доказана однозначно. Здесь мы вовсе не хотим сказать, что сообщества (по крайней мере, там, где они имеются, т. е. могут быть отличны от группировок растений, не являющихся сообществами, не индицируют условий местообитаний. Однако, следует отчетливо представлять, что индикация сообществами условий среды в тундрах во многих случаях основана на сходной толерантности эдификаторных видов. Но самое суждение о сходстве толерантности этих видов основывается на визуальных оценках условий их обитания. Поэтому пока не будут выработаны критерии для сравнения местообитаний, совершенно необоснованно утверждать, что различные по систематическому составу сообщества (в том числе и монодоминантные) индицируют различную экологию местообитаний. Говоря языком биометрии, ошибка в такого рода суждениях может превышать результат. Понятие ЛЭ заключает то преимущество, что в силу своей объемности устраняет возможность обобщать единичные наблюдения до «закономерностей». По этой же причине (объемности) оно объединяет сообщества и группировки по многофакторному принципу, а не на основе иерархической классификации, что по мнению Д. Уэбба (Webb, 1954) более соответствует природным являениям. Д. Уэбб вполне справедливо считает, что отношения во временной серии сообществ аналогичны отношениям между эмбрионом и взрослым организмом, выросшим из этого эмбриона, а не отношениям между родителями его отпрыском. Другими словами, на одной и той же генетической основе развивающийся объект имеет весьма широкую амплитуду варьирования. В применении к сукцессивному ряду сообществ, это положение означает, что итогом развития на каждый данный момент может быть любое сообщество из некоторого множества их.
Таким образом, если уж говорится об иерархической класси фикации, то в это понятие не должен вкладываться тот же смысл, что и при родстве двух организмов. Необходимость принципа многофакторности Д. Уэбб показывает на следующем примере. Примем во внимание четыре типа сообществ: (1) верещатник, (2) болото сфагновое, (3) луг на мелах, (4) болото на известняках. (1) и (2) сходны по кислотности и торфянистости почв; (3) и (4) сходны по щелочности и минеральным почвам. Но
(1) и (3) —сухие, а (2) и (4) —сырые. Является ли (1) связанным с (2) или с (3) или же (3) более связано с (1) или (4)?
Вопросы не имеют смысла, так как по одному фактору классификация будет выглядеть так, а по другому -- иначе. Используя концепцию ЛЭ, мы должны объединить (1), (2) и (4) и противопоставить это объединение лугу на мелах. Вопрос,
о классификации растительности в пределах одного ЛЭ является
для нас второстепенным — мы можем заострять на нем внимание или оставить открытым. Синэкологические особенности ЛЭ
учитываются лишь в общей форме, а основной упор делается
на аутэкологические проблемы.
Отдельные особи какоголибо активного в районе вида могут встречаться в ЛЭ, совершенно чуждом для него, поселяясь, например, на некоторых формах микро- и даже нанорельефа. Можно думать, что экофон особи в таких случаях близок к ЛЭ, где этот вид процветает. Это предположение не является обязательным, и в ряде случаев экологическое сходство может быть лишь кажущимся. Нельзя сказать (если этот случай не массовый), в деиствительности ли условия на таких микроформах рельефа соответствуют условиям ЛЭ, определенно являющимися оптимальными для этого вида, или он расширяет здесь широту своей экологической амплитуды в результате формирования новых эколого-физиологических рас. Экологический оптимум любого вида находится в одном ЛЭ даже в случае, если вид встречается на нескольких ЛЭ, из этого правила очень мало исключений.
ЛЭ, как это следует из его названия, является понятием экологии ландшафта, так как среда на нем оценивается с точкизрения объективных условий для произрастания растений. Поэтому следует стремиться определять ЛЭ по признакам самой среды и не полагаться на индикацию растений, которая может использоваться лишь как дополнительный аргумент характеристики среды. Сказанное можно пояснить следующим примером. Исследователь, начинающий работать в бассейне р. Чантальвеергын, несомненно, выделит, как самостоятельный ЛЭ, щебнисто-мелкоземистые участки южных склонов гор и каменисто-песчаные южные склончики моренных гряд, имеющие степоидную растиительность. Но если исследователь начнет работать в 200 км от Телекая к востоку, он может и не обратить особого внимания на те же самые каменисто-песчаные южные склоны тех же самых древних отложений, поскольку здесь они мало интересны в ботаническом отношении. Нельзя утверждать, что это тот же самый ЛЭ, что и в долине Чантальвеергын (хотя отложения по всей вероятности те же самые), но можно рассматривать все поверхности с подобными отложениями как местный ЛЭ, по набору видов существенно отличный от аналогичного ЛЭ в бассейне Чантальвеергын и, вероятно, имеющий отличный индекс среды. ЛЭ является топологической единицей в наибольшей степени гомеостатичной, т. е. мезоклимат его (в ландшафтном отношении серии подурочищ нескольких урочищ) более постоянен в исторической перспективе, чем местный климат (в данном районе) и микроклиматы различных местообитаний. В Арктике и Субарктике относительное постоянство мезоклимата имеет тем большее значение для всего живого, что общий климат может подвергаться несистематическим резким колебаниям. Внутрисезонная изменчивость погод более обычна, чем редкие сезонные «потрясения» климата, но она может лишь в малой степени кардинально воздействовать на весь мезоклимат ЛЭ. Несистематическое сильное воздействие или направленное изменение макроклимата смягчится плавной постепенностью микроклиматов — их изменением по типу частичной «перестановки слагаемых». Изменение микроклимата на одних местообитаниях приводит к тому, что их среда становится близкой среде других местообитаний этого ЛЭ до начала общего изменения. Другими словами, здесь проявляется инвариантность ЛЭ — общее разнообразие экологических режимов (ЭМ) в пределах ЛЭ в значительно меньшей степени подвергается изменениям, чем экологический режим каждого отдельного местообитания. Это дает возможность части видов растений с узким диапазоном толерантности существовать в районе, где происходит изменение климата, переселяясь с одних местообитаний на другие в пределах какого-либо ЛЭ. Здесь уместно привести слова В. Б. Сочавы (1974. С. 51): «... постоянство и разнообразие взаимосвязаны. Чем разнообразнее экосистема, тем она стабильнее».
Инвариантность ЛЭ способствует не только «удержанию» малочисленных видов при изменениях климата, но и проявлению интрогрессии среди близких видов, особенно имеющих крупные популяции. При этом скрещивающиеся формы могут обитать в разных ЛЭ, тогда их потомство может заселить, по крайней мере, ту совокупность местообитаний, которая входит в оба ЛЭ с возможным поселением и в пределах других ЛЭ. Исторические изменения среды создают региональный физико-географический фон, являющийся по отношению к живым организмам экологическим фоном, который определяет «быть или не быть» в данном районе аборигенным видам и территориально соседним. Согласно X . Мэзону (Mason, 1954), на новые территории мигрируют преадаптированные биотипы, т. е. такие, которые физиологически пригодны для заселения новых местообитаний. Преадаптация понимается Мэзоном как результат генетической предопределенности особей, вырабатывающейся под влиянием среды обитания на родительские особи. Таким образом, расселение вида происходит постепенно, шаг за шагом, в результате физиологической изменчивости популяций. География вида обусловлена его экологией.

