Структура и морфогенез кустарников
М. Т. МАЗУРЕНКО, А. П. ХОХРЯКОВ © 2010
Глава III
Изменение систем побегов в онтогенезе
Моноподиальная группа
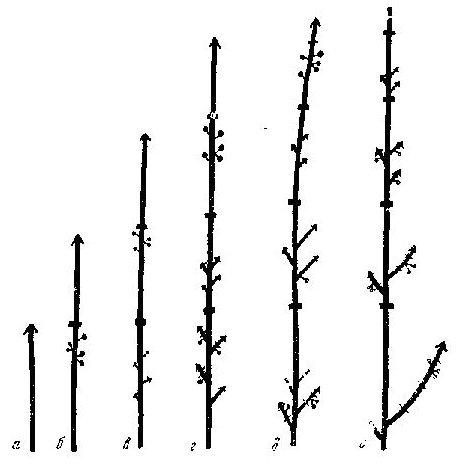
Рис. 39. Карагана гривастая. Нарастание побега и цикл развития его приростов
а — первый год; б — второй; в — третий; г — четвертый; д — пятый; е — шестой год
|
Карагана гривастая — Caragana jubata ( Pall .) Poir . 1. Бурятская АССР, Тункинский р-н, с. Монды. Галечники. 1970. Проростки первого года с 2 овальными довольно крупными семядолями и укороченным стеблем с 2-3 листьями. Последующие приросты небольшие, 2-3 мм, розеточного типа, но постепенно все более увеличивающиеся до 0,5 и 1 см. До 10 лет ветвления первичной оси не происходит, затем, по достижении ею 5-8 см, появляются первые, также розеточного типа побеги ветвления, нарастающие также в течение длительного времени. У растений 15-20 лет приросты достигают 1,5-2 см, но затем темп нарастания главной оси замедляется и верхушка ее может отмереть. Ее замещает ближайший боковой побег. К 25-30 годам могут развиться побеги из спящих почек, находящихся в нижней части первичной оси, но необязательно в ее основании. Рост их более интенсивен, до 5 см за год. К этому времени растение достигает 1,5 м, имея крону, начинающуюся почти от земли, но все же с главным стволиком в основании, постепенно расширяющуюся кверху, с разветвлениями не более 4 порядков. Все ветви покрыты плотной обкладкой из широких оснований листьев и прилистников, с далеко торчащими из нее колючками — неопадающими рахисами листьев. Цикл развития приростов главных ветвей таков: размеры их около 10 см, на второй год в верхней и средней их частях отрастают укороченные, до 4 см, побеги ветвления, наполовину скрытые в пазухах неопавших оснований листьев. Последующие их приросты равны предшествующему приросту, по всей длине которого развиваются укороченные пазушные цветоносы с 1-3 цветками (рис. 39). На каждом последующем приросте побега формирования продолжается развитие новых побегов ветвления и их рост в течение 4-5 лет, но все менее интенсивный. Цветки образуются только на их двухлетних приростах. На четвертый - пятый год побеги ветвления отмирают, но не опадают. Побег формирования нарастает 10-20 лет. Постепенно в его нижней части отваливаются отмершие побеги ветвления, прилистники и колючки. Основание стволика постепенно очищается, жизнедеятельная часть кроны переносится все выше и выше. |
2. Окр. Магадана, пос. Атарган. Каменистые приморские склоны. 1973. Стелющийся кустарничек с приподнимающимися ветвями. Наиболее часто встречаются кусты с 5-10, иногда (в конце онтогенеза) с 20 радиально распростертыми главными скелетными осями, 0,7-1,5 м. С началом вегетации раскрывается терминальная почка ветви и по мере ее роста распускаются цветочные почки, уже сформированные в прошлом году. Красивые, до 3 см, розовые цветки располагаются в 3-6 нижних пазухах листьев прошлогоднего прироста.

Caragana jubata
|
Верхняя часть прироста вегетативная, с 5-6 перистыми опушенными листьями, в пазухах которых формируются почки, раскрывающиеся на следующий год. К концу видимого роста конечный прирост имеет длину от 2 до 3 см. Вегетативные побеги ветвления отрастают из пазух листьев верхней части двухлетнего прироста, следующего за прошлогодним, в числе 2-3. Они небольшие, от 0,5 до 1 см. Если материнская ветвь лежачая, то они появляются только на ее верхней стороне. Боковые побеги развиваются по типу материнского побега т. е. цветки появляются только на прошлогодних приростах в их нижней части в числе 1-3. После цветения большая часть побегов ветвления отмирает, некоторые из них живут до 5 лет. Только единичные побеги набирают силу, превращаясь в скелетные оси второго порядка. Их развитие идет по типу материнского побега. На скелетных осях второго порядка иногда образуются оси третьего порядка. Размеры их убывают. Например, средние размеры главных скелетных осей 1 м, осей второго порядка — 20-30 см, третьего — 8-10 см. Оси второго порядка появляются иногда из спящих почек на разных участках главной скелетной оси. Таким же образом образуются оси третьего порядка и в очень редких случаях — четвертого. Вследствие этого от многолетней распростертой оси в разные стороны змеевидно расползаются дочерние оси, благодаря чему куст приобретает вид округлой колючей подушки. Главные скелетные оси живут 70-90 лет, оси второго порядка — 40-50, третьего порядка —20-30 лет. На взрослых растениях отмирают скелетные оси второго - третьего порядков целиком, иногда и 1-2 главных. К концу основного цикла на длинных скелетных осях часто нет цветков, ежегодно развиваются одни вегетативные побеги. |
На стареющих скелетных осях, особенно активно в основании, из спящих почек в обилии образуются небольшие побеги дополнения. Они без цветков и недолговечны, боковых побегов, как правило, не образуют. После 2-10 лет нарастания эти побеги полностью отмирают. Скелетные оси не освобождаются от колючих сухих листьев и все почти до основания покрыты густым их покровом. Скелетные оси большей частью не укореняются, только в конце основного цикла в их основании могут образоваться 1-2 придаточных корня, но связь с материнской корневой системой не прерывается. Лишь у нескольких стареющих кустов мы наблюдали выгнившую материнскую корневую систему и отмирающие скелетные оси, из которых 1 или 2 продолжали жить за счет придаточной корневой системы.
В онтогенезе караганы гривастой четко выделяются следующие фазы: 1) розеточная (от 1 до 5 лет); 2) столбиковидная (от 5 до 10-15 лет); 3) системы первичного побега (от 10-15 до 25-30 лет); 4) собственно кустовидная — образование побегов формирования и СПФ (от 25-30 до неопределенно долгого времени, вероятно, не менее 100 лет). Для стланиковой формы прибавляется еще одна фаза: 5) партикуляция — образование придаточных корневых систем и обособление отдельных ветвей.
Трагаканты — Astragalus subgenus Tragacantae. По данным Т. П. Надёжиной (1962) и собственным наблюдениям в окр. Душанбе (1970) и Тбилиси (1971). Зонтиковидные, подушковидные, шаровидные или распластанные аэроксильные кустарнички, до 1 м, с колючими ветвями. Колючки образованы черешками листьев, остающимися на ветвях в течение 3-5 лет. Обитают на каменистых и щебнистых склонах и плато в составе растительности нагорных ксерофитов. Проростки розеточного типа, к концу первой вегетации достигают 2 см и состоят из 8-15 узлов. В дальнейшем первичный побег нарастает в течение всей жизни или верхушка его отмирает от действия различных повреждающих факторов. Отрастание боковых побегов из верхних частей приростов происходит на второй — пятый годы в зависимости от видовой принадлежности и условий местообитания. При лучших условиях оно более обильно. В начале развития ветвление, как правило, нерегулярное, повторяется через 2-3 года, затем становится ежегодным. К 7-8 годам основные разветвления, которые в дальнейшем станут главными скелетными осями, закладываются полностью. Далее происходит лишь нарастание ветвей и их акро- или мезотонное моноподиальное ветвление.
Первичная ось обычно сохраняет ортотропное направление роста, реже — косо наклоняется, так что боковые ветви отходят от нее вверх. В возрасте 8 лет растения достигают 7-10 см высоты. Примерно к десятому году интенсивность роста увеличивается, приросты достигают 3-5 (7) см. В это же время на приростах вначале главных разветвлений, а затем и второстепенных появляются соцветия (изредка с шестилетнего возраста). Междоузлия при этом несколько растягиваются, приросты теряют розеточный облик, однако у вершин приростов листья располагаются более тесно. С началом роста побега на прошлогоднем приросте, в средней его части распускаются цветки, собранные в пазухах листьев в короткие кистевидные соцветия из 2-5 цветков. Несколько выше их отрастают боковые побеги в числе 2-5. Большинство из них на второй - пятый год отмирают, проходя предварительно тот же цикл, что и прирост главного побега, но обычно без дальнейшего разветвления. Лишь некоторые из них становятся многолетними ветвями, образующими крону кустарника.
В среднем кусты (по структуре скелетных осей — аэроксильные кустарники или деревца с коротким главным стволом) трагакантов имеют около 0,5 м высоты, в некоторых случаях — до 1 м. Отмирание верхушек побегов и переход к симподиальному ветвлению происходит лишь вследствие их повреждения. Появление побегов из спящих почек в основании кустов возможно лишь при сильном повреждении всего куста. Очень редко наблюдается слабое укоренение боковых ветвей, лежащих на почве. В течение всей жизни особь трагакантов обычно (если не учитывать сильных повреждений) представляет собой систему первичного побега, в морфогенезе которой четко можно выделить лишь две фазы: фазу розетковидного побега, проходящую всего за 1-5 лет, и фазу системы первичного побега. Лишь факультативно в конце онтогенеза появляется фаза развития побегов из спящих почек.
Вереск — Calluna vulgaria (L.) Hill. По Р. А. Ротову (1960), И. Г. Серебрякову (1962) и собственным наблюдениям в Подмосковье и окр. Ленинграда (1968-1975). Первичный побег длительно нарастающий. Приросты его вначале 1-2 см. С первого - третьего года на нем силлептически возникают короткие боковые веточки, большинство которых опадает на третий - пятый год. Но некоторые из них на третий - четвертый год могут перейти к усиленному росту и формировать длинные плагиотропные побеги (турионы) до 1 м, переходящие через 3-4 года к восходящему и ортотропному росту. Затем они, как и главная ось, ветвятся моноподиально. На третий - седьмой годы на первичной и главных плагиотропных осях в пазухах листьев в верхней части приростов появляются цветки. Примерно к десятилетнему возрасту структура приростов становится такой: размеры их достигают 10 см, верхняя их треть (кроме самой верхушки) занята пазушными цветками, непосредственно над которыми мутовкой отрастает несколько сильных боковых ветвей, развивающихся далее по типу материнской ветви. Ниже цветоносной части располагается большое количество более мелких и менее долголетних (живущих от 2 до 7 лет) далее не ветвящихся побегов. Они, очевидно, и выполняют главным образом ассимиляционную функцию. Все боковые побеги развиваются силлептически.
Долговечные боковые ветви первого и последующих порядков переходят к цветению позже, чем материнские. При ортотропном росте число этих порядков ограничено 4-5. Однако обычно первичная ось полегает в основании и становится восходящей, чему способствуют более тенистые и влажные условия существования. В сухих же борах система первичного побега вереска долго остается ортотропной, деревцевидной, достигая свыше 0,5 м. Цветение здесь более обильно. Цветки часто располагаются у самых верхушек приростов однобокими кистями, и после цветения все соцветие вместе с главной осью отмирает, что приводит к переходу от моно- к симподиальному ветвлению. В пятнадцати - двадцатилетнем возрасте полегшие нижние части ветвей сильно оголяются, и на них возможно образование побегов дополнения, а при различного рода повреждениях (например, на опушках вследствие выпаса) — побегов формирования, вначале вегетативных, а затем — с обильными пазушными цветками, расположенными довольно далеко от верхушки. При полегании главная, первичная ось быстро теряется среди боковых, число порядков которых сильно увеличивается (примерно до 10), чему способствует образование придаточных корней в их основании. Укоренившиеся ветви достигают 1,5—2 м, кустарник в целом приобретает вид куртины. Однако в большинстве случаев до конца жизни вереск остается стержнекорневым растением, лишь в более тенистых и влажных условиях его придаточные корни столь усиливаются, что укоренившиеся ветви могут существовать самостоятельно в виде плагиотропных, почти не образующих цветков ветвей. Таким образом, в онтогенезе вереска можно выделить следующие фазы: 1) столбиковидного первичного побега с укороченными боковыми побегами (первые 3-5 лет); 2) развития удлиненных боковых побегов (нижние из которых могут быть ростовыми, длинными, плагиотропными). Далее следуют факультативные фазы: 3) полегания и укоренения главных скелетных осей, симподиального ветвления; 4) образования клона.

Рис. 40. Бересклет. Побег формирования и поникшая верхушка материнской CПФ
|
Бересклет бородавчатый — Evonymus verrucosa Scop. По данным И. Г. Серебрякова (1962) и собственным наблюдениям в подмосковных смешанных лесах (1970-1975). Геоксильный кустарник, до 2 м, из нескольких или многих тонких скелетных осей, частью лежащих на почве. Сеянец высотой 3-4 см с парой семядолей и розеткой из 2-3 пар листочков. В последующие годы происходит нарастание первичного побега. В первые 2-4 года приросты около 3 см, с вечнозелеными листьями, сосредоточенными у их верхушек. Семядоли и листья живут до 3 лет. С четвертого - пятого года интенсивность роста главной оси у сеянца возрастает, достигая к 10-15 годам кульминации — у некоторых экземпляров 15-18 см в год. Всего главная ось, образующая стволик, живет до 30 лет. С седьмого - десятого года начинается ветвление главной оси по акротонному типу. Боковые ветви сохраняют плагиотропное направление роста и ветвятся далее преимущественно в горизонтальной плоскости, так что создается мутовчатое строение кроны. Приросты боковых ветвей первого и последующих порядков весьма значительны, не уступают приросту главной ветви или даже превышают его. Цветки находятся но нескольку на длинных тонких цветоножках в нижней и средней части прироста. Цикл его развития — год. По мере роста кроны, увеличения ее массы слабая главная ось начинает постепенно пригибаться книзу, вначале только верхушкой, а затем на половину или более своей длины, что происходит на 15-20-й год. Одновременно от нижней части главной оси, у самого ее основания или несколько выше, из спящих почек отрастают 1-2 побега формирования, создающие свои СПФ по типу СПП. Сложные стволики (ССО), образующиеся в результате этого, небольшие, до 1,5 м. Одновременно с отрастанием новых побегов формирования в основании первичной оси ее ветви, лежащие на почве, укореняются и продолжают рост. Близ укорененных мест из спящих почек развиваются турионы, но более мелкие, чем идущие от нижней части главной оси. Их основные циклы более короткие, и они полегают на 10-15-й год. Ветви, составляющие верхушки систем побегов (СПП или СПФ), вследствие отрицательного геотропизма растущих стеблей обычно саблевидно приподняты (рис. 40). |
По мере роста верхушкой они полегают основанием, присыпаясь опадом и придавливаясь снегом. Бересклет представляет собой очень оригинальную жизненную форму: в центре куста находятся ортотропные скелетные оси — несколько более мощных ССО, составленных 2-3 порядками СПФ (при большем их количестве они полегают), затем несколько более тонких стволиков, образованных СПФ, далее к периферии куста несколько еще более тонких и низких стволиков, образованных также СПФ от укорененных ветвей, и, наконец, на самой окраине все далее расползающиеся от центра укореняющиеся ветви — СПИ. Таким образом, бересклет бородавчатый — и прямостоячий, и ветвеукореняющийся, и ползучий (стланиковый) кустарник в одно и то же время. При этом на более осветленных местах преобладают более высокорослые прямостоячие формы, в более затененных — ползучие. Некоторые виды, как например, бересклет европейский Evonymus europaea, образуют преимущественно прямостоячую, деревцевидную форму, большинство же — стланиковую (кавказский Е. lalifolius Mill., среднеазиатский Е. koopmannii, дальневосточные Е. macroptera Rupr., E. maackii), способную существовать только за счет плагиотропного разрастания СПБ. Это — типичные лесные стланики, но не облигатные, как линнея и барвинок, а факультативные. Плодоношение у них отсутствует или очень ослаблено. Подобные формы могут образовывать и многие другие лесные кустарники, как-то: липа, жимолости, калины (см. ниже), но у бересклетов они наиболее обычны. В онтогенезе бересклета бородавчатого четко выделяются две основные фазы: 1) образование системы главной скелетной оси (СПП) и 2) фаза клона. Первая фаза делится на следующие подфазы: а) рост первичной оси, б) образование СПП — первичной кроны, в) замедление роста первичной оси и ее полегание. Фаза кущения, четкая у большинства других геоксильных кустарников, как бы смазана и совпадает с началом образования клона.
Смородина душистая — Ribes fragrans Pall. Магаданская обл., Омсукчанский р-н. Каменистые осыпи до 1000 м над уровнем моря. 1973. Единичные кусты из множества разновозрастных скелетных осей, до 1,5 м. Прорастание надземное. Первичная ось дает 2—3 прироста от 2—3 до 5 мм, с 2—3 листочками. На третий - четвертый год от ее основания начинают отрастать 1-2 побега кущения (формирования). Они также имеют 2-3 прироста: первый — до 1,5 см, последующие — до 0,8 см. После отмирания их верхушки развивается второй порядок побегов кущения, уже более крупных и более долговечных, с первым приростом до 2 см и последующими 2-3 — около 1 см. Они уже ветвятся по моноподиальному типу, образуя СПФ. Боковые побеги укороченные, 0,1 см, с 2 листьями 1х1 мм, живут 1-2 года. На пятый - седьмой год первичный побег и побеги кущения первого порядка отмирают. Оставшиеся от них «пеньки» утолщаются. Здесь образуется узел кущения, откуда из почек, покоившихся 3-5 лет, ежегодно отрастают по 1-2 туриона, все более крупных, с более долговечными укороченными побегами. Начиная с третьего порядка, замирание верхушечной почки туриона не ведет к окончанию основного цикла, так как нарастание его оси в течение 3-4 лет продолжает ближайший к верхушке боковой побег продолжения. У турионов последующих порядков бывает по 2-3 побега продолжения.

Ribes triste
|
На 10-15-й год отрастают побеги кущения четвертого - шестого порядков из 3-4 приростов, первый из которых — до 10 см, последующие — 3-4 см (в целом - 20 см и более), с 5-7 листьями, достигающих максимальных в данных условиях размеров (3-4х4-5 см). В основании турионов появляются первые придаточные корни. В это время развиваются боковые побеги двух типов: непосредственно под верхушкой прироста главного побега, 1-2, удлиненные до 8 см, с 7-8 листьями и ниже их — укороченные. Примерно на 20-й год куст достигает 25-30 см в высоту и состоит из 10-15 побегов кущения, 4-5 из которых — скелетные оси, построенные из туриона и 2-3 порядков побегов продолжения. В середине куста они ортотропны, по краям — с полегшими на 4-6 см основаниями. Новые турионы появляются не только в зоне кущения, но и на этих полегших, часто засыпанных щебенкой частях осей, благодаря чему они превращаются в ССО. В это же время на укороченных побегах ветвления появляются первые кисти, пока еще короткие и малоцветковые.
К 20-30 годам вследствие усиленного роста кусты смородины достигают максимальной в данных условиях высоты — до 1,5 м и состоят более чем из 30 скелетных осей, основания которых могут полегать на 15-20 см и укореняться. Турион нарастает 3-5 лет. Первый его прирост 20-25 см, последующие — 10 см и менее. Затем его сменяют до 5 порядков побегов замещения, формируя главную ось. Удлиненные побеги ветвления нарастают 3-5 лет, их приросты до 10 см. На них, как и на главной оси, располагаются плодушки и свои удлиненные боковые побеги, и так до 4 порядков. Плодушки сидят почти по всей длине приростов, кроме нижней части. Кисти их до 6 см, с 10-15 цветками. |
Некоторые из них могут перейти со временем к нормальному росту и иметь пазушные кисти. Живут они до 10 лет. По море нарастания туриона и главной оси они, в конце концов, начиная снизу, отмирают, но долго еще остаются на ветви в виде многочисленных черных пеньков, придавая ей корявый вид. В течение основного цикла размеры побегов и количество приростов в них уменьшаются, к середине его начинают пробуждаться спящие почки, производя побеги дополнения. На 10-15-й год от основания СПФ отрастают новые турионы, образующие новые СПФ, равноценные предыдущим. Всего в этот период, от 20-30 до 60-80 лет, сменяется до 5 порядков СПФ с основными циклами, длящимися около 20 лет.
У растений в возрасте 80-100 лет приросты удлиненных побегов сокращаются до 2-3 см, нарастание туриона длится 5-6, а побегов ветвления — до 10 лет. Плодушки живут до 15 лет, однако они короткие, 2-3 см, соцветия с 2-5 цветками образуются редко. Уменьшаются в размерах и листья (1,5х2 см). На приростах их всего 3-4. СПФ утрачивает вид компактной ортотропной ветви. Активно образуются побеги дополнения с 2-3 порядками ветвления, в основном вегетативные. За счет длительного роста побегов, в особенности дополнения, основной цикл сильно растягивается, до 30 и более лет. Новые турионы под конец его отрастают в большом числе, но обычно не образуют боковых побегов и через 3-б лет отмирают. Выжившие турионы продолжают развитие по описанной схеме. Так как па смену 2-3 отмирающим СПФ приходится 1, редко 2 СПФ, куст, начиная с середины, редеет. Краевые стволики, полегающие в основании, более долговечны. Однако отдельные части куста не обособляются. Вслед за центральными, несмотря на обильное появление молодых, но недолговечных побегов формирования, деградируют и краевые СПФ и ССО. Полностью куст отмирает, по-видимому, в возрасте около 150 лет.
Таким образом, в онтогенезе смородины душистой можно выделить следующие основные фазы: 1) рост первичной оси (первые 3-4 года); 2) образование первых турионов (побегов кущения) и развитие все более крупных СПФ (до 10 лет); 3) продолжение роста туриона замещающими побегами, появление придаточных корней, продолжение развития все более крупных СПФ не только в зоне кущения, но и на полегших частях материнских побегов (до 15-20 лет); 4) достижение СПФ максимальных размеров в данных условиях существования, состояние равновесия между ростом и отмиранием (от 20-30 до 60-80 лет); 5) деградация куста — преобладание отмирания над нарастанием, сокращение живой биомассы, активный рост побегов из спящих почек (примерно до 100 лет); 6) то же, но с постепенным угасанием побегообразующей деятельности, заканчивающейся полным отмиранием.
Моноподиально-симподиальная группа
Лещина, орешник обыкновенный — Corylus avellana L. По И. Г. Серебрякову (1962) и собственным наблюдениям в Подмосковье, в лиственных и смешанных лесах. 1968-1970. Раскидистый высокий кустарник, до 3-4 м, с 2-5 главными стволиками. Прорастание подземное, прирост первого года до 20 см, второго — 10-15 см. В последующем нарастание может быть симподиальным. Побеги продолжения отрастают из самой верхней живой почки, формируя ложный побег. Примерно на пятый год приросты достигают максимума — 30-40 см. Боковые побеги, возникающие с третьего года, мелки и недолговечны. Затем интенсивность приростов падает и в 12-14 лет составляет 2-3 см, а в возрасте около 15 лет первичная ось несколько наклоняется, вовсе прекращает рост, а затем начинает отмирать. В период ослабления роста первичной оси боковые побеги усиливаются и она теряется среди них. Ветвление происходит в основном в горизонтальной плоскости.
В это же время, на шестой — десятый год, в основании первичного побега просыпаются спящие почки, из них отрастают турионы первого порядка с 2 первыми приростами 20-30 см. Приросты второго — четвертого годов могут быть крупнее — до 70 см, затем нарастание идет по симподиальному типу с побегами продолжения 20-30 см. С трех- четырехлетнего возраста начинается интенсивное акросимподиальное ветвление, также с постепенным падением приростов боковых побегов. При этом верхушка всей СПФ постепенно наклоняется и ветвление происходит главным образом в горизонтальной плоскости. Имея приросты одной величины с боковыми ветвями, главная ось вскоре теряется среди них и вовсе прекращает нарастать. СПФ к этому времени намного перерастает СПП. Последующие СПФ, развивающиеся на основе турионов второго порядка, еще более крупные и лишь СПФ третьего - пятого порядков достигают максимальных в данных условиях размеров, в среднем 2,5-3 м, развивая до 20 порядков побегов ветвления. Первые 2-3 из них (в нижней части туриона) выполняют еще ростовые функции, имея 25-30 см в длину, последующие — долгое время дают приросты около 10 см, заканчиваясь 1-3 сережками. Ниже них располагаются 1-3 почки с женскими цветками и только за ними — 3-4 вегетативные почки. Таким образом, генеративные побеги ветвления лещины, так же как берез и ольхи, в типе дициклические.
Побеги ветвления последних порядков слабые, вегетативные и недолговечные. К концу основного цикла, одновременно с возрастающим интенсивным отмиранием побегов, усиливается рост их из спящих почек. В основном это побеги дополнения. Однако в месте перегиба ветви, на многолетней части туриона, отрастают и настоящие побеги формирования, создающие второй ярус ветвей образованной таким образом ССО. Однако ярусность кроны у лещины выражена плохо, так как побеги формирования обычно возникают в кроне редко, уже в конце основного цикла, когда материнская СПФ уже сильно наклонена. Лишь на стволиках стареющих кустов они становятся более или менее обычными, напоминая «водяные побеги» деревьев.
Подземные части первых турионов имеют несколько восходящее основание, от которого со временем отходят придаточные корни. При отмирании СПФ они долго остаются живыми и в совокупности с такими же частями других турионов составляют ксилоподий. С возрастанием порядка побегов формирования увеличивается и восходящая, подземная их часть, достигая 20 см и превращаясь, таким образом, в подобие настоящего корневища с чешуевидными листьями, придаточными корнями и загнутой верхушкой. Так как дочерние приземные турионы отходят в направлении от центра куста, то куст разрастается по краю, а середина его со временем оголяется. С одной стороны, разрастание, развитие придаточных корневых систем, с другой — отмирание первых СПФ и ССО ведет к распадению единого куста на ряд частей, в которых находится больше отмирающих СПФ, чем развивающихся.
Таким образом, онтогенез куста лещины можно подразделить на следующие фазы: 1) рост первичного побега и развитие СПП (этап древовидного роста И. Г. Серебрякова), в котором, в свою очередь, выделяются этапы: а) усиленного роста, б) усиленного ветвления СПП, в) ее деградации ; 2) появление и развитие все более крупных СПФ вплоть до достижения ими максимального в данных условиях размера; 3) «стабилизация» — появление СПФ, одинаковых по размерам, но со все более длинной корневищной частью побегов формирования, развитие их также и в кроне; 4) партикуляция — распадение куста на несколько автономных, но менее жизнеспособных, чем первоначальный, кустов.
Степная вишня — Cerasus fruticosa (Pall.) Wronow. По данным Ю. Е. Алексеева (1959, 1963, 1974) и собственным наблюдениям в Московской и Курской обл. (1968-1970). Невысокий, до 1 м, кустарник, с многочисленными редко стоящими стволиками. Прорастание надземное. В первые 2-3 года приросты в 2-3 см, в последующие — около 10 см и более. Со второго - третьего года акротонно образуются боковые удлиненные побеги. Главная ось нарастает 4-7 лет, затем ее сменяет побег замещения, так что пятнадцати—двадцатилетний стволик оказывается построенным из главного и нескольких порядков замещающих побегов. В верхней части главного корня в четырех—десятилетнем возрасте закладываются придаточные почки. Позже они образуются на боковых горизонтальных корнях. Из придаточных корневых почек отрастают турионы как в основании главной оси в качестве побегов кущения, так и на горизонтальных корнях в качестве парциальных осей или побегов парцелляции. Первые их приросты большие, 15-20 см, на четвертый год они падают, в последующем — 5-10 см. Нарастание продолжается в течение 3—6 лет, затем рост стволика, образованного турионом, идет, как и у СПП, за счет побегов замещения. Одновременно с нарастанием побеги формирования и замещения акротонно ветвятся до шестого порядка. Удлиненные вегетативные побеги отрастают от верхних частей приростов, по всей длине которых располагаются плодушки разной степени специализации. Обычно они укороченные, все заканчиваются зонтиковидным соцветием (см. рис. 9, б), живут не более 2-3 лет. Начиная примерно с десятого года основного цикла, происходит развитие побегов дополнения в кроне последовательно на двух-трехлетних и более долговечных побегах. Однако системы, образуемые ими, недолговечны. Турионы от основания стволиков первичного побега или побега формирования предыдущего порядка отрастают редко, так что кусты состоят из небольшого числа (2-3) стволиков главных скелетных осей, которые в случае усиления побегов дополнения и перехода их в категорию ростовых могут быть и сложными. Всего стволики (СПФ) живут до 30 лет.
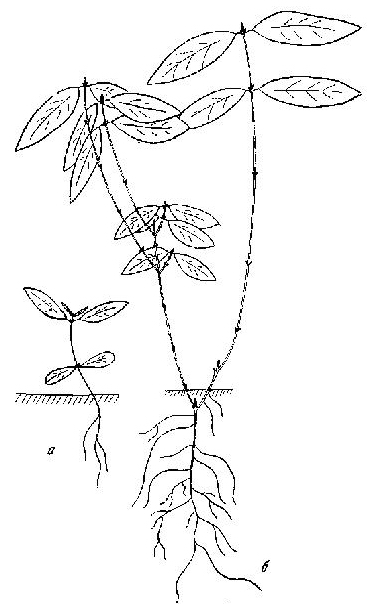
Рис. 41. Свидина белая
а — первый год развития; б — третий год развития
|
Корни, как и у терна, утолщаются только между побегами формирования (корневыми отпрысками) и растущим концом. Нетрудно видеть, что эти этапы соответствуют выделенным нами для основных циклов периодам: рост, стабилизация и отмирание. Участок корня, идущий к материнскому кусту, остается тонким. Зрелая особь вишни степной представляет собой материнский куст с отходящими от него несколькими горизонтальными корнями, которые достигают 5-7 м и несут ряд разновозрастных корневых отпрысков — побегов формирования. После отмирания материнского куста (в разных условиях в возрасте 12-30 лет) возникает клон из нескольких особей, каждая из которых состоит из горизонтального корня с системой разновозрастных побегов формирования придаточного происхождения. Такой корень биологически выполняет роль корневища, нарастая с одного конца и отмирая вместе с парциальными кустами с другого. Таким образом, можно выделить следующие фазы онтогенеза: 1) первичного побега; 2) системы первичного побега; 3) первичного куста (образованного, кроме СПП, 1-2 СПФ); 4) парциальных кустов (начало этой фазы накладывается на конец или даже середину предыдущей); 5) клона (начинающуюся с отмирания материнского куста).
Свидина белая — Thelycrania alba (L.) Pojark. Магаданская обл., среднее течение р. Колымы, пос. Сеймчан. 1975. Пойменные тополевники. Кустарник, с красными тонкими прутьевидными ветвями, до 2 м. Первичный побег нарастает в течение 2-3, а если проросток попадает в глубокую тень и во время весенних паводков полегает, то и большего числа лет, до 10. Ветвления при этом не происходит и стелющийся побег может достигать более 0,5 м, в случае отмирания верхушки замещаясь ближайшим к ней боковым. На осветленных местах сохраняется ортотропная форма роста. На второй-третий год акротонно появляется пара побегов ветвления, нарастающих 1-2 года по 5-15 см. После окончания их роста появляется еще по паре замещающих побегов. К третьему - четвертому году кустик достигает 30 см и имеет разветвления 2-3 порядков. Во время паводков основание его заносится илом и укореняется, но не полегает. На четвертый - пятый год под отмершей верхушкой прошлогоднего прироста появляются уже 2 пары замещающих побегов: нижняя слабая с приростами до 1 см, живущая 1-2 года, и верхняя более сильная, прутьевидная. Если эти побеги имеют 2 прироста, то они ветвятся моноподиально, давая слабые и недолговечные боковые веточки. При акросимподиальном ветвлении верхушки длинных хлыстовидных побегов не вызревают и их верхняя часть (5-10 см) отмирает. Под ними отрастает пара таких же длинных побегов продолжения. Один из них может лидировать (рис. 41). В течение последующих 4-5 лет происходит акросимподиальное ветвление простых и дициклических хлыстовидных побегов 10—20 см. Верхушки их, как правило, отмирают, у простых — в большей степени. К десятилетнему возрасту в 4-5 их верхних узлах появляются силлептически укороченные побеги, которые на следующий год дают терминальное щитковидное соцветие, под которым также силлептически отрастают 1-3 пары дициклических генеративных побегов ветвления следующего порядка, и т. д. Такой порядок ветвления характерен для наиболее сильных верхних побегов, возникающих под самым основанием соцветия из пазух ближайшей к нему пары листьев. Чем пазушный |
побег расположен ниже, тем он слабее и недолговечнее. Самые нижние отмирают на второй - третий год, не переходя к цветению. Нарастание системы, следовательно, продолжают главным образом верхние пары побегов. Размеры их многолетних частей вначале возрастают, а затем постепенно падают (табл. 3). Верхняя часть образованной таким образом системы побегов (ветвь) постепенно наклоняется. Первичная ось и основание первых двух ее разветвлений, ставших главными скелетными осями, заносится илом. Начиная с образования генеративных побегов ветвления седьмого — десятого порядков, все больше появляется ослабленных и недолговечных вегетативных побегов, начинают отмирать не только мелкие, по и более крупные ветки. Основания главных скелетных осей полегают на 30—40 см, и па них появляются первые турионы из почек, покоившихся не менее 10 лет.
Таблица 3
Средняя длина побегов с системе первичной оси Thelycrania alba
Возраст, годы |
Порядок ветвления |
Длина верхних боковых побегов, см |
Длина
нижних
боковых
побегов, см |
Возраст, годы |
Порядок ветвления |
Длина верхних боковых побегов,
см |
Длина нижних боковых побегов,
см |
2-3
4-5
6-7 |
1-3 3-5 5-6 |
7,7 10,5 33,5 |
2,7 3,1 |
7-10
10-15
15-20 |
6-8
8-10 10—15 |
16,1
12,4 2,4 |
1,5 1,5
- |
|
Система же первичного побега (СПП) живет всего около 20 лет. В ее развитии четко выделяются периоды: а) усиленного роста (побеги длинные, вегетативные), б) усиленного ветвления (побеги короткие, генеративные), в) деградации (побеги мелкие, вегетативные). Нарастание побега формирования может длиться до 4 лет, в течение которых он достигает более 1 м, заканчиваясь терминальным соцветием, ветвясь акротонно, моно- или симподиально. Во втором случае побеги продолжения до 0,5 м, ди- трициклические. Нижние боковые побеги более мелкие, вегетативные и недолговечные, они, не ветвясь, могут нарастать до 10 лет.
|
Развитие СПФ идет за счет верхних боковых побегов (табл. 3), силлептически отрастающих под соцветием предыдущего. Первый их прирост короткий, второй, генеративный — длинный. Иногда побеги ветвления моноциклические и не имеют укороченного прироста. В дальнейшем ростовые функции этих побегов ослабевают, они становятся всё короче. Таким образом, в течение 4-6 лет образуется 5-7 порядков побегов. Затем ослабевают и их генеративные функции, соответственно увеличивается длительность их нарастания — до 10 лет, основной цикл сильно растягивается — до 40-50 лет. В конце его вершина СПФ , достигшая 2 м, дуговидно изгибается. Начинают просыпаться спящие почки и производить побеги дополнения, 1-3 см, нарастающие обычно до 10 лет. Еще в середине основного цикла основание туриона полегает и на нем образуются несколько СПФ второго порядка, затем третьего и т. д. Таким образом, ССО у этого кустарника оказываются с самого начала простратными, лежачими, поэтому-то не образуется и ярусов в кроне.
Кроме того, могут укореняться не только полегающие многолетние турионы, но и верхушки наклонившихся СПФ. Поблизости от придаточных корней при этом вскоре появляются турионы, дающие начало новым СПФ и парциальным кустам. К усиленному ортотропному росту, превращаясь тем самым в турионы, могут перейти и сами укоренившиеся побеги ветвления. Дальнейший онтогенез идет за счет образования все новых парциальных СПФ и кустов. Таким образом, к выделенной выше фазе — развитие СПП — прибавляются следующие: образование первичного куста, СПФ которого, особенно первая, проходят примерно те же фазы, что и СПП, и образование парциальных кустов и клона. Вследствие того, что последующие системы развиваются тогда, когда предыдущие еще находятся в состоянии расцвета, а ветви всех систем сильно переплетаются между собой, фазы их деградации и усиленного роста накладываются друг на друга и тем самым взаимно «гасятся».
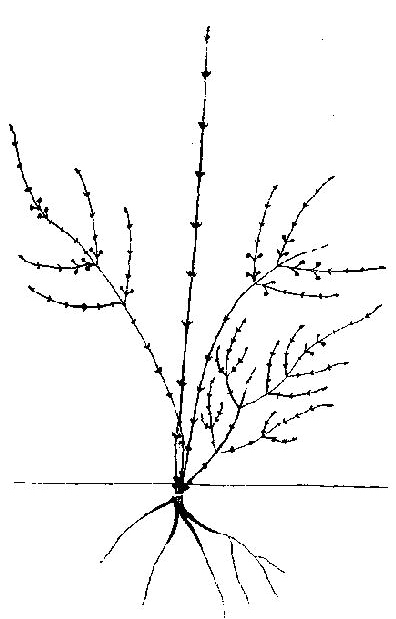
Рис. 42. Жимолость камчатская. Фаза усиленного роста
|
Жимолость камчатская — Lonicera kamtschatica (Sevast.) Pojark. Начальные фазы изучались в культуре, ГБС (Москва), последующие — в Камчатской обл., долина р. Камчатки, каменноберезники. 1969. Раскидистый кустарник с ярусным строением кроны. Первичная ось проростка нарастает до 3 лет, достигая 20 см и образуя короткие боковые веточки. Основание ее, как правило, коленчато изгибается. Уже со второго года на этой полегшей части возможно появление первого побега кущения, вырастающего за один период роста до 20 см, т. е. до размеров СПП. Далее, в течение примерно 10 лет, отрастают новые турионы, все более крупные, с побегами ветвления 2 порядков, часто уже с пятого года несущими двуцветники. Всего к десятилетнему возрасту отрастает 3-4 порядка СПФ до 30-40 см, последние из них — с горизонтально простертыми верхушками. Основной цикл их проходит за 2-3 года, после чего они, начиная сверху, отмирают. СПП отмирает на четвертый - пятый год почти до основания, кроме маленького пенька, служащего узлом кущения (рис. 42). Примерно с десятого года новые турионы начинают появляться не только в узле кущения, но и в кроне, в местах перегиба побегов формирования, достигающих 1 м. СПФ в это время состоит из 3 порядков побегов ветвления от 10-15 до 2-3 см. Таким образом начинают формироваться главные составные скелетные оси . Однако последующие турионы некоторое время продолжают перерастать предыдущие, увеличивается и число ярусов СПФ. Вeличина побегов, число их в СПФ и ССО достигает предельных величин примерно к двадцатилетнему возрасту. Но еще некоторое время образование новых побегов и их систем явно преобладает над отмиранием, что ведет к увеличению живой биомассы куста. Лишь в 25-30 лет наступает более или менее полное равновесие между нарастающей и отмирающей массой побегов. Основной цикл в это время наиболее типичен. Турионы состоят из 2-3, редко — более приростов, первый из которых, наибольший, около 1 м, а последующие — такой же величины, как и побеги ветвления соответствующего порядка: 40-50, 15-20, 10-15 см. Ветвление происходит в горизонтальной плоскости, отчего и зависит ярусное строение кроны.
Постепенно, однако, несмотря на регулярное появление побегов дополнения, число отмирающих побегов начинает превышать число нарастающих. ССО, расположенные в центре куста, полностью отмирают. Краевые ССО дугообразно наклоняются, касаясь почвы верхушечными ветками, которые укореняются. Близ мест укоренения отрастают новые турионы, что ведет к образованию парциальных кустов, а затем в возрасте около 50 лет клоны в условиях, связанных с затенением, в камчатских ельниках возникающие у ювенильных растений побеги кущения (5-6 см) на второй - третий год полегают на почву, имея всего 1 порядок ветвей. Первое время укоренения полегающих турионов не происходит. СПФ отмирают почти до основания. Однако последующие побеги кущения превышают предыдущие. С увеличением размеров СПФ увеличиваются и остающиеся живыми части турионов, на которых отрастают придаточные корни и новые турионы уже на некотором расстоянии от первичной корневой системы. Эта стадия развития стланиковой формы соответствует периоду образования ССО ортотропных форм (рис.43). Отмирание первичной корневой системы приводит к образованию клона (рис. 44). |

Рис. 43. Жимолость камчатская, полупростратная форма
|

Рис. 44. Жимолость камчатская, простратная форма
|

Lonicera kamtschatica
|
Размеры побегов, число порядков их в СПФ и ССО, длительность основного цикла всецело зависят от условий существования. В более благоприятных, светлых местах они больше, в менее благоприятных — меньше, т. е. меньше размеры ортотропных частей, длительность основного цикла, быстрее происходит полегание СПФ, ветвь в целом имеет более «стланиковый» вид. Но длительность роста побегов может увеличиться. Отдельные побеги формирования и ветвления у полегших СПФ имеют восходящее направление роста, и, как и у бересклетов, нарастая и полегая верхушкой, могут очень долго пребывать в таком «ползучем» состоянии, образуя СПВ. Фазы онтогенеза ортотропной формы: 1) развитие СПП , 2) появление турионов и все более усиливающийся рост СПФ, 3) стабилизация — равновесие между ростом и отмиранием, 4) деградация первичного куста, 5) ветвеукоренение, образование клона. У простратной формы все фазы протекают очень быстро, четвертая — вовсе не выражена, но зато последняя очень растянута.
Курильский чай — Dasiphora fruticosa (L.) Rydb. По данным Шафрановой (1964, 1970) и собственным наблюдениям на юге Магаданской области, в сухих лиственничниках (1971-1974). Кустарник, из нескольких разновозрастных осей, до 1 м. Приросты первичной оси 0,5-1 см, на третийчетвертый год — до 2 см, с листьями, ясно сближенными у верхушки. Ветвления не происходит. На третий - шестой годы основание первичной оси на протяжении до 2 см полегает, и на нем трогаются в рост спящие почки в основании первичной оси, образуя побеги кущения (формирования) первого порядка с более интенсивными приростами. Первичная ось нарастает примерно до 15 лет, после чего производит несколько ветвей, на 20-25-й годы СПП полностью отмирает. Турионы |
проходят тот же цикл, что и первичная ось, однако приросты их длиннее. Образуются турионы все более высокого
порядка, все более крупные и долговечные. К 30-40 годам, когда появляются первые цветки, они достигают полной величины (около 1 м высотой). Основания их с придаточными корнями, более долговечны, чем СПФ, и из них формируется подземная многолетняя деревянистая часть куста — ксилоподий с более сильным стержневым корнем. От нее отходят многочисленные разновозрастные скелетные оси, проходящие свой цикл развития за 20-25 лет. Сильно разросшиеся кусты со временем партикулируют. Стержневой корень у них отмирает, и куст распадается на несколько частей по числу наиболее мощных придаточных корней. Отдельные его части благодаря более интенсивному развитию побегов с несколько восходящими основаниями выступают вперед.
В Забайкалье у кустов, достигших двадцатилетнего возраста и состоящих из нескольких скелетных осей, начинают развиваться подземные корневищеобразные стебли, вначале короткие, 3-5 см, затем все более длинные и более длительно нарастающие под землей. Выходя на поверхность, они производят длинный ортотропный прирост, становясь турионами. От их основания отрастают 1-2 порядка дочерних побегов формирования, образуя парциальный куст. Побеговая система первичного и парциальных кустов живет около 30 лет, их корневые системы намного дольше. Однако и конце концов они также отмирают, в результате чего куртина, образованная парциальными кустами, превращается в клон.
На юге Магаданской области цикл развития СПФ происходит следующим образом. Первый прирост туриона, 15-20 см, часто заканчивается терминальным цветком. В пазухах его несколько сближенных верхних листьев, ближайших к терминальному цветку, силлептически развиваются укороченные побеги, которые также могут заканчиваться цветками и в этом случае эфемерны. Второй прирост, если он бывает, 10-15 см, имеет такой же вид, что и первый. В его отсутствие на следующий год укороченные побеги ветвления дают длинные, до 15 см, приросты того же строения. Третий порядок ветвей отрастает от ветвей второго порядка почти по всей их длине, а четвертый — часто от самого основания ветвей третьего порядка, так что создается впечатление расположения в одной пазухе листа пучка из 2-3 веточек. Начиная с четвертого - пятого порядков, все больше появляется укороченных побегов, не переходящих к нормальному росту, или систем побегов, дающих попеременно то удлиненные, то укороченные приросты или побеги продолжения. Очень часто турион под тяжестью ветвей СПФ полегает в основании и на нем развивается следующий порядок турионов, проходящих тот же цикл. Чем суровее условия, тем полегание побегов выражено резче, тем меньше длина приростов и больше количество укороченных побегов, в результате чего возникают настоящие стержнекорневые стланики. Придаточные корни появляются только в основании крупных ветвей. Факты партикуляции или тем более отрастания корневищ установить не удалось. Фазы онтогенеза: 1) первичной оси и СПП; 2) первичного куста: а) усиленного роста последовательных СПФ, б) стабилизации, в) деградации; 3) факультативная — системы парциальных кустов и клона.
Куропаточья трава — Dryas grandis Juz. Якутская АССР, Момский р-н, окр. оз. Дарпир. На приречных галечниках. 1974. Стелющийся вечнозеленый кустарничек, с крупными белыми цветками и ветвями, одетыми плотным покровом из оснований отмерших листьев. Проростки розеточного типа. Последующие приросты первичной оси 0,5-1 см, также розеточные. С третьего - четвертого года появляются боковые побеги, вскоре ложащиеся на почву и дающие более длинные приросты, 1,5-2,5 см, с листьями, ясно сосредоточенными в плотную розетку у верхушки. Первичная ось нарастает 5-10 лет, достигая 8-12 см, завершаясь иногда терминальным цветком, образуя ежегодно 2-3 боковых побега, верхние из которых отмирают вместе с ее верхушкой. Нижние боковые ветви с полегающим основанием имеют восходящее направление роста. Ортотропная их часть до 5 см, состоит из 3-5 розеточных приростов. В то время как боковые побеги первого порядка переходят к ортотропному росту, они дают пазушные боковые побеги второго порядка с длинной, 5-10 см, нерозеточной частью и ортотропной розеткой. Последующие их 2-3 прироста производят менее длинные плагиотропные и короткие ортотропные боковые побеги. Подобный тип ветвления, кроме дриад, очень характерен для некоторых вересковых: Rhododendron schlippenbachii Maxim., Menziesia pentandra Maxim., Pieris japonica (Thunb.) D. Don. На пятый - десятый год роста боковые ветви начинают укореняться в основании. Однако система стержневого корня сохраняется еще долго, в течение не менее 10-15 лет после отмирания главной оси, после чего начинается фаза клона — автономное существование отдельных ветвей. Каждый из боковых побегов первого порядка через 2-5 лет нарастания, как и первичная ось, завершается крупным
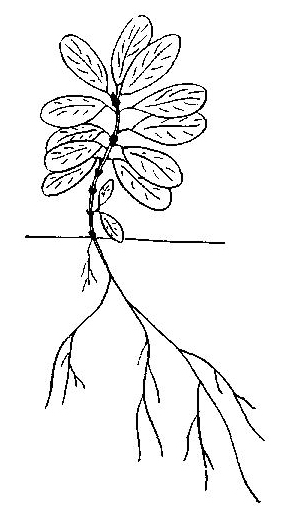
Рис. 45. Рододендрон золотистый
|
терминальным цветком на длинной цветоножке. Уже к концу цветения из пазух верхних листьев верхушечной розетки начинают отрастать 2-3 замещающих побега, один из которых может лидировать. В более благоприятных условиях побеги дициклические, в менее — полициклические. Остатки листьев держатся на них 3-5 лет. Через столько же времени на них могут образоваться придаточные корни, живущие, повидимому, не менее 10-15 лет, после чего прирост, на котором они сидят, отмирает и постепенно отгнивает. Приросты у взрослых растений в зависимости от условий 2-5 см. Постоянное лидирование одного из замещающих побегов может привести к формированию главных скелетных осей, боковые ветви которых недолговечны. На многолетних их частях в случае повреждений могут возникать побеги дополнения. Таким образом, в течение всего онтогенеза дриада представлена, собственно, системой первичного побега (в чем она сходна с трагакантами). Его основные фазы следующие: 1) стержневая, 2) клоновая. Первую можно подразделить на подфазы: а) существования первичной прямостоячей оси розеточного типа, хотя и с базитонным усилением лежащих на почве ветвей; б) стланика с восходящими ветвями и придаточными корнями на них.
Рододендрон золотистый — Rhododendron aureum Georgi . Окр. Магадана. Лиственничник с кедровым стлаником, багульником и другими вечнозелеными кустарниками. 1971-1975. Вечнозеленый стланик с приподнимающимися до 0,5 м ветвями. Проросток до 1,5 см с 3-4 листьями 2X4 мм (рис. 45). Нарастание первичной оси длится 5-6, иногда до 10 лет, приросты около 0,5 см со сближенными у верхушки листьями. Боковые побеги образуются начиная с третьего-четвертого года. С пяти - семи лет приросты первичной оси начинают заметно снижаться, а боковых — увеличиваться. Основание проростка изгибается, 2-3 первых прироста первичной оси оказываются лежащими на почве. От этой части в случае малой интенсивности ветвления могут отрастать побеги из спящих почек, первые приросты которых 2-4 мм, а последующие — до 15 мм. Таким образом, они представляют собой своеобразные турионы, перерастающие вскоре первичную ось. К 10 годам нарастание ее заканчивается, однако около верхушки развиваются 1-2 замещающих побега, продолжающие нарастание системы первичной оси. Один из них лидирует и вместе с первичной осью формирует ложный побег. Турионы, выполняющие в данном случае функцию кущения, нарастают до 10-15 лет, изредка моноподиально ветвясь. Вскоре они полегают основанием, так что ортотропными остаются лишь шестилетний проросток, который может привести к формированию главных скелетных осей, боко вые ветви которых недолговечны. На многолетних их частях в случае повреждений могут возникать побеги дополнения. |
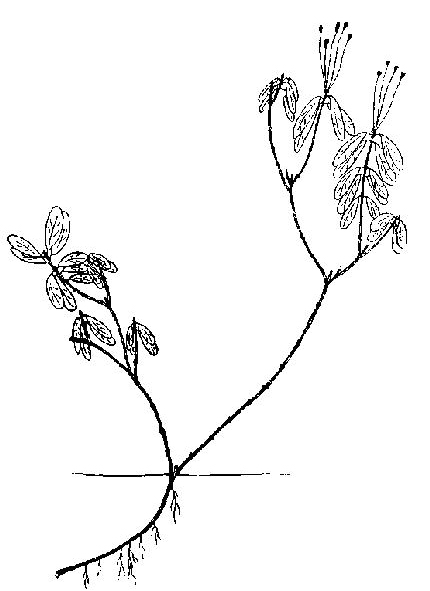
Рис. 46. Рододендрон золотистый. Восходящая форма роста
|
На пятом - шестом году от начала роста от основания побегов кущения первого порядка и от лежащей части первичной оси отрастает второй порядок или генерация турионов, также полегающих основаниями. Нарастают они более длительно (хотя более слабые отмирают на третий — пятый год). После цветения образуются замещающие побеги, продолжащие рост ветви, в основании которой появляются придаточные корни. К 20-25 годам укорененная плагиотропная часть побега кущения становится многолетней скелетной осью. Нарастание некоторых побегов длится до 20 лет. Приросты их 0,5-0,7 см, листья — 3,5х1,5 см. Иногда появляются побеги ветвления 0,3-0,5 см, живущие всего 2-4 года. В местах укоренения из почек, покоившихся 5-7 лет, иногда появляются саблевидно приподнимающиеся, затем полегающие основанием и укореняющиеся побеги кущения третьего порядка. В основном нарастание идет за счет побегов продолжения, которые нарастают 5-8 лет, вновь замещаясь побегами следующего порядка. К 30-35 годам связь с первичной корневой системой не утрачивается, но ослабевает. Куст имеет чашевидную форму, с оголенной серединой, до 10 см в высоту. Побегов кущения больше не образуется. Одновременно с разрастанием и укоренением приросты увеличиваются. Соцветия и листья достигают максимальных размеров 6х3 см. Число цветков в зонтиках повышается до 3-4. Под соцветием отрастают 2-4 замещающих побега. Ближайшие к верхушке побеги развиты сильнее, 3-4-циклические приросты их до 3 см, ослабленные побеги спустя 2-3 года отмирают, но затененные побеги могут медленно, по 5-7 мм в год, нарастать в течение 10-15 лет (рис. 46). Появляющиеся на многолетних частях ветвей побеги дополнения укороченные, недолговечные.
Разрастаясь в стороны, куртина, образуемая разными ветвями, увеличивается в диаметре, и ее живые краевые олиствленные части постепенно отодвигаются от первичной корневой системы. К 50-60 годам, когда длина ветвей достигает 20-25 см, связь с ней, вследствие отмирания первых приростов побегов кущения, прерывается и растение распадается в зависимости от числа укорененных ветвей на 3-5 парциальных кустов. Вскоре после этого давно полегшие побеги кущения отмирают полностью и побеговая система всей куртины тем самым из СПФ превращается в СПВ. Ко времени отрастания восьмого — десятого порядков побегов ветвления размеры ветвей достигают максимума. Устанавливаются постоянные в данных условиях существования размеры живой плагиотропной и ортотропной частей СПВ. В лучших условиях эта последняя выше, долговечнее и разветвленнее, достигая в каменноберезниках 30-50 см. Состоит она из 2-5 порядков побегов и имеет возраст от 10 до 20 лет (время от образования прироста на вершине ветви до его полегания и укоренения — цикл СПВ). Плагиотропная, стелющаяся живая часть не менее 1 м, погружена в почву и опад и по всей длине укоренена. Отрастающие от старых, десяти - двадцатилетних приростов побеги дополнения обычно недолговечны, но некоторые из них все же набирают силу, укореняются и создают свои системы — вначале СПД, а затем, по отмирании родоначального побега дополнения, превращающиеся в СПВ. |
Развитие побегов дополнения стимулируется в неблагоприятных условиях, на щебнистых склонах в высокогорьях. Приросты здесь максимум до 1 см, надземные части СПВ составлены 1-3 порядками побегов, цикл СПВ — 10-15 лет. Активно возникающие побеги дополнения недолговечны и быстро сменяют друг друга. Лишь в случае отмирания верхушки главной ветви, что бывает нередко, они выполняют замещающую функцию. Таким образом, в онтогенезе рододендрона золотистого можно выделить следующие основные фазы: 1) образование первичной оси и СПП; 2) кущение и первичный куст, состоящий из одной - нескольких СПФ; 3) превращение путем отмирания первичной корневой системы и формирования придаточных корней СПФ в несколько автономных СПВ. В экстремальных условиях существования добавляется фаза активного развития побегов дополнения и СПД.
Читать далее: окончание главы "Изменение систем побегов в онтогенезе"
|

