Структура и морфогенез кустарников
М. Т. МАЗУРЕНКО, А. П. ХОХРЯКОВ © 2010
Глава III
Циклы развития побегов и их систем. Окончание
Симподиальная группа>
|
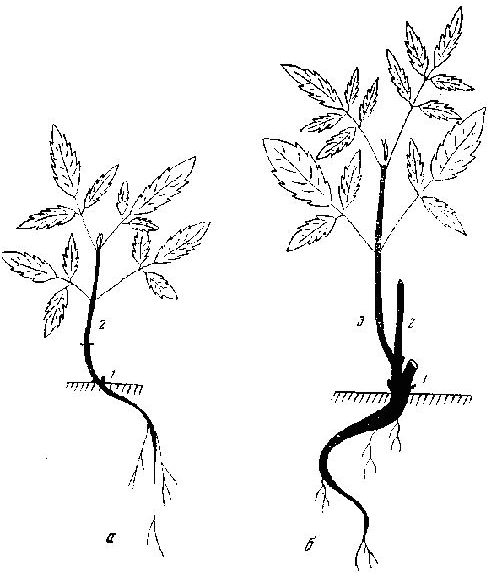
Рис. 47. Бузина кистистая (красная)
а — фаза нарастания первичной оси; б — фаза кущения; 1 — первый год развития; 2 — второй; 3 — третий год развития
|
Бузина красная — Sambucus racemosa L. Подмосковье. В старых парках. 1971-1973. Высокий малоствольный, часто деревцевидный кустарник, до 3 м, с кроной из 3-5 ярусов. Проростки 2-2,5 см, с 2 парами тройчатых и пятерных листьев. Нарастание первичной оси длиться 2, реже 3 года с приростами 2-5 см или она остается простой. На следующий год по окончании роста первичный побег отмирает, за исключением 1-2 нижних узлов, откуда отрастает побег кущения (турион) в 10-15 см, также с 2 парами сложных листьев. По окончании вегетации он отмирает, за исключением базальной части в 1-1,5 см с 1 нижним узлом. Затем ежегодно появляются все более крупные побеги кущения со все большей остающейся живой частью. К пятому - седьмому году очередной турион достигает 20-30 см, имея 5-7 пар листьев. В конце вегетации у него отмирает только верхняя половина. На следующий год от живой его части отходят 1-2 пары побегов ветвления, 10-20 см, с 2-3 парами листьев, появляется, следовательно, первая СПФ, основной цикл которой длится всего 2 года (рис. 47). Побеги формирования пятого — восьмого порядков появляются уже в числе 2, до 60 см, к концу вегетации отмирает только верхняя их часть (10-15 см). В СПФ развивается 2 порядка побегов ветвления, основной цикл длится 3 года. Новые турионы отрастают не только от основания, но и от средних частей предыдущих, формируя ССО. К десяти - пятнадцатилетнему возрасту растение достигает почти предельной величины в 2-2,5 м и состоит из 20-30 СПФ разного возраста. Турионы, идущие от нижней части куста, до 1,5 м, с изогнутой горизонтально верхушкой. Основной цикл длится до 5 лет. Побеги ветвления первого порядка, сидящие близ живой вершины туриона, закрытые, с ажурным головчатым соцветием, 20-30 см, с 3-4 парами листьев. Ниже них на турионе находятся вегетативные побеги ветвления, 40-60 см, с 4-6 парами листьев. Иногда от одного узла отрастает пучок побегов из главной и дополнительных пазушных почек. К концу вегетации слабые генеративные побеги и верхушки сильных генеративных и вегетативных побегов отмирают. На живой их части, а также в основании генеративных побегов на турионе отрастает следующая генерация вегетативных и генеративных побегов ветвления, но уже меньших размеров. Всего бывает 3-4 порядка ветвей, последний из которых состоит из мелких вегетативных побегов. После этого верхушка СПФ со всеми побегами ветвления отмирает, а нижняя часть туриона входит в состав CСO благодаря развитию на ней дочернего побега формирования. В основании куста в этот период турионы, как правило, не образуются. Дочерние СПФ выносятся всё выше, а в нижней части скелетные оси (их обычно 2-4) постепенно оголяются. |
В первое время по достижении максимального размера куста ростовые процессы еще преобладают, образуется все больше СПФ. В середине основного цикла, на третий - четвертый год все более активно образуются побеги дополнения, отрастающие пучком в основании побегов ветвления первого - второго порядков. Первый период основного цикла — «рост» — длится 2 года, так как побеги ветвления первого порядка также выполняют ростовые функции. Период «отмирание» длится последние пятый и шестой годы. К 25-30 годам число новообразующихся СПФ сокращается. Они начинают появляться на скелетных осях из почек, покоившихся не менее 10 лет, но большая часть их недолговечна и многие эфемерны. Размеры их сокращаются до 60 см, побеги ветвления и дополнения большей частью вегетативные. Вследствие сокращения основного цикла максимум до 3-4 лет крона редеет. Отмирают не только СПФ, но и скелетные оси и крупные сучья. В конце онтогенеза, как правило, живым остается один стволик. В основании его ежегодно появляются хлыстовидные эфемерные побеги до 1 м. Онтогенез заканчивается полным отмиранием ССО. Его фазы: 1) рост первичной оси (первые 1-3 года), 2) кущение и усиленный рост (до 7-10 лет), 3) возникновение ярусности кроны (ССО) и достижение максимальной высоты (от 7-10 до 20 лет), 4) «стабилизация» — равновесие роста и отмирания (20-30 лет), 5) преобладание отмирания, возникновение деревцевидной формы и конец онтогенеза (до 50 лет).
Леспедеза двуцветная — Lespedeza bicolor Turz. Приморский край, окр. Владивостока. Светлые дубняки и их опушки. 1975. Малоствольный аэроксильный кустарник, с длинными полуплакучими ветвями, до 2,5 м. Прорастание надземное, Размеры стебля в первую вегетацию сильно зависят от условий. В хороших — он достигает 7-10, иногда до 20 см. По мере роста развертываются все новые листья, верхушечные — осенью, уже тогда, когда нижние опадают. В конце вегетации побег четко разграничен на тонкую зеленую верхнюю часть без почек или с очень маленькими пазушными почками и нижнюю, одетую перидермой, с крупными часто сериальными почками. Граница между ними очень резкая. В хороших условиях существования соотношение этих частей примерно 3 : 1, в плохих — больше. Травянистая часть не перезимовывает. Весной из самой верхней живой почки отрастает дочерний побег с увеличенной многолетней частью. Иногда образуется 2 дочерних побега, тогда один из них, обычно верхний, более мелкий — эфемерный. На третий год многолетняя часть эфемероидного побега достигает более половины — около 10 см. Так развивается типичная деревцевидная форма аэроксильного куста. В неблагоприятных условиях, особенно на сильно вытаптываемых или каменистых жарких местах, многолетние части побегов нарастают очень медленно. Происходит типичное кущение с возрастанием порядка и размера турионов на пятый—седьмой год до 50 см, после чего медленно, но неуклонно многолетняя часть побегов все же начинает увеличиваться. В обоих случаях корневая система остается стержневой, придаточных корней но образуется, но в последнем случае с самого начала формируется более или менее типичный куст из 2-3 скелетных осей — стволиков. Встречаются, разумеется, и различные переходные формы между типично «кустовидной» и «деревцевидной».
На четвертый - пятый год деревцевидная форма начинает ветвиться, так как на побеге предыдущего порядка отрастают 2, редко 3 эфемероидных побега, 25 см. Примерно тогда же в верхней их части силлептически появляются небольшие веточки с цветками. Соцветие занимает верхнюю часть побега (3-5 см). Ниже него располагаются вегетативные ветки, также возникшие силлептически. Все они помещаются на однолетней части побега. Эфемерные побеги отрастают и из спящих почек на многолетней уже части куста, на его стволике в основании ветвей. В дальнейшем идет рост как самих побегов, так и генеративной их части. Многолетняя часть более половины побега не занимает.
В 10-15 лет леспедеза представляет собой разветвленный ку стик с коротким, 10-25 см, стволиком, составленным нижними частями первых 5-7 порядков побегов, отмершие остатки которых у молодых кустов выглядят как короткие «пеньки». Нарастание ветвей идет по следующему типу: из верхних почек па побеге предыдущего порядка отрастают 2-3, редко до 5 длинных эфемероидных побегов, до 50-100 см, с поникающей или даже плакучей верхушкой из многочисленных силлептических ветвей соцветия до третьего порядка, заканчивающихся длинными кистями. Нижние ветви могут быть вегетативными и располагаться пучками. Все они находятся на однолетней, эфемерной части побега. На многолетней части силлептических побегов не отрастает, лишь в его основании иногда появляется несколько эфемерных и вегетативных. На следующий год из верхних живых 3-5 почек многолетней части (20-30 см) вновь отрастают описанные выше эфемероидные побеги.
|
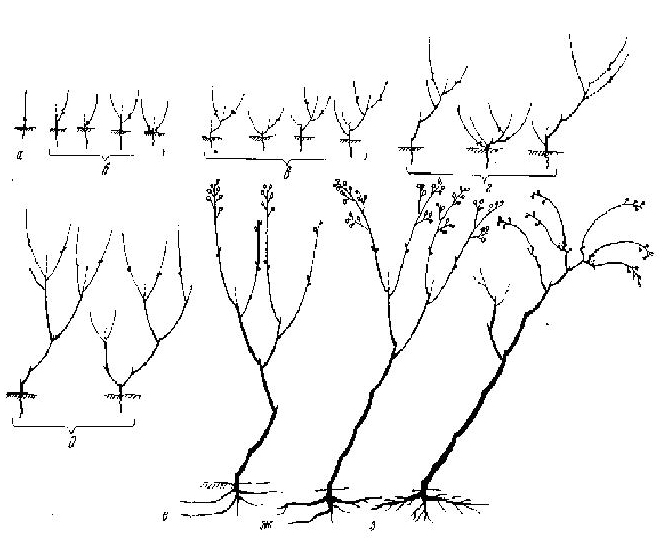
Рис. 48. Леспедеза а — з — фазы морфогенеза
|
Разделение функций (ростовой и скелетной, с одной стороны, и ассимиляции и плодоношения, с другой) здесь связано с приуроченностью не столько к разного типа побегам, сколько к разным частям одного туриона, верхняя эфемерная часть которого представляет собой не что иное, как силлептически развившуюся СПФ или «систему годичного побега» (Гатцук, 1967).
Из 2-5 дочерних СПФ 1-2 оказываются более сильными и долговечными, другие, менее сильные, дав еще 1-2 разветвления, отмирают. Таким образом, куст в возрасте около 10 лет имеет около 10-15 концевых разветвлений. В дальнейшем размеры побегов продолжают увеличиваться, особенно размеры соцветий, которые становятся все более плакучими. Однако куст, достигнув 2-2,5 м (редко - более), растет далее уже крайне медленно, так как размеры многолетней части турионов начинают быстро падать. В конце концов они отмирают почти до основания, так что последующие порядки ветвей располагаются на предыдущих как бы пучками. Всего в кроне развивается до 30-40 таких почти эфемерных побегов. Генеративная часть и общие их размеры сокращаются. Из многочисленных спящих почек начинают отрастать вегетативные побеги дополнения, до 20 см, полностью эфемерные. В конце концов куст обычно полностью засыхает. Но в некоторых случаях от основания стволиков начинают отрастать новые турионы, в числе 1-3, почти до 1 м, начинающие развитие новой ССО. Однако цикл ее более короток, чем первой, не превышает 10 лет, что связано, очевидно, с отсутствием придаточных корневых систем (рис. 48). |
Фазы онтогенеза: 1) первичной оси (первый год), 2) усиленного роста многолетней части ростовых побегов, включая кущение (до 5 лет); 3) стабильности многолетней части побегов и роста многолетней части куста (ССО) (до 10-15 лет); 4) стабильности многолетней части куста и уменьшения многолетней части побегов вплоть до полного отмирания ССО; 5) развития ССО второго порядка (факультативная).
|
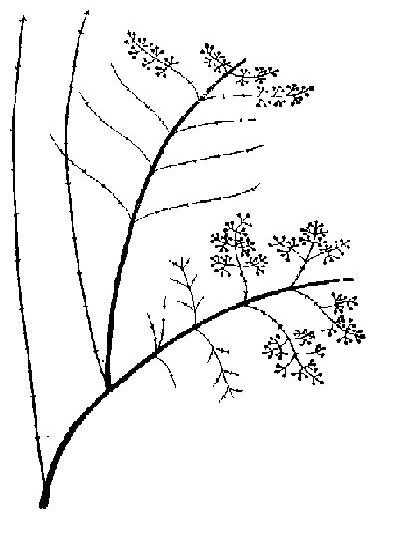
Рис. 49. Дейция. СПФ на третий год развития
|
Дейция — Deutzia scabra Thunb. Батумский ботанический сад. Самосев среди посадок декоративных экзотов. 1973-1975. Плотный раскидистый кустарник из множества разновозрастных скелетных осей, до 3, реже 4 м. Проросток до 3 см, с 3-4 листьями 1,0х0,6 см. Первичная ось нарастает 2-3 года, достигая 6-8 см. Уже на второй год в ее основании отрастает первый побег кущения, 5-7, иногда до 15 см, с 3-5 листьями. Обычно он простой, изредка дает второй прирост, 2-3 см. Основание первичной оси в это время утолщается, и одна из почек на нем производит следующий побег кущения, до 20 см, с 5-7 листьями 3,2х1,5 см. На четвертый год отрастают побеги кущения второго - третьего порядков, до 40 см, с приростами 3-5 см и боковыми косо вверх направленными веточками, 2-10 см. К пятому - шестому годам первичная ось и побеги кущения первого порядка отмирают, развиваются СПФ третьего порядка, до 80, иногда 100 см, примерно с 20 листьями, достигшими предельных размеров 12х4 см. Основной цикл их длится 3 года. Ежегодно размеры побегов и СПФ в целом увеличиваются в зоне кущения появляется по 2-3 новых туриона, куст разрастается не только в высоту, но и в ширину. С седьмого года побеги формирования отрастают не только от основания куста, но и от нижних частей турионов предыдущего порядка, которые таким образом становятся ССО. Побеги формирования в первый год вырастают от 1 до 2 м, часто дают второй прирост 3-7 см, представляющий собой обычно разветвленное метельчатое соцветие. Верхушка его горизонтально изгибается, ближе к ней появляются преимущественно генеративные ветви с метельчатыми соцветиями, дальше от него — вегетативные. Первые на следующий год полностью отмирают, вторые обычно заканчиваются терминальным соцветием. На первом их приросте ближе к вершине появляются генеративные, б-8 см, и далее более длинные вегетативные побеги ветвления второго порядка.
Уже на второй, а также и на третий годы основного цикла от нижней части первого туриона отрастают несколько дочерних, образующих второй ярус СПФ, в то время как первая постепенно отмирает. Благодаря активному появлению турионов нарастание ярусов в кроне ССО идет очень быстро, но вследствие гибкости турионов они быстро наклоняются книзу, чему очень способствуют снегопады, поэтому ярусы СПФ, нарастающие сверху, как бы спрессовывают находящиеся под ними (рис. 49). К 10-15 годам кусты вырастают до 4 м, ветви их раскидываются в стороны до 1 м от центра куста. В кусте множество разновозрастных осой, живых и отмирающих ССО, особенно в центре. Многие турионы эфемерны, другие — производят лишь 1-2 порядка побегов ветвления. К 20-25 годам даже в СПФ, проходящих полный основной цикл (за 3 года), большинство побегов ветвления |
вегетативные. Цветение и плодоношение ослабляются. Все более увеличивается количество недолговечных турионов и СПФ, в середине куста все в большем количестве концентрируются отмершие и отмирающие CСО. К 30 годам куст отмирает полностью. Фазы онтогенеза: 1) нарастание первичной оси; 2) усиленный рост вновь образуемых турионов и СПФ; 3) появление ССО и ярусности; 4) стабилизация; 5) отмирание куста.
|

Рис. 50. Рябинолистник. Видно корневище, отходящее от основания скелетной оси |
Рябинолистник — Sorbaria sorbifolia (L.) A. Br. Приморский край, Супутинский (Уссурийский) заповедник. Смешанные леса, в сырых низинах. Проростки находились по краям лесных дорог. 1975. Раскидистый кустарник с крупными белыми соцветиями и перистыми л истьями. Прорастание надземное. Первичный побег в первый год достигает 5-10 см, листья несколько сближены у верхушки. Прирост второго года — 2-3 см, третьего, если он есть,1-2 см. На приросте первого, реже второго года могут образоваться короткие боковые побеги. На второй - третий годы от основания первичного побега, иногда на высоте 2-3 см от поверхности почвы, отрастает первый турион, 10-15 см, также нарастающий 2-3 года с быстрым падением последующих приростов. На третий - четвертый год появляется второй, еще более крупный турион того же характера. Примерно с пятого года от основания первичного куста отрастают турионы,
нижняя часть которых, около 5 см, плагиотропная, корневищная. В последующие годы отрастают еще 2-3 более длинных корневища, до 20 см, с придаточными корнями. Побеги формирования (оси парциляции), которыми они заканчиваются, всё более крупные, до 0,5 м, производят боковые ветви второго - третьего порядков, заканчивающиеся соцветиями, и дочерние турионы у своего основания в числе 2-3 или реже — 1-2 от середины. Таким образом возникают парциальные кусты, состоящие из 1-2 ССО и 2-3 СПФ, с основными циклами, проходящими за 2-4 года, циклами ССО, проходящими за 4-6 лет, и циклами парциальных кустов, проходящими за 10 лет. На более поздних фазах онтогенеза формирование парциального куста начинается с роста от основания одного из турионов длинного, достигающего 1,5-2 м корневища с белым растущим концом в 20-30 см и остальной частью, покрытой перидермой, с редкими, но многочисленными придаточными корнями. В случае повреждения растущего конца возможно ветвление корневища. Выходя на поверхность, оно дает надземный прирост, достигающий 1,5 м и заканчивающийся крупным пышным соцветием. На следующий год верхняя треть или четверть побега отмирает, под отмершей частью отрастают 3-5 ветвей (иногда по две вместе из одного узла), до 1 м, также заканчивающихся крупным соцветием, с нижними его ветвями, развивающимися силлептически из пазух верхних листьев (рис. 50). На третий год основного цикла отрастает вторая генерация побегов ветвления как на самих ветвях первого порядка, так и у их основания на турионе. Размеры их еще меньше — около 50 см. Всего развивается до 5 генераций и порядков побегов ветвления, последние из которых 10-20 см, вегетативные и эфемерные. Вся СПФ при этом сильно наклоняется, конечные ее ветви имеют как бы восходящее направление роста. В месте перегиба СПФ на верхней части туриона начинает рост дочерний турион, формирующий ССО и развивающий свою СПФ, но с более коротким основным циклом (4-5 лет). Всего в ССО бывает до 3 ярусов СПФ, и цикл ее развития длится немногим более 10 лет. Однако за это время турионы отрастают и от основания материнского побега, продолжая цикл развития парциального куста, длительность которого 15-20 лет. Фазы онтогенеза: 1) первичной оси и СПП (до 3 лет), 2) первичного куста (до 10 лет), 3) системы парциальных кустов и клона (неопределенно долго). В окрестностях Владивостока в посадках вдоль дорог кусты рябинолистника достигают 3 м, число ярусов в кроне — до 5, размеры турионов, идущих от оснований куста или корневищ, до 2 м, в СПФ 5-6 порядков побегов ветвления, основные циклы длятся 6-7 лет, циклы ССО — 20-25 лет. Напротив, в окр. Магадана рябинолистник не превышает 1,5 м, побеги формирования - 1м, число порядков побегов ветвлений в СПФ — 2-3, редко 4, цикл развития ССО менее 10 лет, парциальных кустов — около 15 лет. |
Черника — Vacсinium myrtillus L. По данным И. Г. Серебрякова (1962) и собственным наблюдениям в Подмосковье и Ленинградской обл. (1970—1975). Листопадный кустарничек, до 30 см, с вечнозелеными ветвями. Прорастание надземное, проросток до 1 см, нарастание первичной оси идет по симподиалъному типу в течение 2-5 лет с приростами (побегами) до 2 см. На третий год в основании се отрастают несколько восходящих побегов из спящих почек (кущения), 1,5-2 см, которые служат основой для формирования ложных побегов, нарастающих 3-4 года. В их основании, в свою очередь, появляются побеги из спящих почек с еще более ярко выраженной восходящей частью или плагиотропной, до 5 см, покрытой иногда придаточными корнями, с той же интенсивностью приростов, нарастающих 2-3 года. В результате на пятый — восьмой год образуется небольшой стержнекорневой кустик высотой около 10 см, с уже отмершей первичной осью. На восьмой - десятый год от его основания отрастает несколько корневищ 10-15 см, а затем и более, до 50 см, с придаточными корнями. Выйдя на поверхность, концы этих корневищ дают олиственные приросты, достигающие за один период роста 5-8, см. Близ их верхушек отрастают уже не 1, а 2-3 побега замещения, формирующих как главную ось, так и боковые ветви. Главная ось симподиально нарастает не более 4-5 лет, достигая общей высоты до 30 см, с постепенным падением ежегодных замещающих побегов. Ветвление ее акросимподиальное, идет до пятого - шестого порядков с постепенным падением размеров ветвей. Весной из верхних 2-3 почек побега предыдущего (второго - третьего) порядка начинается рост новых побегов, около 5 см, с 5-7 листьями. В пазухах 3-5 нижних листьев находятся цветки; 2-3 верхних — крупные пазушные почки, из которых отрастают побеги следующего порядка. На четвертый - пятый годы, когда верхушка главной оси ложного туриона отмирает (но идет еще ветвление СПФ), от ее основания или на высоте 5-7 см от поверхности почвы отрастает дочерний турион несколько более мощный, чем материнский, производящий дочернюю СПФ. Дочерние СПФ (2-3, редко более) 1, иногда 2 порядков формируют парциальный куст с 1-2 ССО. Основные циклы длятся около 10 лет. Циклы развития ССО, если они формируются, — 15-20 лет, а циклы развития всего парциального куста — до 50 лет. На четвертом — шестом году существования парциального куста (или его первой СПФ) от основания туриона первого порядка отрастают длинные, до 1,5 м, корневища, нарастающие в почве 6-7 лет. Выходя на поверхность, корневище превращается в надземный турион, дающий начало новому парциальному кусту. Этапы онтогенеза: 1) первичная ось (первые 2-3 года), 2) первичный куст (первые 5-10 лет), 3) система парциальных кустов и клон (от 10 лет и неопределенно долго).
Рассмотренные нами онтогенезы 17 видов различных кустарников показывают насколько разнообразны не только их побеговые системы и циклы их развития, но и так называемые большие жизненные циклы в целом. При всем том нельзя не отметить, что в онтогенезе кустарников, насколько бы ни были они непохожи друг на друга, всегда есть и общие элементы. У всех вначале развивается стержнекорневой первичный побег, затем — система первичного побега, у всех в той или иной фазе «просыпаются» спящие почки, у большинства образуются ростовые побеги, составные скелетные оси, у многих в той или иной степени выражена фаза придаточного корнеобразования и клона. При этом каждый из рассмотренных видов характеризуется своим, иногда довольно оригинальным сочетанием разных фаз и более длительным течением какой-либо одной, редко двух, фаз (в этом последнем случае одна из них чаще всего фаза клона). Это и позволяет классифицировать жизненные формы кустарников, поскольку они зависят, конечно, в основном от строения и особенностей наиболее длительной фазы. Ясно, что кустарники с отсутствующей или слабо выраженной фазой кущения будут относиться к аэроксильным, а с хорошо выраженной — к геоксильным.
В каждой из трех выделенных групп по типу СПФ и СПП кустарники очень разнообразны, имеют между собой многочисленные переходы, но вместе с тем нельзя не отмстить, что в первой преобладают типы с длительным развитием первичного побега и СПП, а в третьей — с быстро проходящим периодом СПП. Это, повидимому, свидетельствует об ускоренном течении онтогенеза в данной группе, подтверждением чему служит и более активная смена в ней СПФ и более интенсивное отмирание побегов.
* * *
В данной работе приведены сведения о строении и циклах развития более 100 видов кустарников и кустарничков различных как в систематическом, так и в эколого-географическом отношении. При этом мы не можем указать на какие-либо особенности, которые были бы свойственны исключительно какой-либо одной группе, что, очевидно, свидетельствует о весьма общем и широком значении выявленных структур и циклов развития. Всего нами были выделены следующие уровни структур и соответствующие им циклы развития: 1) приросты, 2) побеги, 3) системы побегов первого порядка (СПП, СПФ, СПВ), 4) системы побегов второго порядка (ССО), 5) системы побегов третьего порядка (куст в целом), 6) совокупности этих кустов (клоны). Их можно разделить на две неравные части: в первую войдут уровни 1 и 2, с простыми неразветвленными осями, во вторую — все остальные, с разветвленными.
Ветвление в данном случае выступает как способ полимеризации осевых систем все более высокого ранга, который уже на третьем уровне может приводить к образованию кустовидных форм. Следует оговориться, что ветвление, а следовательно, и образование кустовидных форм, свойственно всем прикрепленным организмам, как низшим и высшим растениям, так и животным (губки, кораллы, мшанки и др.) и даже некоторым неживым объектам (дендриты). Следовательно, ветвление можно рассматривать как способ полимеризации всех систем, независимо от их природы. В органическом мире оно до некоторой степени компенсирует неподвижность прикрепленных форм, и именно это служит одним из стимулов его развития и совершенствования.
По внешнему виду описанные кустарники могут очень сильно различаться между собой. Способ и направление роста, степень отмирания и долговечность их побегов у разных видов могут быть очень различными. По этим признакам издавна выделяют группы аэроксильных и геоксильных кустарников, стлаников и лиан, полукустарников и кустарничков. Но все же основные различия между всеми этими формами роста, как нам кажется, лежат в той степени сложности побеговых систем и циклов их развития, которой они достигают. Так, есть формы роста, представляющие собой, собственно, один первичный побег, или даже прирост (тип пальм, древовидных лобелий и других «колонновидных» растений), или систему первичного побега, например, в случае трагакантов, а также большинство подушковидных стержнекорневых растений от деревянистых до травянистых. Большинство аэроксильных кустарников представлены системами первичного побега или 1-2 СПФ, геоксильные достигают пятого порядка полимеризации. Совершенно очевидно, что клонообразующие (ветве- укореняющиеся и корневищные ксилоризомные) кустарники нуждаются в выделении в особую группу, достигающую наивысшей, шестой ступени полимеризации побеговых систем.
Данная классификация ни в коем случае не может рассматриваться как некое указание на эволюцию жизненных форм растений по пути от древесных к травянистым. Трансформация их в этом направлении связана со своими закономерностями и к полимеризации побеговых систем имеет лишь косвенное отношение. Здесь можно указать лишь на то, что по строению побеговых систем и циклам их развития так называемые вторично-древесные растения, к которым причисляют трагаканты, полыни, все солянковые, гвоздичные, крестоцветные, губоцветные и некоторые другие, совершенно невозможно отличить от первично-древесных. Это, по нашему мнению, свидетельствует о единстве природы тех и других и, следовательно, об отсутствии вторичного одревеснения первично-травянистых форм. В дальнейшем было бы чрезвычайно интересно применить принцип степени полимеризации побеговых систем для классификации жизненных форм как древесных, так и травянистых растений с целью выявления между ними как сходства, так и различия по этому признаку.
Литература
Алексеев Ю. Е. 1959. Морфология прорастания некоторых видов рода Сеrasus. — Биологические науки, № 4.
Алексеев Ю. Е. 1963. Морфолого-биологические особенности образования кустарниковой формы у степной вишни. — Бюл. МОИП, отд. биол., т. 68. вып. 3, 155.
Алексеев Ю. Е. 1974. Вишня степная. — В сб.: Биологическая флора Московской области, вып. 1. С.78—87.
Барыкина Р. П. 1971. Морфолого-анатомические исследования барбариса обыкновенного и барбариса тунберга в связи с вопросом преобразования жизненных форм в семействе барбарисовых. — В сб.: Морфология цветковых. М., «Наука».
Булыгин Н. Е. 1965. Динамика формирования цветочных зачатков у древесных растений в Ленинграде. Автореф. канд. дис. Л.
Вахрамеева М. Г., Денисова Л. В. 1974. Волчье лыко обыкновенное. — В сб.: Биологическая флора Московской обл., вып. 1, 124—130.
Гаврилюк В. А. 1966. К биологии растений юго-востока Чукотского полуострова. — В сб.: Приспособление растений Арктики к условиям среды. М., «Наука».
Гатцук Л. Е. 1967. Морфогенез копеечника кустарникового Hedysarum frutioсosum Pall. при постоянном уровне песчаного субстрата. — В сб.: Онтогенез и возрастной состав популяций цветковых растений. М., «Наука», 9-34.
Гатцук Л. Е. 1970. Элементы структуры жизненных форм геммаксилярных растений и биоморфологический анализ копеечника кустарникового. Автореф. канд. дис. М.
Гатцук Л. Е. 1974а. Геммаксиллярные растения и система соподчиненных единиц их побегового тела. - Бюл. МОИП, отд. биол., т. 79, вып. 1, 100-113.
Гатцук Л. Е. 1974 б. К методам описания и определения жизненных форм в сезонном климате. — Бюл. МОИП, отд. биол., т. 79, вып. 3, 84—100.
Головкин Б. И. 1973. Переселение травянистых многолетников на полярный север. Л., «Наука».
Грудзинская И. А. 1960. Летнее побегообразование у древесных растений и его классификация. — Бот. журн., № 7.
Гурский А. В., Каневская И. Б., Остапович Л. Ф. 1953. Основные итоги интродукции растений в Памирском ботаническом саду. — Труды АН ТаджССР, т. 16. Душанбе, Изд-во АН ТаджССР.
Дервиз-Соколова Т. Г. 1966. Анатомо-морфологическое строение Salix polaris L. и S. phlebophylla And. — Бюл. МОИП, отд. биол., № 2.
Жуйкова И. В. 1969. Морфогенез и ветвление побегов. —В сб.: Экология и биология растений восточноевропейской лесотундры. Л., «Наука», 212— 227.
Зайцев Г. Н. 1962. Интродукция жимолости в Ленинграде. — Труды Бот. ин-та им. Комарова, серия 6, интродукция растений, вып. 8.
Закржевский Б. С., Коровин Е. П. 1935. Экологические особенности главнейших растений Бетпак-Дала, — Труды Среднеазиатского гос. университета, серия 8, ботаника, т. 23.
Колищук В. Г. 1968. О морфологической эволюции от деревьев к травам в ряду стелющихся форм растений. - Бот. журн., т. 53, № 8.
Колищук В. Г. 1970. Стелющиеся древесные растения (эколого-морфологический анализ). Докт. дис. Львов.
Крылова И. Л. 1958. Развитие замещающих побегов у сосны и бука. — Бюл. МОИП, отд. биол., т. 63, вып. 3.
Кудряшов Л. В., Барыкина Р. П. 1970. Морфогенез и старение взрослых растений, относящихся к жизненной форме стланиковых кустарников. — Рефер. докл. на Всесоюз. симпозиуме морфологических основ и онтогенеза высших травянистых растений. Ставрополь.
Курындин И. И., Малиновский В. В., Венъяминов А. Н ., Бенохозов И. В. 1947. Плодоводство. М;, Сельхозгиз.
Лучник З.И. 1960. Обрезка кустарников. М., Сельхозгиз, с. 96.
Любименко В. И. 1900. О спящих почках. — Труды Спб. об-ва естествоиспытателей, т. 30, отд. бот., вып. 3.
Ляшенко Н. И. 1964. Биология спящих почек. М.— Л., «Наука».
Мазуренко М. Т. 1973а. Две формы роста жимолости этрусской в Крыму. — Бюл. ГБС, вып. 88
Мазуренко М. Т. 19736. Формы роста жимолости камчатской. — В сб.: Почвы и растительность мерзлотных районов СССР. Магадан, 244—250.
Мазуренко М. Т. 1974. Системы побегов и циклы их развития у кустарников семейства жимолостных. Автореф. канд. дис. М.
Мазуренко М. Т., Антропова Г. Л. 1973. Основные циклы трех форм багульника болотного с берегов Байкала и из окрестностей Магадана. — В сб.: Почвы и растительность мерзлотных районов СССР. Магадан, 225—233.
Мастинская Р. А. 1971. Морфогенез и некоторые биологические особенности ежевики сизой (Rubus caesius L.) на северо-восточной границе ареала. Автореф. канд. дис. М.
Михайлова Т. Д. 1972. Побегообразование и жизненная форма Astragalus macropodium Lipsky. — Биологические науки, № 6.
Надёжина Т. П. 1962. Особенности роста и развития некоторых трагакантовых астрагалов Копет-Дага и их фенологический цикл. — Труды ВИН АН СССР, серия 5, вып. 10, 65—118.
Нечаева Н. Т., Васильевская В. К., Антонова К. Г. 1973. Жизненные формы растений пустыни Кара-Кумы. М., «Наука».
Нухимовский Е. Л. 1970. Структурная организация и морфогенез некоторых семенных растений. Автореф. канд. дис М.
Нухимовский Е. Л. 1973. О соотношении понятий «партикуляция» и «вегетативное размножение». — Бюл. МОИП, отд. биол., т. 78, вып. 5, 107— 120.
Петров А. А. 1963. Что считать основным структурным элементом кроны древесных растений.— Докл. Болгарской АН, т. 16, № 5.
Родман Л. С. 1957. Образование кустарниковой формы у Rosa spinosissirna L. в условиях кустарниковых степей западных предгорий Алтая. — Бюл. МОИП, отд. биол., т. 62, вып. 1.
Рогов Р. А. 1960. Морфогенез жизненной формы кустарничка у Calluna vulgaris ( L .) Hill в условиях Прибалтики. — Бюл. МОИП, отд. биол., т. 65, вып. 2, 91—94.
Рогов Р. А. 1962. О бореальных полукустарничках. — Бюл. МОИП, отд. биол., т. 67, вып. 6.
Рогов Р. А. 1969. Биолого-морфологические особенности многолетних пустынных растений. М., «Наука».
Серебряков И. Г. 1949. Структура и ритм в жизни цветковых растений. — Бюл. МОИП, отд. биол., т. 54, вып. 1.
Серебряков И. Г. 1952. Морфология вегетативных органов высших растений. М., «Советская Наука».
Серебряков И. Г. 1954. Биолого-морфологический и филогенетический ана лиз жизненных форм покрытосеменных. — Уч. зап. МГПИ им. В. П. Потемкина, т. 37, вып. 2.
Серебряков И. Г. 1955. Основные направления эволюции жизненных форм у покрытосеменных растений. Бюл. МОИП, отд. биол., т. 60, вып. 3, 71—91.
Серебряков И. Г. 1962. Экологическая морфология растений. М., «Высшая школа».
Серебряков И. Г., Доманская Н. П., Родман Л. С. 1954. О морфогенезе жизненной формы кустарника на примере орешника. — Бюл. МОИП, т. 59, вып. 2.
Серебряков И. Г., Чернышева М. Б. 1955. О морфогенезе жизненной формы кустарничка у черники, брусники и некоторых болотных вересковых. — Бюл. МОИП, отд. биол., № 2.
Смирнова О. В., Торопова Н. А. 1972. Большой жизненный цикл Galeobdolon luteum ( L.) Huds. — Бюл. МОИП, отд. биол., т. 77, вып. 1, 76—87.
Соколова Н. П. 1974а. Эволюционные взаимоотношения жизненных форм в роде Rubus L. Автореф. докт. дис. М.
Соколова Н. П. 19746. Классификация и морфогенетические отношения жизненных форм рода Rubus L. — ежевика. — Докл. Моск. с.-х. акад. им. К. А. Тимирязева, вып. 204, 231—239.
Солоневич Н. Г. 1970. Эколого-биологическая характеристика растений (лесо– тундрового стационара БИН). Гл. IV. — В кн.: Экология и биология растений восточноевропейской лесотундры. Л., «Наука».
Федоров А. А., Кирпичников М. Э., Артюшекко 3. Т. 1962. Атлас по описательной морфологии высших растений. М.—Л., Изд-во АН СССР.
Шафранова Л. М. 1964. Жизненные формы и морфогенез лапчатки кустарниковой в различных условиях произрастания. — Бюл. МОИП, отд. биол., т. 69, вып. 4.
Шафранова Л. М. 1967. О некоторых возможных путях перехода от кустарников к травам. — Научные доклады высшей школы. Биологические науки, № 7.
Шафранова Л. М. 1968. Пути трансформации кустарниковой жизненной формы в роде лапчатка. — В сб.: Рефераты докладов Межвузовской конференции по морфогенезу растений. М.
Шафранова Л. М. 1970. О некоторых путях перехода от кустарников к травам в роде лапчатка — Potentilla L. Автореф. канд. дис. М.
Шитт П. Г. 1952. Биологические основы агротехники плодоводства. М., Сельхозгиз.
Dode L. А . 1905. Extraits dune monographie une dite du genere Populus. Extraits des Mem. Soc. histoire nater. autumn, t. 18. Paris.
Du-Rietz G. E. 1931. Life forms of terrestrial floweing plants. Uppsala.
Gimingham C. H. 1970. The morphology of vegetative regeneration in Calluna vulgaris. - New Phytologyst, v. 69, N3.
Lems K. 1962. Adaptive radiation in the Ericaceaea. I. Schoot development in the Andromedeae. — Ecology, v. 43, N 3, p. 524—528.
Spat H. L. 1912. Der Johannestrieb. Berlin.
Troll W. 1937. Vergleichende Morphologie der hoheren Pflanzen, Bd 1. Berlin.
Troll W., Weberling F. 4966. Die Infloreszenzen der Caprifoliaceen und ihre systematische Bedeutung. Wiesbaden.
Warming E. 1908. Om Planterigets Lifsformer. Kobenhavn.

