Структура и динамика экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2011. – 128 с.
Предисловие C.6-8
Беликович А.В., Василенко Н.А., Галанин А.В., Галанина И.А., Долгалева Л.М., Козырь И.В., Маврин И.Б., Роенко Е.Н., Яковченко Л.С. Динамика растительного покрова (результаты мониторинга на постоянных пробных площадях на юге Дальнего Востока и в Забайкалье С.9-85
Беликович А.В., Галанин А.В., Галанина И.А., Долгалева Л.М. Высотная дифференциация растительности Сохондинского биосферного заповедника С.86-129
Галанина И.А. К вопросу о скорости роста лишайников эпилитов на юге Приморского края С.130-143
Киселёва А.Г. Сохранение биоразнообразия сосудистых растений морских побережий Приморского края С. 144-156
|
УДК 581.14
К вопросу о скорости роста лишайников эпилитов на юге Приморского края
Галанина И.А.
Ботанический сад-институт ДВО РАН, г. Владивосток
On the question of epylithic lichens growth rate in the southern Primorskii Krai
Galanina I.A.
Botanic Garden Institute of the Far-Eastern Branch of the Russian Academy of Science (BGI FEB RAS), Vladivostok
Ключевые слова: лишайники, эпилиты, скорость роста, Приморский край, размерная структура ценопопуляций.
The author determined growth rate of three lichen species on the roofing slate of single-storey house situated in the village of Zarubino, south of Primorskii Krai. The roof was builtin 1954, and was replace din 2004 forre pair. In 2004, roofing slate sheets of northern exposition, angle 30 o , were covered by lichens for 70–80%. The most abundant were three species: Caloplaca sp.1, Xanthoria sp. and Physconia detersa. The author measured all lichen individuals on the sheets along the transects. In order to analyze size structure of lichen populations the author designed variational series where all measured individuals are ranged by decreasing their maximal diameter. The form of variational series depends on the ratio between large, middle, and small individuals. Variatonal series are approximated by curves with equations. Three quations of logarithmic trend sallow to characterize, compare and analyze sets of lichens-indicators. The research helped to determine growth rate of three lichen species in the southern Primorskii Krai: Caloplaca sp.1 – 1-1.3 mm per year, Xanthoria sp. – 0.7-1 mm, and Physconia detersa – 0.4–0.6 mm per year. It is necessary to notice very fast and effective lichen settlement on roofing slate comparatively with fresh rock chips.
Введение
Лишайники – это симбиотические системы, состоящие из трех компонентов: гриба, синезеленой водоросли и азотфиксирующей бактерии. Морфологические особенности лишайников в основном определяются генотипом грибного компонента. В теле лишайника водоросль является продуцентом, а гриб редуцентом органического вещества (Определитель лишайников ..., 1975). Лишайники в процессе жизнедеятельности преобразуют абиотическую среду, изменяют ее свойства и увеличивают разнообразие экологических ниш. Появление в процессе эволюции жизни на Земле лишайников является результатом одного из «экспериментов выхода жизни» на сушу несколько миллиардов лет назад (А.А. Галанин, 2002).
Размножение лишайников осуществляется несколькими способами: путем переноса обломков слоевища на другие места, где они затем приживаются и разрастаются; лишайники могут размножаться и специальными соредиями и изидиями. Соредии представляют собой одну или несколько клеток водоросли, тесно оплетенных грибными гифами, соредии часто образуют компактные скопления – соралии. Изидии – это простые или разветвленные выросты, покрытые снаружи коркой из грибных гиф, внутри содержащие водоросли. Изидии легко обламываются и так же, как соредии, переносятся ветром на новые места, где и дают начало новым системам – симбионтам (Мартин, 1967). Кроме этого, лишайники размножаются спорами. Споры образуются в плодовых телах гриба. С помощью спор размножается только грибной компонент. В этом случае новая особь лишайника развивается только в том случае, если спора гриба встречает на месте своего прорастания с соответствующей одноклеточной водорослью. Иногда лишайниковые водоросли, свойственными данному виду лишайника, проникают в плодовые тела на ранних стадиях, там размножаются и рассеиваются вместе со спорами гриба.
Скорости роста разных видов лишайников существенно различаются. Относительно быстрорастущими являются некоторые тропические кустистые виды, ежегодный линейный прирост которых достигает более 10 см. Лишайники, живущие в Арктике и высокогорьях, в год прирастают на доли миллиметра, в связи с чем некоторые из них живут до 10000 лет. Медленнее всего растут тонкие накипные циркумарктические виды, которые наиболее часто используются в лихенометрии, например роды Rhizocarpon, Aspicilia, Lecanora и др. По имеющимся данным, возраст некоторых накипных лишайников, обитающих в Альпах, достигает 600–1300 лет, в Гренландии – 4500 лет, а некоторых видов, которые произрастают в Норвегии, Колорадо и Антарктиде, – даже 10000 лет (А.А. Галанин, 2002). Скорости роста особей одних и тех же видов лишайников зависят от конкретных физико-географических условий и возраста самих особей. В целом, для достижения равновесия в сообществе медленно растущих эпилитных лишайников на плакорных поверхностях в Арктике и в альпийском поясе гор необходимо от 4 до 10 тыс. лет. Жизнедеятельный для эпилитных лишайников поверхностный слой большинства каменных морфоскульптур за этот период, как правило, разрушается и претерпевает обновление в результате воздействия рельефообразующих процессов. Кроме того, в течение этого периода происходят изменения климата, а также катастрофические природные и техногенные процессы, замедляющие и прерывающие ход сукцессионного развития. На большинстве плакорных поверхностей предельный рост накипных лишайников и характер конечной сукцессионной стадии ограничены, главным образом, степенью денудационной устойчивости поверхности и временным периодом цикла полного возобновления каменной поверхности в местных физико-климатических условиях (А.А. Галанин, 2002).
Определение скорости роста лишайников является актуальной, но недостаточно изученной проблемой лихенологии. Использование лишайников для датировки дневных поверхностей и в качестве индикаторов для оценки атмосферного загрязнения затрудняет именно незнание скорости и особенностей их роста. Натурные наблюдения показывают, что каменные поверхности, не имевшие контакта с атмосферой, будучи вынесенными на дневную поверхность длительное время не заселяются лишайниками. Так, на отвалах кассетеритовых шахт в Сохондинском заповеднике (Хентей-Чикойское нагорье) спустя 50 лет на свежих сколах горной породы, извлеченной из недр, лишайники еще даже не поселились. Только поверхности находившиеся в контакте с атмосферой порядка 100 лет могут служить субстратом для, казалось бы, столь неприхотливых живых организмов.
Существуют различные методы, определения скорость роста лишайника. Можно ежегодно измерять радиус (максимальный поперечник) слоевища, можно периодически измерять площадь особи накипного или листоватого лишайника, можно пронаблюдать рост путем проведение фотосъемки участка, покрытого лишайниками, через определенные интервалы времени. Иногда бывает полезно воспользоваться и косвенными методами. Сложность заключается в том, что поверхность, на которой определяется скорость роста лишайников эпилитов, должна быть точно датирована, должно быть точно известно, сколько лет поверхность доступна для поселения лишайников. Лишайники часто поселяются на оградах, крышах и стенах многих старинных зданий, на опорах мостов, могильных камнях и памятниках. Зная дату постройки этих объектов и определив размеры слоевищ, можно узнать скорость роста каждого вида. Возраст лишайников-эпифитов часто определяют по годичным кольцам стволов и ветвей дерева-хозяина, на которых эти лишайники обитают.
Зная скорость роста и размеры видов лишайников в определенных условиях, можно определяют возраст поверхности, на которой они растут. Этот метод назвается лихенометрией ( Beschel ,1958). Сравнивая фотографии статуй на острове Пасхи, сделанные с промежутком в 47 лет, заметили, что лишайники, покрывающие их, за это время выросли. Оказалось, что возраст лишайников на поверхности статуй варьирует от 380 до 850 лет, а это значит, что и статуям не меньше 800 лет. С помощью лишайника Rizocarpon geographicum был определен возраст ледниковых морен в Канаде и Гренландии, составляющий более 1000 лет. Виды лишайников, используемые в лихенометрии должны удовлетворять следующим требованиям. Слоевище их должно быть четко очерченным и близким к кругу; это должен быть легко распознаваемый визуально вид; данный лишайник должен иметь большую продолжительность жизни (Исаенко: http://www.crimea.edu/internet/Education/geoecology/24.htm ). Скорость роста у разных видов лишайников сильно отличается. Например, особи Umbillicaria sp. за год нарастают на 0,01–0,04 мм, а Peltigera sp. – на 25–27 мм. Есть данные о годовом приросте пельтигеры окаймленной – до 45 мм в год.
О скорости роста лишайников эпилитов на юге Приморского края пока данных нет. Поэтому целью настоящей статьи является определение скорости роста трех видов накипных лишайников на Зарубинском полуострове в Хасанском районе на побережье Японского моря.
Материалы и методы анализа размерной структуры ценопопуляций накипных лишайников
Определить скорость роста трех видов лишайников эпилитов оказалось возможно на шифере крыши одноэтажного дома в п. Зарубино, которую начали ремонтировать в 2004 г. Кровля была построена в 1954 г. К 2004 г. листы шифера на поверхности кровли, имеющей северную экспозицию с уклоном 30 градусов были покрыты лишайниками на 70–80 % поверхности. Самыми массовыми оказались три вида: Caloplaca sp.1, Xanthoria sp. и Physconia detersa. Измерялись поперечники особей штангенциркулем, при этом у каждой особи измерялся масимальный поперецник. Измерялись все особи подряд вдоль трансектов, заложенных вдоль листов шифера.
Для анализа размерной структуры ценопопуляций были построены вариационные ряды, в которых все измеренные особи выстраиваются в ряд по мере убывания их максимального поперечного размера, каждая особь в ряду изображена в виде столбца диаграммы, высота которого соответствует размеру данной особи, ее максимальному поперечнику. Чем больше измерено особей в ценопопуляции, тем длиннее получается вариационный ряд. Форма вариационного ряда зависит от соотношеня в ценопопуляции крупных, средних и мелких особей. Вариационные ряды аппроксимируются кривыми линиями, для которых могут быть подобраны уравнения. Уравнения логарифмических трендов вариационных рядов особей, расположенных по мере убывания размера особи, позволяют характеризовать, сравнивать и анализировать выборки особей лишайников-индикаторов.
Использование вариационных рядов для определения скорости роста предпочтительнее использования только максимальных по размерам особей, так как наиболее крупные талломы не несут полной информации о системе и не могут быть использованы для надежной датировки поверхности. Максимальная особь иногда может оказаться старше датируемой поверхности, иногда за одну особь могут быть приняты две слившиеся в результате роста. Также не могут дать полной информации о возрасте каменной поверхности и особи со средними размерами таллома, так как среднее арифметическое значение может не совпадать с модальным, и оценка возраста в этом случае всегда будет занижена (А.А. Галанин, 1992).
Для лихенометрических оценок Дж. Метьюс ( Matthews , 1975) вывел уравнение, использующее логарифмическую функцию. Выполненный А.А. Галаниным анализ данной зависимости для некоторых видов лишайников эпилитов в Магаданской области показал, что она отражает представления Р.Е. Бешеля о фазе быстрого нелинейного роста и, следующей за ней линейной фазе. Однако по данной функции в интервале возрастов более 100 лет, она принимает форму, близкую к линейной (А.А. Галанин, 1996; 2002). Среди известных лихенометрических методик она является наиболее удовлетворительной, позволяя рассчитать ошибки возрастных оценок.
Для субарктической тундры побережья Анадырского лимана была определена скорость роста лишайников: секция Rhizocarpon – 0,14±0,01 мм/год, род Aspicilia – 0,15±0,01 мм/год. В лесотундровой зоне центральной части Корякского нагорья для секции Rhizocarpon – 0,18±0,03 мм/год, а начальный годовой прирост листоватого лишайника Parmelia sp. – 0,47±0,08 мм/год. В Примагаданье (зона северной тайги) параметры роста лишайников секции Rhizocarpon составляют 0,23±0,03 мм/год. Таким образом, скорости роста исследуемых лишайников в разных физико-географических районах существенно различаются (А.А. Галанин, 2002).

Лишайники на старой шиферной крыше.
|

Старые крыши, крытые шифером, в условиях муссонного климата южного Приморья за 50 лет почти полностью покрываются накипными и чешуйчатыми лишайниками.
Фото с сайта: http://images.google.ru/images?q
|

Яркожелтая ксантория - Xanthoria sp. (лопастная) и коричневыя калоплака - Caloplaca sp.1 (накипная) на старом шифере. Время обростания около 50 лет
|

Яркожелтая ксантория - Xanthoria sp. (лопастная) и коричневыя калоплака - Caloplaca sp.1 (накипная) на старом шифере. Время обростания около 50 лет
|

Коричневая калоплака - Caloplaca sp.1 (накипная) и серая фискония -
Physconia detersa на старом шифере. Время обростания около 50 лет.
Лишайники на шифере, возраст около 50 лет.
|

Фискония стёртая -
Physconia detersa на старом шифере. Время обростания около 50 лет.
|
Папа, фото выбрала что получше .
Слушай, я бы все же поставила такие названия, после просмотра фото:
Caloplaca sp. 1 накипная - так и есть
А вот Caloplaca sp.2 - это все же Xanthoria sp.
И Physconia detersa - лучше написать Physconia sp. - все же полноценно я не смотрела образцы
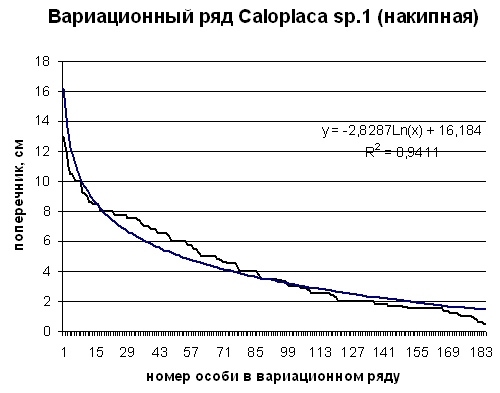
Рис. 2. Вариационный ряд особей Caloplaca sp.1 (накипная) ценопопуляции на шифере крыши на юге Приморского края (черная ломаная линия). Гладкая синяя линия – логарифмическая аппроксимация вариационного ряда. Пояснения в тексте
|
Результаты и обсуждение
Выборка особей накипного лишайника Caloplaca sp.1 (накипная) насчитывает 183 особи. При измерении учитывались все особи подряд, что позволило получить вполне репрезентативную выборку. Вариационный ряд этого вида приведен в виде диаграммы (Рис 2). Вариационный ряд аппроксимирует логарифмическая кривая. Точность аппроксимации оценивает коэффициент R =0,9411. Логарифмическая функция часто используется для описания роста живых систем – организмов и ценопопуляций. Отклонения от логарифмической зависимости в росте живых систем обычно бывают связаны с тем, что в реальных системах имеет место конкуренция между особями, которая осложняется в многокомпонентных системах, где в конкурентные отношения вступают особи разных видов.
Аппроксимирующая логарифмическая кривая описывается уравнением, которое приведено на диаграмме. Здесь а = -2,8287; b = 16,184, коэффициент b – это точка пересечения кривой с осью ординат. Эта величина есть рассчитанное максимальное значение поперечника особи данного вида в ценопопуляции. В нашей выборке таких особей нет, самая большая особь имеет поперечник 12,5 см. Это можно объяснить тем, что рост особей при достижении ими поперечника 8 см сильно тормозится напряженной конкуренцией. Положительное отклонение вариационного ряда от аппроксимирующей логарифмической линии в диапазоне поперечников особей от 4 до 8 см говорит о том, что в этой размерной группе особей больше теоретически ожидаемого. |
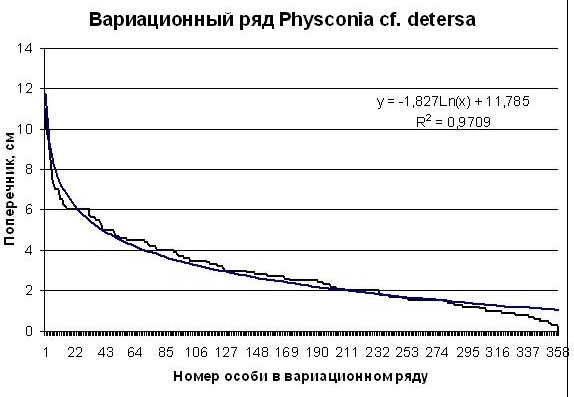
Рис. 3. Вариационный ряд особей Physconia cf. detersa ценопопуляции на шифере крыши на юге Приморского края (черная ломаная линия). Гладкая синяя линия – логарифмическая аппроксимация вариационного ряда. Пояснения в тексте |
Выборка Physconia cf. detersa насчитывает 358 особей. На диаграмме рис. 2 видно, что вариационный ряд особей Physconia cf. detersa более точно апроксимируется логарифмической зависимостью (R =0,9709). Но и в этом случае в популяции наблюдается «недобор» крупных и мелких особей и небольшой «избыток» особей среднего размера. Как и в случае с первым видом мы склонны объяснять это давлением конкуренции в ценопопуляции. Если это так, то ценопопуляция Physconia cf. detersa в данном сообществе испытывает меньшее конурентное давление, чем ценопопуляция Caloplaca sp.1 (накипная). С этим выводом вполне согласуется и большее количество особей фисконии (358 по сравнению со 183 у калаплаки), Измерялись все особи на некоторой ограниченной поверхности, поэтому количество особей в выборках пропорционально их встречаемости на поверхности крыши. По количеству особей вида в выборке можно судить о его обилии в данном лишайниковом сообществе.
В выборке Xanthoria sp. оказалось 146 особей. На диаграмме рис. 3 изображен вариационный ряд второго вида рода Xanthoria sp. (лопастная). Встречаемость этого вида в ценопопуляции меньше, чем у первого вида калаплаки (140 особей). Дефицит крупных и мелких особей и избыток средних в этой ценопопуляции также достаточно высокий (R =0,956). О напряженности конкурентных отношений в ценопопуляции говорит и коэффициент логарифмического уравнения а, в нашем случае он наибольший у Xanthoria sp. (лопастная) (а = -3,174), средний у Caloplaca sp.1 (накипная) (а = -2,8287) и наименьший у Physconia cf. detersa (а = -1,827). Этот коэффициент измеряет степень вогнутости аппроксимирующей кривой. Чем больше степень вогнутости, тем напряженнее конкурентные отношения в ценопопуляции данного вида в данном сообществе, тем больший прессинг испытывают особи в ценопопуляция данного вида как со стороны особей того же вида, так и со стороны особей других видов этого сообщества. |
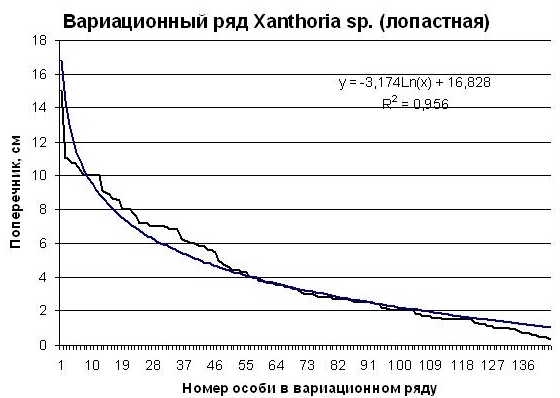
Рис. 4. Вариационный ряд особей Xanthoria sp. (лопастная) ценопопуляции на шифере крыши на юге Приморского края (черная ломаная линия). Гладкая синяя линия – логарифмическая аппроксимация вариационного ряда. Пояснения в тексте
|
Для анализа размерной структуры ценопопуляций трех видов лишайников были построены кривые статистических распределений особей видов в зависимости от величины максимального поперечника особи (рис. 5, 6, 7). Кривые всех трех распределений отчетливо деформированы. У Caloplaca sp.1 (накипная) кривая имеет три деформации. Наибольшее количество особей имеет модальное значение в области 1–2 см. Эту группу образуют особи последней фазы активного возобновления данного вида лишайников. Небольшая деформация имеется в области модального значения поперечника 4 см. Эта группа особей поселилась раньше первой. Вторая деформация в области с модальным значением поперечника 6–7 см, четвертая деформация с модальным значением 10 см самая малочисленная. Особей с поперечником таллома больше 10 см в ценопопуляции очень мало. Может быть, это первопоселенцы с возрастом 50 лет, дожившие до настоящего времени. В таком случае скорость роста таллома у Caloplaca sp.1 (накипная) составляет 13 см в 50 лет по диаметру, или 6,5 см за 50 лет (1,3 мм в год) по радиусу. Однако возможны ошибки при выборе особей для измерения, и самые крупные особи на самом деле представляют собой не 1, а 2–3 слившихся таллома разных особей. В таком случае за максимальный размер особи правильнее взять 10 см (четвертую деформацию). В таком случае скорость роста таллома у Caloplaca sp.1 (накипная) по радиусу в данном районе и в данном сообществе составляет 1 мм в год. |
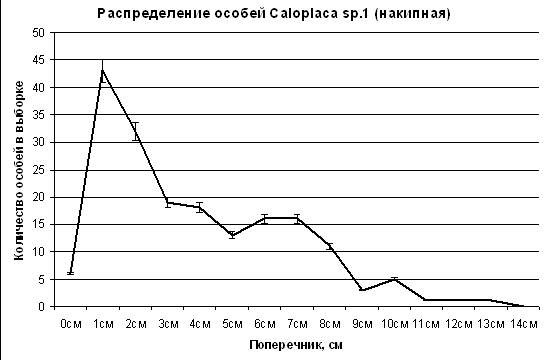
Рис. 5. Статистическое распределение особей Calaploca sp.1 (накипная) ценопопуляции на шифере крыши на юге Приморского края в зависимости от величины поперечника особи. Пояснения в тексте.
|
Четыре деформации на кривой распределения скорее всего соответствуют четырем благоприятным периодам в течение 50 лет, когда ценопопуляция данного вида активно пополнялась новыми особями. Эти периоды были отделены друг от друга периодами менее благоприятными для возобновления Caloplaca sp.1 (накипная). Интересно, что за 50 лет произошло 4,5 одиннадцатилетних цикла солнечной активности. В каждый новый цикл ценопопуляция пополнялась все большим количеством новых особей, так как диазачатки поступали от особей всех предшествующих циклов, количество которых увеличивалось с каждым новым циклом. Таким образом, ритм изменений климата, совпадающий с 11-летним ритмом солнечной активности, отразился в размерной структуре ценопопуляции.
На рис. 6 мы видим, что кривая частотного распределения Physconia cf. detersa имеет не 4, а 3 деформации: в диапазонах поперечника особей 6, 4 и 2 см. Возможно, это связано с тем, что Physconia поселилась позднее, чем Caloplaca, поэтому одна деформация на этой диаграмме выпала. Скорость роста таллома у Caloplaca sp.1 (накипная) составляет 6 см за 50 лет по диаметру (первая положительная деформация), или 3 см за 50 лет (0,6 мм в год) по радиусу. Но если рассчитывать по второй деформации, то скорость роста составляет только 0,4 мм в год.
Кривая распределения вида Xanthoria sp. (лопастная) имеет также 3 положительные деформации в диапазонах значений поперечника особи 10, 6–7 и 1–2 см. В таком случае скорость роста таллома у Xanthoria sp. (лопастная) по первой деформации скорость роста составляет 10 см за 50 лет по диаметру, или 5 см за 50 лет (1 мм в год) по радиусу. Но если рассчитывать по второй деформации, то скорость роста таллома у этого вида составляет 0,7 мм в год по радиусу. |
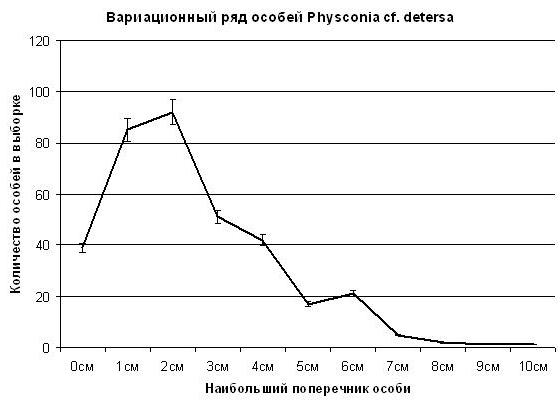
Рис. 6. Статистическое распределение особей Physconia cf. detersa ценопопуляции на шифере крыши на юге Приморского края в зависимости от величины поперечника особи. Пояснения в тексте.
|
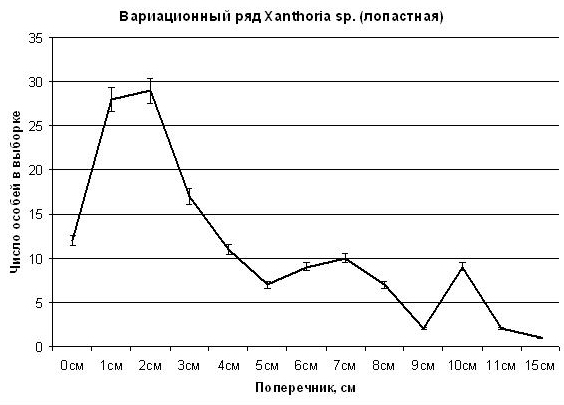
Рис. 7. Статистическое распределение особей Xanthoria sp. (лопастная) ценопопуляции на шифере крыши на юге Приморского края в зависимости от величины поперечника особи. Пояснения в тексте.
|
Проведенное исследование позволило определить скорость нарастания в одну сторону (по радиусу) таллома трех видов лишайников эпилитов на шифере на юге Приморского края: Calaplaca sp.1 – 1–1,3 мм в год, Xanthoria sp. – 0,7–1 мм в год и Physconia cf. detersa – 0,4–0,6 мм в год. Необходимо отметить очень быстрое и эффективное поселение лишайников эпилитов на шифере по сравнению со свежими сколами скальных пород.
Литература
• Галанин А. А. Лихенометрическая индикация динамики современных ландшафтных процессов: Препринт. – Магадан: СВНЦ ДВО РАН, 1996. – 45 с.
• Галанин А.А. Лихенометрия: современное состояние и направления развития метода. – Магадан: СВКНИИ ДВО РАН, 2002. – 74 с.
• Исаенко О. В. Применение лихенометрического метода для датировки средней стены средневековой крепости Чуфут-Кале. Режим доступа: http://www.crimea.edu/internet/Education/geoecology/24.htm
• Мартин Ю.Л. Формирование лишайниковых синузий на моренах ледников Полярного Урала // Автореф. дис. …канд.биол.наук. – Свердловск, 1967. – 20 с.
• Определитель лишайников СССР. – Л., 1975. Вып. 3. С. 85–105.
• Beschel R.E. Lichenometrical studies in West Greenland // Arctic, 1958, V.11. N 4.P. 254.
• Matthews J.A. Experiments on the reproducibility and reability of lichenometric datas, Storbreen gletchervorfeld, Jotunheimen, Norway // Norsk geografisk Tidsskrift, 1975. N 29. P. 97–109.
|

