Структура и динамика экосистем Сибири и Дальнего Востока: сборник научных статей. – Находка: Институт технологии и бизнеса, 2011. – 128 с.
Предисловие C.6-8
Беликович А.В., Василенко Н.А., Галанин А.В., Галанина И.А., Долгалева Л.М., Козырь И.В., Маврин И.Б., Роенко Е.Н., Яковченко Л.С. Динамика растительного покрова (результаты мониторинга на постоянных пробных площадях на юге Дальнего Востока и в Забайкалье С.9-85
Беликович А.В., Галанин А.В., Галанина И.А., Долгалева Л.М. Высотная дифференциация растительности Сохондинского биосферного заповедника С.86-129
Галанина И.А. К вопросу о скорости роста лишайников эпилитов на юге Приморского края С.130-143
Киселёва А.Г. Сохранение биоразнообразия сосудистых растений морских побережий Приморского края С. 144-156
|
УДК 581.55
Динамика растительного покрова (результаты мониторинга на постоянных пробных площадях на юге Дальнего Востока и в Забайкалье)
Беликович А.В., Василенко Н.А., Галанин А.В., Галанина И.А., Долгалева Л.М.*, Козырь И.В., Маврин И.Б., Роенко Е.Н., Яковченко Л.С.
Ботанический сад-институт ДВО РАН, г. Владивосток
*Институт технологии и бизнеса, г. Находка
Vegetation cover dynamics (results of monitoring on constant sample plots in the southern Russian Far East and Transbaikalia)
Belikovich А.V., Vasilenko N. A., Galanin А.V., Galanina I. A., Dolgalyeva L.М.*, Kozyr I.V., Mavrin I.B., Roenko E.N., Yakovchenko L.S.
Botanic Garden Institute of the Far-Eastern Branch of the Russian Academy of Science (BGI FEB RAS), Vladivostok
*Institute for Technology & Business, Nakhodka
Ключевые слова: мониторинг, растительность, растительный покров, постоянные пробные площади
Глобальные изменения климата выдвигают на первый план проблему прогноза процессов, которые происходят в растительном покрове. Эти изменения зачастую наносят колоссальные убытки экономике. Например, длительная многолетняя засуха в Читинской области, способствовала распространению лесных и степных пожаров на огромные территории, в результате этой засухи в регионе стало нерентабельным земледелие. К сожалению, мониторинговые исследования растительного покрова в этом регионе все еще не находят должной поддержки со стороны Академии наук и правительств областей и краев. Как и 20 лет назад, в стране отсутствует государственная программа мониторинга растительност., Финансирование подобных исследований по-прежнему явно несоизмеримо с масштабами проблемы.
Vegetation dynamics in Dahuria have been studied on the system of constant sample plots – relevees. The Dahuria, which includes Northern Mongolia, Khingan Mtsin China, Buryatiaand Transbaikaliain Russia, is characterized by the original weather rhythm – 5 phenological seasons including extremely dry earlier summer (April 20 – July 20) and wet late summer (July с 20 – September 10). The latter is favor able for mesophytes' growth and development. The rhythm has been main tained in Dahuria for many thousands years thanks to Pacific mon soon. Forsuch long period, two original vegetation types have been formed in local landscapes: Dahur prairie and Dahur kharganat. Dahur prairieis a combination of xerophytic plants, mesophytes and meso-xerophytes. Vegetation monitoring showed that for the last 40 years obvious climate warming and aridization in Dahura takes place resulting in the increase in soil moisture in ecosystems of the upper forest belt due to intense permafrost thawing in alpine and subalpine belts. In the alpine communities of Khentei Mts, many species started to degrade, because there has become less snowing and snow shelters in the talus places disappeared. For example, Pinus pumila as well as Abies sibirica decreased the irs populations due to climate warming. Vegetation of different habitats has been changed with different race in climate perturbations. Instability of weather conditions year by year and oscillations of humidity-aridity of Central Asia climate cause a phenomenon of many-species communities (competition processes act not enough time to replace xeric or mesic species in communities). In wet years, mesophytes turn out to be more competitive, in dry years – plant xerophytes happen to be more stable. High taonomic diversity of Dahuria prairie proves that conditions here have been very unstable for significantly long period.
Введение
Под мониторингом растительного покрова , или ботаническим мониторингом (БМ), мы понимается специальное длительное слежение за его состоянием (флорой и растительностью) на постоянных пробных площадях и ключевых участках (Галанин, 2006, 2009) . БМ – это один из главных методов изучения динамики растительного покрова под воздействием естественных и антропогенных факторов. К сожалению, из-за трудоемкости БМ, до сих пор основным методом изучения динамики РП является метод трансформации пространственных рядов в ряды временные. В этом случае подбираются пробные площади с растительным покровом, находящимся на разной стадии восстановления, затем эти площади выстраиваются в гипотетический ряд последовательных смен, и этот ряд интерпретируется как временной ряд изменения растительного покрова в одном месте, т.е. на одной пробной площади. Превалирование этого метода над длительным мониторингом стало причиной создания ряда «геоботанических мифов» и теоретических заблуждений. К числу таких мифов принадлежит, например, учение о климаксовой растительности. В отличие от метода трансформации пространственных рядов в ряды временные, БМ позволяет выявлять и изучать не только демутационные смены растительных сообществ, но и необратимые изменения растительного компонента экосистем. Пренебрежение методом БМ привело геоботаников к переоценке роли сукцессионных процессов в динамике растительного покрова над необратимыми изменениями растительности.
На первом этапе развития геоботаники преобладало стремление выделить элементарные объекты и явления в РП, научиться распознавать, описывать и классифицировать их. На разработку классификационной парадигмы в геоботанике ушло около 200 лет. И сегодня описательно-классификационный этап все еще продолжается, так как в Сибири и на Дальнем Востоке немало неизученных в геоботаническом отношении территорий. Однако, начиная с середины ХХ века, в геоботанике усиленно развивается динамическая парадигма, суть которой – изучение закономерностей динамики растительного покрова, так как нерешенность именно этих проблем тормозит развитие общей и прикладной экологии.
Сукцессия растительности – это последовательный ряд смены серийных растительных сообществ на конкретном местообитании после выведения конкретной экосистемы из состояния динамического равновесия. В результате сукцессии на конкретном местообитании восстанавливается исходное растительное сообщество, называемое геоботаниками климаксовым, или коренным . Коренное сообщество растений устойчиво и в данных климатических условиях не изменяется. В результате сукцессии растительности и животного населения конкретная экосистема возвращается в свое исходное состояние и пребывает в нем до тех пор, пока не изменятся климат, рельеф, гидрологический режим, пока не произойдет пожар, или не случится какая-то другая катастрофа. Если в лесу или степи прошел пожар, если лес вырубили, луг или степь распахали и потом забросили, то, скорее всего, сукцессия на нарушенном участке завершится восстановлением исходного коренного сообщества. Однако если изменился климат, понизился или повысился уровень грунтовых вод, сообщество растений, выведенное из состояния равновесия, в процессе динамики и смены фитоценозов не восстановится. Ряд сукцессионных смен в этом случае пройдет, но завершится эта серия смен новым коренным сообществом, которое также будет находиться в состоянии относительного равновесия с внешней средой, но оно будет существенно отличаться от исходного коренного сообщество. Нередко в районах с неустойчивым муссонным климатом коренные сообщества вообще не успевают сформироваться. Существенные для растительности подвижки климата происходят здесь быстрее, чем длится сукцессионный процесс. Поэтому растительный покров на значительных площадях находится в таких районах в перманентно неравновесном состоянии.
В континентальных районах с более устойчивым климатом коренные растительные сообщества в ходе сукцессий успевают восстанавливаться. Одним из таких районов является Южное Забайкалье. Однако и здесь в условиях глобального изменения климата в растительном покрове происходят необратимые изменения. Так, например, мониторинг растительного покрова на постоянных пробных площадях, заложенных нами в 1982–1984 гг. в Сохондинском заповеднике показал, что нарушенная коренная растительность за сукцессионный цикл не успевает восстанавливаться, в результате происходит смена одних коренных растительных сообществ другими. Климат изменяется быстрее, чем заканчивается один сукцессионный цикл. Следовательно, в современную климатическую эпоху нарушенная коренная растительность в результате сукцессионных смен сформироваться не успевает, и поэтому в растительном покрове преобладают серийные сообщества.
Геоботаниками за полтораста лет были составлены сотни тысяч геоботанических описаний, но почти все они были разовыми и не предполагали ревизии – повтора через определенные промежутки времени. Для типизации и классификации растительности этих разовых описаний было достаточно, но изучать динамику растительности на этом фактическом материале можно было только весьма поверхностно. Для этого были необходимы длительные наблюдения на постоянных пробных площадях (пп). Закладывать и описывать пп пытались на стационарах и в заповедниках. Но старел исследователь, заложивший пп, уезжал или умирал, начиналась война, приходили новые сотрудники с иными интересами, иными методами исследований, старые пп забрасывались, терялись. Когда через 30–40 лет в этот район приходил сотрудник, заинтересованный в мониторинге растительности, то он не мог отыскать заброшенные площади и был вынужден закладывать новые.
Много постоянных пробных площадей закладывалось при лесоустройствах. Древостои на них характеризовались весьма подробно: деревья нумеровались, измерялись, наносились на план, учитывались возобновление и подрост, однако остальные ярусы растительности характеризовались весьма поверхностно и небрежно, так как главной целью лесоустройств было определение запаса древесины. Флора на этих площадях выявлялась едва на 10–20%, так как специалистов флористов в таких экспедициях не было. Да и трудно себе представить, чтобы в лесоустроительной экспедиции, базирующейся, например, в Минске, были флористы, ориентирующиеся в биоразнообразии растительного покрова и Белоруссии, и Северного Кавказа, и Южной Сибири, и Дальнего Востока. А работать сотрудникам лесоустроительных экспедиций приходилось во всех районах огромного Советского Союза. Завышенные нормативы для сотрудников лесоустроительных экспедиций и слабый контроль за их работой нередко приводили к откровенной халтуре. Мне и моим коллегам, пытавшимся провести ревизию постоянных пробных площадей, заложенных лесоустроителями в заповедниках, нередко приходилось сталкиваться с такой халтурой. Иногда неверно определялись не только виды трав и кустарников, но и деревьев. Так, для Алханайского национального парка в северной части Хэнтея на одной пп была указана чозения (Chozenia arbutifolia). При проверке чозения оказалась тополем душистым (Populus suaveolens). Лиственницу даурскую (Larix dahurica) лесоустроители приняли здесь за лиственницу сибирскую (Larix sibirica).
Мониторинг растительного покрова должен проводиться на разных уровнях в соответствии с хорологической (пространственной) дифференциацией биосферных систем. Это может быть уровень ландшафтного геоботанического района, уровень мезокомбинации растительного покрова, уровень микрокомбинации и растительного сообщества, или контурфитоценоза (Беликович, 2001, 2005). Если мы намереваемся вести мониторинг растительного покрова геоботанического района, то нам следует заложить постоянные пробные площади во всех типах мезокомбинаций и контурфитоценозов, характерных для этого района. С помощью одной пробной площади размером 1 га проводить мониторинг растительного покрова целого заповедника невозможно. Для этого для равнинного геоботанического района необходимо заложить не менее 10–12 постоянных пробных площадей размером 1 га, а для горного района – не менее 30–40. Именно к такому выводу пришло большинство исследователей, работавших в разных регионах северной Евразии.
Мониторинг растительных сообществ – это мониторинг одного из компонентов локальной экосистемы. Он должен учитывать характерное время или хроноинтервал этой экосистемы. Хроноинтервал экосистемы – это время, необходимое для возвращения данной экосистемы в равновесное состояние после отклонения от него. Иными словами, хроноинтервал – это время релаксации экосистемы. Для колебательных систем хроноинтервал – это время полного периода колебания. Для большинства лесных экосистем ранга мезокомбинации растительного покрова хроноинтервал составляет 150–200 лет, для степных экосистем – 50–100 лет, для луговых – 20–30 лет. Но для экосистемы целого геоботанического района (элементарной биосферной системы) хроноинтервал имеет размер 1500–2000 лет (Миркин, 1985; Галанин, 1993; Галанин, Беликович, 2002). Для биосферной системы еще более высокого ранга (физико-географической области) хроноинтервал составляет время порядка 10000–20000 лет. Считается, что хроноинтервал биосферы в целом свыше 100000 лет.
Исследования последних 30–40 лет показали, что для изменений климата характерна цикличность. При этом существует не один, а несколько циклов с разными периодами, где короткие циклы накладываются на циклы более длительные. Хорошо доказан и обоснован 11-летний цикл колебаний климата, который связывают с колебаниями солнечной активности. Многие авторы указывают на наличие в природе 90–100-летнего цикла. Некоторые исследователи считают, что еще существует 600–700-летний цикл, отражающийся в биосфере, в том числе и в растительном покрове. Если мы сравним длительность этих циклов с характерным временем экосистем разного ранга, то увидим, что эти хроноинтервалы меньше, чем длительность одного цикла. Следовательно, растительность, выводимая из равновесия периодическими изменениями климата, никак не может восстановиться полностью. Пока такая растительность восстанавливается, наступает новый климатический цикл, равновесие в экосистеме сдвигается, и снова идет сукцессия. Сукцессия растительности в этом случае становится перманентным состоянием растительного покрова.
В условиях перманентных колебательных изменений климата в растительном покрове выработался специальный механизм, который позволяет экосистеме быстро перестраиваться, изменяя состав видов доминантов. Если мы проанализируем видовой состав растительного сообщества, то увидим, что в нем уживаются виды самого разного склада, имеющие разные пределы экологической толерантности. В одной части климатического цикла на данном участке доминируют одни виды, а в другой части активизируются другие виды, а бывшие доминанты переходят в разряд сопутствующих. Такими парами в лесах среднего Сихотэ-Алиня, например, являются дуб монгольский и лиственница даурская, кедр корейский и ель аянская, а в лесах Хэнтей-Чикойского нагорья в Даурии – кедр корейский и лиственница даурская, сосна обыкновенная и лиственница даурская.
При анализе изменений растительного покрова исследователь всегда должен задаваться вопросом о термодинамической направленности этих изменений. Одни изменения приводят к росту энтропии в открытой системе, а другие, напротив, к ее снижению. В первом случае экологическая система движется к состоянию равновесия со средой обитания, а во втором удаляется от него. При катастрофических изменениях энтропия экосистемы увеличивается, а при сукцессионных снижается. Как исследователь может определить термодинамическую направленность изменения экосистемы и растительности как ее части? Ответить на этот вопрос можно, только анализируя динамику биоразнообразия растительного сообщества. Если биоразнообразие экосистемы в ходе ее изменения снижается, то экосистема деградирует, энтропия ее возрастает. Напротив, если биоразнообразие экосистемы при этом увеличивается, – энтропия ее снижается, и система развивается.
Таким образом, важной для мониторинга растительности является оценка биоразнообразия растительных сообществ. Проблема эта не так проста, как это представляется сегодня большинству исследователей. Чаще всего биоразнообразие оценивается как таксономическое разнообразие, отождествляется с числом видов. При этом вряд ли кто скажет, в каком случае разнообразие выше – когда в сообществе 20 видов одного рода, или когда в нем 5 видов, но из 2 разных родов. А когда в сообществе только 2 вида, но из 2 разных семейств – это больше, или меньше? По-моему, эту задачу геоботаники пока даже и не пытались решать. Но задача эта еще сложнее, например, если мы станем рассматривать не таксоны, а экобиоморфы растений. Порой таксономически близкие виды растений принадлежат к совершенно разным экобиоморфам и, наоборот, таксономически несходные виды растений имеют сходные экобиморфы. Геоботаники знают, что биоморфологическое разнообразие чаще всего не совпадает с таксономическим. Но именно биоморфологическое разнообразие является более важным с точки зрения функциональной структуры растительного покрова.
Для поддержания биогеохимического равновесия экосистем важно не таксономическое разнообразие само по себе, а разнообразие функциональное, или биоморфологическое. Мерилом функционального разнообразие растительного покрова может служить число экобиоморф. В конечном счете, таксономический процесс – это «поставщик» не только видового и родового разнообразия растительного покрова, но и разнообразия функционального биоморфологического. Несмотря на то, что таксономическая структура, в конечном счете, подчинена функциональному или биогеохимическому аспекту организации биоты, она, несомненно, обладает высокой автономностью от нее (Любищев, 1982). Таксономический и биоморфологический процессы могут быть инвариантны друг другу. На многоаспектность структурной организации растительного покрова указывал, например, Б.Н. Норин (1980, 1987). Экобиоморфы растений следует выделять как экологические категории в широком смысле, учитывая соотношение между массой многолетних побегов – стволов и сучьев – и ежегодно обновляемой фитомассой листьев, учитывая общий габитус растений, сроки активной вегетации, способы экспансии на свободную территорию, форму перенесения неблагоприятных условий и т.п. (Мазуренко, Хохряков, 1977). Виды же и таксоны более высокого порядка выделяются по сходству генотипов, а не фенотипов.
Экобиоморфы растений – это своего рода их адаптивные стратегии в растительном покрове. Число таких стратегий ограничено. Ценогенез и экогенез лишь опосредованно связаны с филогенезом. Так, становление кислицы обыкновенной (Oxalis acetosella) наверняка происходило под тенистым пологом широколиственных лесов. Однако особи этого вида, благодаря характерной для них жизненной форме, могут расти и в хвойно-широколиственных, и в темнохвойных, и в тенистых мелколиственных лесах. Сходные экологические ниши могут возникать в таксономически несходных экосистемах. Биоморфологически весьма сходными могут быть виды из разных родов, например из родов Picea, Tzuga, Pseudozuga, а виды из одного рода порой могут иметь очень несходные жизненные формы, например Salix polaris и S. caprea. Мы считаем, что функциональная (не таксономическая) стратификация фитоценозов – важнейшая задача геоботаники и экологии.
Классификация растительности по доминантным видам и экобиоморфам растений никак не может быть заменена флористической классификацией растительности. Мы считем, что, описывая растительность постоянных пробных площадей, следует очень скрупулезно описывать и учитывать вертикальную надземную и подземную ярусность, фенологическую неоднородность, способы возобновления ценопопуляций растений, способы перенесения растениями неблагоприятных условий и другие биоморфологические особенности.
Благодаря способности к миграции виды растений получают шанс пережить резкую смену условий, «уходя» из прежних районов и прежних мест обитания в новые, ставшие для них в изменившихся условиях для них вполне пригодными. Но и прежние виды, которые образовывали растительный покров района в прежнюю климатическую эпоху, способны длительное время сохраняться в растительном покрове на уникальных местообитаниях, резко снижая при этом свою активность в растительном покрове. Таким образом, в результате миграций видов, переживания ими изменившихся условий в уникальных местообитаниях и за счет автохтонного видообразования таксономическое богатство локальных флор увеличивается. Однако при очень резком изменении климата происходит массовое вымирание видов, и это вымирание не компенсируется ни мигрантами из соседних флор, ни новыми видами, образующимися в процессе видообразования. В этом случае таксономическое богатство локальных флор снижается, их энтропия увеличивается.
Разнообразие видов в одном фитоценозе также зависит от устойчивости или неустойчивости условий в данной локальной экосистеме. Например, повышенное потребление зеленой массы хвои и листвы делает леса более сквозистыми. Тем самым под пологом леса увеличивается освещенность, изменяется фитоклимат для растений нижних ярусов. В таких условиях большую роль в продукционном процессе начинают играть растения нижних ярусов, которые не имеют мощных многолетних стволов и, таким образом, способны резко ускорять биологический круговорот. Выедая преимущественно доминирующие виды растений, травоядные животные тем самым препятствуют их безграничному лидерству в сообществе и способствуют развитию тех видов, которые в обычных условиях (без выедания) не смогли бы выдержать конкуренцию со стороны лидера и выпали бы из состава сообщества (Рафес, 1980).
В Южном Забайкалье при вспышках численности непарный шелкопряд способен полностью уничтожить хвою у лиственницы (Larix dahurica, L. sibirica), как это было в 2001 г. При этом происходит осветление полога леса, замедляется работа корневых систем лиственниц, создаются благоприятные условия для роста и развития травянистых растений, кустарников и возобновления березы плосколистной (Betula platiphylla) или осины (Populus tremula). Если бы нашествий непарного шелкопряда в Забайкалье не случалось, то участие березы и осины в сложении древостоя лиственничных и березово-лиственничных лесов резко снизилось, а вместо травяного покрова в таких лесах сформировался бы устойчивый мохово-лишайниковый покров.
Локальная (конкретная) флора понимается нами как общий генофонд растительного покрова ландшафтного района. При формировании каждого конкретного фитоценоза в пределах конкретной флоры отбор подходящих компонентов (видов) осуществляется из большого числа «претендентов», диазачатки которых попали в данное местообитание. Таким образом, при увеличении разнообразия и интенсивности семенного дождя вероятность формирования экологически более оптимального растительного сообщества увеличивается. Благодаря многообразию и большому совершенству способов распространения спор, семян и плодов растений на большие расстояния, мы можем рассматривать конкретную флору как некий экологический потенциал и можем соотносить этот потенциал с каждым конкретным местообитанием, каждой конкретной экосистемой (Галанин, 1981, 2005, Галанина, 2005, 2008). Интенсивность и разнообразие семенного дождя на каждой участок в пределах ландшафтного района значительно выше экологической и ценотической емкости конкретного фитоценоза. Эта интенсивность и качество семенного дождя и определяют флористический потенциал конкретной флоры в конкретном фитоценозе.
В каждом конкретном растительном сообществе есть виды, представленные большим количеством особей, которые находятся здесь в состоянии близком к эколого-ценотическому оптимуму, и есть виды, представленные меньшим количеством угнетенных особей, для которых условия в данном сообществе не оптимальны. Малообильные виды практически не принимают участия в биогеохимической работе, совершаемой данным фитоценозом, но они образуют его сверхразнообразие на случай, если резко изменятся климатические условия, тогда виды доминанты окажутся за пределами своей экологической толерантности, а виды малоактивные вновь станут активными.
Кедр сибирский (Pinus sibirica) в Хентей-Чикойском нагорье образует чистые и смешанные древостои в верхнем лесном поясе на высоте более 1800 м н.у.м. Тем не менее, в лиственничных лесах и в нижнем лесном поясе в Хэнтей-Чикойском нагорье кедр сибирский часто можно встретить в возобновлении. Однако он в этом поясе не может выйти в полог и гибнет в состоянии возобновления или подроста. Причина гибели подроста кедра в нижнем лесном поясе – отсутствие достаточного снегового укрытия зимой. Но кедровки постоянно заносят орехи кедра в леса нижнего пояса, и всходы здесь появляются довольно часто даже на высоте 1400 м н.у.м. При очередной гумидизации климата особи кедра в таких лиственничниках получат шанс выйти в полог, и нижняя граница кедровых и лиственнично-кедровых лесов снизится на 100–200 м н.у.м. Н.П. Соколов, посетивший голец Сохондо в XVIII в., отмечал, что нижняя граница кедровых лесов тогда проходила примерно на 150 м ниже, чем в современную эпоху (Паллас, 1788). Таким образом, сверхразнообразие растительных сообществ позволяет фитоценозу при резком изменении климатических условий очень быстро перестроиться (а по сути, смениться другим фитоценозом с другими доминантами).
Лиственница даурская и кедр сибрский на подгольцовой террасе горы Сохондо неплохо возобновляются и выше современной границы леса, семена их регулярно и в большом количестве заносятся сюда с нижней облесенной террасы, но древостоя здесь они не образует из-за жестких климатических условий. Особи их здесь часто имеют карликовую «флагообразную», «юбочную», или форму стланника, в высоту деревца лиственницы и кедра на этой террасе едва достигают 1–1,5 м, но в таком угнетенном состоянии они живут до 200 и более лет (фото 1). Однако при незначительном потеплении эти деревца в течение 50–60 лет сформируют на нынешней подгольцовой террасе нормальный кедрово-лиственничный древостой с подлеском из кедрового стланика такой же, какой сегодня растет на склонах и террасах на 100 м ниже.
Процессом видообразования охватываются многочисленные ценопопуляции вида в пределах его обширного географического ареала. Панмиксия удерживает их в рамках одной генетической системы, поэтому вид с обширным ареалом эволюирует как единая генетическая система. Чем устойчивее генетический аппарат у вида, тем большим по протяженности (при прочих равных условиях) может быть его географический ареал, так как панмиксия в этом случае может «цементировать» весьма удаленные друг от друга в пространстве ценопопуляции. Разные ценопопуляции одного и того же вида обитают в разных экологических условиях, при этом особи в этих ценопопуляциях нередко имеют разную экобиоморфу. Например, кедровый стланик (Pinus pumila ) в лесу и в тундре образует разную биоморфу: в лесу это стелющееся дерево, а в тундре – кустарник. Данный феномен несовпадения генетических и эколого-ценотических векторов изменения вида как системы является одним из механизмов, препятствующих сужению экологической амплитуды видов. Это также, несомненно, один из механизмов поддержания определенного биотического разнообразия и стабильности биосферы в целом.
Совмещение особей разных видов и разных экобиоморф в одном фитоценозе приводит к их экологическому и ценотическому взаимодействию. Каналом взаимодействия в этом случае служит внешняя среда. При этом формируются системы особой функциональной интеграции – ценоэлементы. В настоящее время собран достаточный фактический материал, позволяющий говорить о ценоэлементах как об организованных системах. Взаимодействие совместно произрастающих особей разных видов приводит к их упорядоченному распределению в пространстве относительно друг друга, в результате чего в фитоценозе возникают группы положительно сопряженных друг с другом видов, особи которых в непосредственной близости друг к другу встречаются чаще, чем это следует из теории вероятностей, предполагающей их случайное распределение. Исследованиями Н.А. Василенко (2008) было доказано, что даже в том случае, когда население фитоценоза состоит из особей одного вида, происходит дифференциация особей по мощности на вполне дискретные группы и закономерное распределение особей разной фитоценотической мощности относительно друг друга. В этом случае фитоценотическая система «творит свое» биологическое разнообразие, дифференцируя особи одного и того же вида на экотипы, классы толщины, классы высоты и т.д. Такое явление мы называем самоорганизацией фитоценотической системы, благодаря которому она не только поддерживает биоразнообразие, но и создает его.
«Рисунок» растительного покрова, называемый его ландшафтно-экологической структурой, в значительной степени повторяет «рисунок» экологического фона ландшафта. Однако экофон ландшафта (Кожевников, 1989; Галанин, 2005) и растительный покров не находятся в отношениях типа «форма-отливка». Растительность, формируясь, вносит весьма существенную «корректировку» в экофон, трансформируя его в новое состояние, производное от исходного. Чем полнее выработана ценотическая структура, тем больше степень трансформации экофона ландшафта. Экофоны ландшафтов в чистом виде, не трансформированные растительностью, следует рассматривать как некую абстракцию, введение которой полезно только для четкой формулировки основных положений теории растительного покрова и только.
Гомеостаз – стабильность экосистем – понимается нами как тенденция к поддержанию круговорота вещества в таком состоянии, что вещество возвращается обратно к производителям (May, 1976; Breymeyer, 1979) и не покидает конкретную экосистему, не выходит за ее пределы. В процессе сукцессии происходит выработка механизмов гомеостаза так, что коренная растительность биогеохимически более стабильна, чем пионерная. В природе возможна смена типа гомеостаза экологической системы, например, такая смена происходит при антропогенном воздействии или существенном изменении климата, когда леса заменяются лугами, а на месте степей возникают посевы монокультур, когда лес наступает на тундру, а степь наступает на лес. Тип гомеостаза экосистем может измениться в результате пожаров, выпаса скота и т.д.
В грубом приближении формирование и поддержание ландшафтно-экологического «рисунка» растительного покрова можно рассматривать как сортировку видов и экобиоморф по экотопам, а по мере формирования ценотической структуры растительного покрова как сортировку видов по биотопам и ценоэлементам. В процессе такой сортировки подбираются виды и экобиоморфы, способные к совместному существованию в определенных местообитаниях. Образующиеся при этом сочетания видов и экобиоморф (ценоэлементы) должны быть не только комплементарны экотопу, но и комплементарны друг другу. Подбор видов и экобиоморф в фитогеохоры происходит поэтапно в ходе сукцессии и является эколого-ценотическим процессом, а не чисто экологическим, как например первая стадия сукцессии на промышленных отвалах. «Рисунок» растительного покрова в ландшафте определяется: 1 – неоднородностью исходного экологического фона, 2 – характером локальной (конкретной) флоры (флористический потенциал), 3 – ценотическим подбором растений в оптимальные ценотические системы.
Целью настоящей статьи является анализ некоторых результатов многолетних наблюдений на постоянных пробных площадях, заложенных в разных типах растительности и разных высотных поясах в некоторых провинциях и подобластях Восточноазиатской ботанико-географической области.
Объекты и методы
Растительный покров Дальнего Востока и Даурии представляют собой динамичную ботанико-географическую систему в ранге области (рис. 1), претерпевающую изменения, связанные с вековыми колебаниями климата. Здесь имели место миграции видов растений из разных флористических областей: Юго-Восточной Азии, Центральной Азии, Южной Сибири, Северо-Востока Азии. Вектор миграций закономерно менялся по мере прохождения векового климатического цикла.Особенности горного рельефа Даурии обеспечивали и обеспечивают длительное сосуществование видов разных географических и генетических групп в конкретных флорах этой территории, а также интрогрессивную гибридизацию видов в пределах секций и родов, что является причиной явления, которое можно назвать таксономическим континуумом. Попадая в несвойственные условия в пределах климатического цикла, виды растений изменяют свою жизненную форму сначала в пределах толерантности вида, затем, изменяется вектор естественного отбора, что, несомненно, ускоряет процессы видообразования.
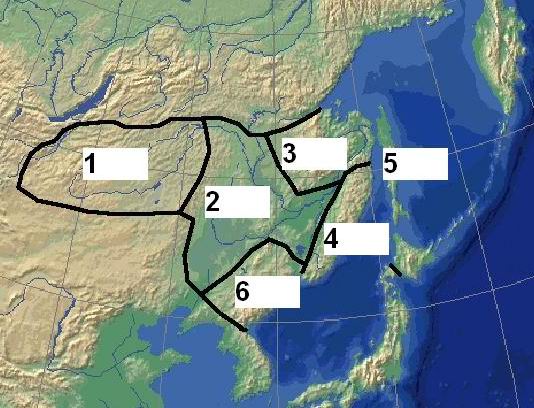
Рис. 1. Границы Восточно-Азиатской ботанико-географической области и ее деление на подобласти: 1 – Даурская; 2 – Маньчжурская; 3 – Нижнеамурско-Северосахалинская; 4 – Сихотэ-Алинская; 5 – Сахалино-Хоккайдская; 6 – Корейско-Хасанская (Галанин, Беликович, 2006; Galanin , Galanina , 2010) |
Система заповедников, заказников и национальных парков (особо охраняемых природных территорий или ООПТ) юга Дальнего Востока и Забайкалья в целом репрезентативна относительно биологического и эколого-ценотического разнообразия ее растительного покрова. Заповедники, как правило, это эталонные территории, на которых сохраняются естественные ландшафты в ненарушенном состоянии. По этой причине такие территории являются более подходящими для проведения мониторинговых исследований растительного покрова. Но методика мониторинга должна быть унифицированной для всех заповедников и национальных парков, только тогда результаты наблюдений будут сравнимы между собой. По нашему мнению, ООПТ обязаны осуществлять экологический мониторинг, после осуществления охраны это должна быть их важнейшая задача. Мониторинг растительного покрова в ООПТ должен проводиться на разных уровнях и в соответствии с пространственной дифференциацией биосферных систем. Это может быть уровень ландшафтного геоботанического района, уровень мезокомбинации или микрокомбинации растительности. Для мониторинга растительного покрова геоботанического района следует закладывать постоянные пробные площади во всех типах мезокомбинаций, характерных для этого района. В пределах пробных площадей мониторинг проводится на уровне отдельных особей, парцелл и микрогруппировок. Для этого они должны наноситься на план постоянной пробной площади и периодически оцениваться по стандартной методике, которая должна быть едина на протяжении всего периода мониторинга.
В качестве модельных районов нами использованы: Сихотэ-Алинский биосферный заповедник (Галанина, 2005), Сохондинский биосферный заповедник, Алханайский национальный парк. Наши исследования показали, что для мониторинга растительности в пределах равнинного геоботанического района необходимо заложить не менее 10–12 постоянных пробных площадей (пп) размером 0,5 га каждая, а для горного района – не менее 30–40 таких площадей (Галанин, 1991; Беликович, 2001). В Сохондинском заповеднике мониторинг |
растительности на пп начат в 1982 г. (Галанин, 1984; Галанин, Беликович, 1988;). Тогда же нами разработана и предложена для заповедников СССР программа и методика длительного геоботанического мониторинга. Эта программа представлена в Главохоту РСФСР и в Комиссию АН СССР по заповедному делу (Галанин, 1984). В последующие годы эта методика развивалась и совершенствовалась (Галанин Беликович, 2001; Галанина, 2006; Vasilenko, Galanin, 2009; и др.)
Согласно нашей методике в пределах заповедника отбирается 30–35 модельных урочищ с наиболее характерными для данного заповедника типами растительности. В пределах каждого типа урочищ закладывается одна пп размером 50х50 м или 50х100 м. Наибольшая сторона площади должна быть вытянута поперек склона. Совокупность пп в заповеднике должна охватывать все основное разнообразие растительного покрова определенного ландшафтно-ботанического района. В этой выборке должна быть представлена не только растительность, занимающая большие площади, но и растительность реликтовая. Скорее всего, именно мониторинг реликтовой растительности позволит установить направление происходящих в растительном покрове изменений. Постоянная пробная площадь внутри себя должны быть максимально однородна по положению в рельефе, характеру почвы, крутизне и экспозиции склона, уровню залегания грунтовых вод, характеру подстилающих горных пород. Неоднородность растительности в пределах пп в основном может носить ценотический, но не эдафический характер. По углам пп маркиру e тся столбами высотой 1,3 м и снабжается дощечкой, или металлической (пластиковой) пластинкой с надписью. В надписи обязательно указывается номер площади и дата ее закладки. Надпись делается так, чтобы она могла сохраниться в течение, как минимум, 10–15 лет до следующей ревизии пробной площади.
Положение каждой пп указывается на картосхеме заповедника, методом космической навигации определяются координаты центра пробной площади и всех ее углов. На каждую пп заводится паспорт, в котором будут накапливаться результаты ревизий в течение многих десятилетий. Кроме номера пп, в паспорте указываются ее координаты, высота над уровнем моря, дается подробное описание положения в рельефе, делается привязка к хорошо заметным и долго сохраняющимся ориентирам (вершины гор и холмов, повороты рек и ручьев, отдельные скалы или крупные камни). При описании пп детально характеризуются: положение ее в макро- и мезорельефе, крутизна и экспозиция склона, почва, материнская порода, уровень грунтовых вод. Результаты глазомерных и инструментальных измерений указанных параметров с указанием даты измерений также помещаются в паспорт площади.
Каждая площадь должна разбивается на постоянные квадраты 10х10 м, примыкающие друг к другу. По углам квадраты маркируются очищенными кольями высотой 1,3 м и диаметром 5–8 см. Деревянные колья могут быть заменены металлическими или пластиковыми. При следующей ревизии сгнившие колья заменяются новыми. Составляется план площади, на котором квадраты 10х10 м нумеруются, нумерация квадратов, заданная при закладке площади, сохраняется при всех последующих ревизиях.
При описании растительности площади выделяются ярусы древостоя (для лесных ценозов), измеряются их средняя высота, сомкнутость каждого яруса, указываются доминирующие в каждом ярусе виды, оцениваются количественно подрост и возобновление, отмечается общее проективное покрытие по ярусам и главным растительным синузиям (мохообразные, мхи эпифиты, кустистые эпигейные лишайники, накипные лишайники эпилиты, лишайники эпифиты, травянистые растения, папоротники, кустарнички, кустарники, лианы). Составляется таблица распределения всех видов сосудистых растений по квадратам 10х10 м с указанием обилия каждого вида в каждом квадрате. Обилие вида в квадрате оценивается глазомерно по пятибалльной шкале. Дифференцированно по квадратам оценивается проективное покрытие основных синузий растительности. На детальный план пп наносятся границы микрогруппировок и парцелл, которые могут быть выделены визуально. Обязательно указывается дата закладки пп, фамилии и имена сотрудников, проводивших закладку, приводится текст подробного описания. Для точного определения видов растений и лишайников собирается гербарий к каждой пробной площади. Он определяется высококвалифицированными специалистами и хранится в музее заповедника или научно-исследовательской организации, ведущей мониторинг. На гербарных этикетках, кроме традиционной информации, обязательно должен быть указан номер постоянной пробной площади.
Большое значение для мониторинга растительного покрова на пробной площади имеют такие показатели, как отношение запаса вещества в лесной подстилке к величине вещества в ежегодном опаде. Ежегодный опад желательно учитывать по видам деревьев и фракциям (листья, ветви, кора, шишки, плоды). Для улавливания опада в каждом типе парцелл следует установить опадоуловитель – неглубокий ящик с дном размером 1х1 м и высотой до 20–30 см. Дно ящика должно быть из рубероида. Вынимать опад из ящиков следует два раза в год – весной после стаивания снега и поздней осенью после окончания листопада. Все ящики должны иметь постоянные номера, учет опада на них следует проводить диффиренцированно. Положение каждого ящика должно быть нанесено на план пробной площади с указанием номера этого ящика. Перед взвешиванием собранный опад высушивается до воздушно сухого веса, затем, перед вторым взвешиванием, до абсолютно сухого веса.
Ревизию, или повторное описание растительности постоянных пробных площадей, желательно проводить регулярно через 5, в крайнем случае, через 10 лет. При повторном описании восстанавливается система постоянных квадратов, проводится перечет деревьев, отмечаются усохшие и вывалившиеся деревья, измеряются длины окружностей каждого дерева на той высоте, на которой они измерялись в первый раз. Для этого место замера толщины дерева при первом описании отмечается полоской краски на стволе под номером. Нумеруются и картируются новые деревья, которые подросли, и диаметр их стал больше 4 см. Этим деревьям присваиваются следующие по порядку номера после наибольшего при предыдущей ревизии. Ни в коем случае новым деревьям не следует присваивать номера выпавших или усохших деревьев. Все результаты переописания и повторной таксации помещаются в папку – паспорт конкретной пробной площади. Обязательно указывается дата ревизии, фамилии, имена и отчества лиц проводивших ревизию и того, кто проверил и принял эти материалы.
При описании растительности пп все деревья с указанием их номера и видовой принадлежности наносятся на план. Положение каждого дерева на плане показывается с точностью до 20 см. Размещение деревьев с плана переносится в табличную форму, где в системе двух координат, когда осями-координатами являются две перпендикулярные стороны площади, определяется положение каждого дерева. Система координат позволяет анализировать размещение деревьев с помощью компьютера в программе Exel . Кроме нумерованных деревьев, на план наносятся все особи возобновления (ниже 1,3 м и тоньше 4 см в диаметре) с указанием их видовой принадлежности. С помощью бурава для каждой ступени толщины каждого вида деревьев на каждой пробной площади определяется возраст. При этом для каждой ступени толщины следует определить возраст не менее чем у 10 деревьев. Возраст деревьев заносится в таксационную ведомость перечета всех нумерованных деревьев. В перечетной ведомости, кроме номера дерева, указываются его видовая принадлежность, длина окружности ствола на высоте 1,3 м, ярус, к которому принадлежит дерево, оценивается его жизненность в баллах, возраст, если он измерялся.
Для характеристики почвы в пределах каждого типа парцелл при закладке пп необходимо сделать по одному почвенному разрезу в каждом типе. При описании почвы рекомендуется использовать стандартную методику выделения почвенных горизонтов, необходимо отметить их мощность, характер перехода одного горизонта в другой, цвет, механический состав, наличие горизонта оподзоливания, наличие или отсутствие пятен оглеения. Особое внимание при описании почвы следует уделить описанию самых верхних гумусированных и оторфованных горизонтов, наличию в почвенном профиле угольков, кутан и др. Из каждого горизонта при первоописании площади должны быть взяты образцы для почвенно-химического анализа. Результаты этого анализа также следует вносить в паспорт пробной площади. Места почвенных разрезов следует нанести на план пробной площади, а если они взяты по соседству с ней, то на план данного урочища. Надо помнить, однако, что не следует отбирать образцы почвы от пробной площади дальше, чем на 10 м.
Определенную информацию о состоянии растительности и почвы содержат фотографии. При первом описании и последующих ревизиях пп следует делать фотографии общего характера растительности, основных микрогруппировок, портреты видов растений, главных синузий и т.д. Желательно фотографировать при последующих ревизиях с одних и тех же точек.
При обработке данных мы использовали метод построения вариационных рядов деревьев в зависимости от их толщины. При этом все деревья располагаются в ряд по мере убывания диаметра ствола или длины его окружности, каждое дерево изображается в виде колонки, длина которой соответствует толщине дерева, при этом все колонки на диаграмме одинаковой ширины. Таким образом, по длине вариационного ряда можно судить о количестве деревьев на площади, а по его форме о структуре данной ценопопуляции. Кроме вариационных рядов, для каждого вида и каждой пробной площади построены частотные распределения деревьев в зависимости от их толщины (Ипатов, 1969). При этом количество ступеней толщины зависит от количества деревьев в выборке, для определения количества ступеней толщины использовалось следующее правило математической статистики: на каждую ступень толщины в выборке должно в среднем приходиться не менее 10 деревьев. Если в выборке 100 деревьев, то мы не можем использовать более 10 ступеней, если в выборке 200 деревьев, то количество ступеней толщины можно увеличить до 20 и т.д.
Представление о принципе плотнейшей упаковки особей в фитоценозах можно получить, привлекая идею о многомерном факторном пространстве. По осям этого пространства откладываются, кроме трех измерений реального географического пространства, также степень увлажнения, освещенность, содержание питательных веществ в почве и другие факторы среды. В ходе совместной эволюции экологически близких видов преимущество получали те из них, которые смогли – путем мутаций и отбора – свести до минимума конкурентный шум, исходящий от соседей по экологической нише. Достичь этого стало возможно, главным образом, путем «разбегания» особей в многомерном пространстве факторов (Пузаченко, 1976). Каждый вид осваивает свой объем в этом фазовом пространстве, и приспособление к специфическим условиям обитания закрепляется наследственным кодом. «Давление жизни» приводит к тому, что незаполненных промежутков между факторными осями по мере формирования сообщества становится все меньше. Экологическая система эволюционирует в направлении, предписанном принципом плотнейшей упаковки, но упаковки многомерных экологических ниш в многомерном экологическом пространстве. Каждый реальный фитоценоз с конкретным списком взаимодействующих видов и определенным набором условий среды представляет собой частный случай такой системы. Оценить количественно степень организованности системы в многомерном экологическом пространстве можно путем анализа распределений величин показателей связи в корреляционной матрице зависимостей между видами растений (Арманд, 1987).
В настоящее время наша система мониторинга растительного покрова на Дальнем Востоке и в Даурии насчитывает более 70 пп. В эту систему включены следующие ООПТ: Сихотэ-Алинский заповедник, заказник Борисовское плато, Хинганский заповедник, Сохондинский заповедник, Алханайский национальный парк, Буркальский заказник, а также модельные урочища: в Хасанском районе Приморского края, в окрестностях грязевого вулкана на юге острова Сахалин, в Алтано-Кыринской котловине в Забайкальском крае. В данной статье приводятся лишь наиболее значимые результаты, полученные нами в процессе мониторинга.
Результаты и обсуждение
Ландшафтный уровень
Результаты, полученные в результате мониторинга на постоянных пробных площадях в разных регионах Дальнего Востока оформлены в виде баз данных и обработаны статистически. В настоящей статье невозможно остановиться на всех выводах, которые были получены за 30 лет мониторинговых исследований, поэтому рассмотрим только самые существенные и наиболее интересные из них.
За период с 1982–1984 по 2006 гг. растительность на всех обследованных нами постоянных пробных площадях в Сохондинском заповеднике изменилась весьма существенно (табл. 1). Изменилось обилие ряда видов травянистых, кустарничков и кустарников. В верхнем лесном поясе появилось возобновление сосны обыкновенной (Pinus sylvestris), которое 20 лет назад здесь отсутствовало. В возобновлении почти во всех типах леса усилились позиции кедра сибирского (Pinus sibirica) и ослабли позиции лиственницы даурской (Larix dahurica). Среди кустарничков существенно снизилось обилие багульника болотного (Ledum palustre), ивы скальной (Salix saxatilis), голубики (Vaccinium uliginosum), но более активным видами стали: жимолость алтайская (Lonicera altaica), шиповник (Rosa acicularis) и некоторые другие. Существенно меньше в лесах стало осоки (Carex globularis), но увеличилось обилие другой осоки (Carex sp . из подрода vignea). За прошедший период заметно усилились позиции видов ксерофитов и мезо-ксерофитов (тяготеющих к сухим местообитаниям) и заметно ослабли ценотические позиции видов гигрофитов и мезогигрофитов. Это стало следствием устойчивого иссушения климата в последние 60 лет и значительного протаивания многолетней мерзлоты, в связи с чем понизился уровень залегания грунтовых вод.
Таблица 1
Сенсорные элементы в растительном покрове Сохондинского биосферного заповедника. (По А.В. Беликович, 2005).
№ № |
Краткое описание постоянных пробных площадей |
Изменения в видовом составе за 20 лет с 1982 по
2002 гг. (исчезнувшие виды + вновь появившиеся) |
1 |
2 |
3 |
1 |
Зарастание на шахтных отвалах рудника «Сохондо», закрытого в 1955 г. |
47 |
2 |
Кедово-лиственичный лес на склоне после ветровала в верхнем лесном поясе |
32 |
3 |
Лиственичный зеленомошный лес в нижней части склона зап. экспозиции в верхнем лесном поясе |
38 |
1 |
2 |
3 |
4 |
Редина с кустарниками на каменной россыпи в нижней части склона зап. экспозиции в верхнем лесном поясе |
36 |
5 |
Приручьевой кедрово-лиственичный лес в средней части склона зап. экспозиции в верхнем лесном поясе |
20 |
6 |
Кедрово-лиственичный лес в широкой лощине на пологом склоне зап. экспозиции в верхнем лесном поясе |
27 |
7 |
Возобновление на месте вырубки в верхней части пологого склона зап. экспозиции в верхнем лесном поясе |
39 |
8 |
Гарь на подгольцовой террасе |
58 |
9 |
Заросли кедрового стланника на уступе террасы в подгольцовом поясе |
14 |
10 |
Пятнистая нивальная кустарничково-лишайниковая тундра под уступом террасы в гольцовом поясе |
33 |
10а |
Осоково-разнотравная тундра на гольцовой террасе |
42 |
11 |
Куртинная кустарничково-лишайниковая тундра на вершине горы Сохондо |
44 |
12 |
Приручьевой кедровый с пихтой лес в субальпийском поясе в Букукунском каре |
34 |
13 |
Кустарничковая каменистая тундра с редкими стланниками в судальпийском поясе |
30 |
13а |
Сухая дриадовая тундра на уступе нагорной террасы в альпийском поясе |
50 |
14 |
Сырая осоково-пушицевая тундра на нагорной террасе в альпийском поясе |
52 |
15 |
Гарь на каменистой россыпи в подгольцовом поясе |
32 |
16 |
Возобновление по гари в кедрово-лиственичном лесу на моренной гряде в верхнем лесном поясе |
29 |
17 |
Приручьевое кедрово-лиственичное редколесье в верхнем лесном поясе |
29 |
18 |
Лиственично-кедровая редина на морене |
12 |
19 |
Прибрежная растительность моренного озера в верхнем лесном поясе |
19 |
20 |
Елово-лиственичный лес в ложбине стока в верхнем лесном поясе |
35 |
21 |
Ерниковый лиственично-кедровый лес на дне Джермалтай-Ингодинской депрессии в верхнем лесном поясе |
54 |
24 |
Пойменные кустарники с травяными прогалинами в долине реки Букукун в верхнем лесном поясе |
44 |
1 |
2 |
3 |
29 |
Остепненный луг на надпойменной террасе реки Агуца в нижнем лесном поясе |
29 |
30 |
Приручьевые кустарники ( Salix + Betula ) в долине ручья Бунинда в нижнем лесном поясе |
38 |
31 |
Горная разнотравная прерия с возобновлением осины в нижнем лесном поясе |
25 |
32 |
Разнотравные ерники на надпойменной террасе реки Агуца в нижнем лесном поясе |
36 |
35 |
Березово-лиственичный рододендроновый лес на террасе в нижней части склона в нижнем лесном поясе |
15 |
Примечание. В таблице выделены шрифтом те площади, на которых произошли наибольшие изменения видового состава, эти сообщества мы считаем сенсорными элементами в растительном покрове Сохондинского заповедника.
|
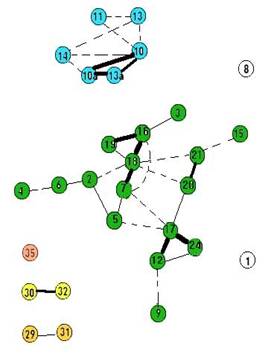
Рис. 1. Граф флористического сходства растительности постоянных пробных площадей в Сохондинском заповеднике в 1984 г. Кружки – вершины графа – постоянные пробные площади. Линии – ребра графа, их толщина пропорциональна уровню флористического сходства. (По И.В. Козырь, 2010)
|
Сравнение графов флористической структуры растительного покрова постоянных пробных площадей Сохондинского заповедника в 1984 и 2004 гг. (рис. 1, 2) позволяет оценить степень произошедших за 20 лет изменений в видовом составе пробных площадей. Всего заложено и подвергнуто ревизии в 2004 г. 29 пробны площадей ( n =29). При этом на графах 1984 и 2004 гг. количество значимых связей (когда R больше 0,2) равно 30 (А=30; В=30), количество совпадающих по положению связей на графах 1984 и 2004 гг. равно 20 (а=20). Мы можем для расчета сходства этих графов использовать индекс сходства Жаккара К=а/ A + B - a ). В нашем случае индекс Жаккара будет равен 0,5. Если учесть, что максимально возможная величина индекса Жаккара 1,0, а минимальная 0, можно заключить, что сравниваемые графы не очень сходны между собой, что флористическая структура растительного покрова Сохондинского заповедника на уровне мезокомбинаций растительности за 20 лет существенно изменилась.
Применение индекса Жаккара для расчета сходства графов не вполне корректно, так как множество ребер на наших графах существенно отличается от множества ребер полного графа при том же количестве вершин, когда каждая вершина с каждой связана ребром: N = M *( m -1)/2, где М – количество вершин графа. В нашем случае N = 29*28/2 = 406. В этом случае более подходящим для расчета сходства графов будет вероятностная, а не множественная мера, например, индекс сопряженности. Для этого мы должны оперировать не количествами событий, а вероятностями событий. Вероятность того что при случайном распределении ребер по вершинам конкретное ребро может занять одно определенное место на графе – связать именно эти две вершины – можно рассчитать как отношение 1/ N = 1/406. Вероятность совпадения двух случайных событий, в данном случае совпадения определенного ребра на графах 1984 и 2004 гг. равна произведению вероятностей (1/ N *1/ N ) = (1/ N ) 2 = 1/406*1/406 = 1/164836. В нашем случае ребер на том и другом графе по 30. Ожидаемое количество совпадающих по положению ребер на этих графах можно рассчитать по следующей формуле: а т = 30*30/406 = 2. Это значит, что при случайном распределении ребер на том и другом графе, совпадающих должно быть 2 ребра, а в нашем случае их 20. Следовательно, по теории вероятностей сходство двух наших графов весьма существенное и явно не случайное. Флористическая структура растительного покрова Сохондинского заповедника за 20 лет изменилась, но не настолько, чтобы перейти в свою противоположность. |
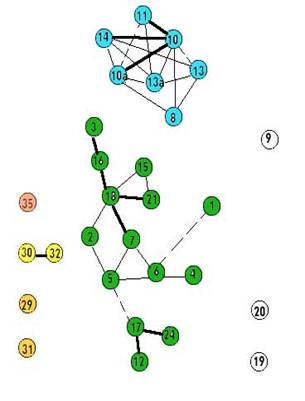
Рис. 2. Граф флористического сходства растительности постоянных пробных площадей в Сохондинском заповеднике в 2004 г. (По И.В. Козырь, 2010)
|
Графовые модели хорологической структуры растительного покрова можно использовать для оценки степени неравномерности структуры. В нашем случае неравномерность структуры графа отражает степень хорологического разнообразия растительного покрова Сохондинского заповедника. В равномерном графе, для которого характерно случайное распределение ребер по вершинам количество вершин с определенным числом исходящих из них ребер, можно рассчитать как произведение вероятностей элементарных событий – вероятность ребра занять определенное положение (Галанин, 1989). На рис. 3 изображены кривые статистических распределений количеств вершин графов в зависимости от количества исходящих из них ребер. Одна кривая получена для равномерного (случайное распределение ребер по вершинам) графа. Вторая кривая – это распределение числа вершин в зависимости от количества исходящих ребер в реальном графе. Степень несовпадения кривых случайного и реального графов можно оценить с помощью критерия Пирсона (Василевич, 1969). Величина критерия Пирсона для графа 1984 г. равна 56,6, а для графа 2004 г. только 9,0. Это говорит о существенном снижении хорологического разнообразия растительного покрова заповедника, произошедшего за 20 лет. Мы считаем, что такое снижение структурного разнообразия растительного покрова ландшафтного района есть явный признак природной катастрофы.
Графовые модели хорологической структуры растительного покрова можно использовать для оценки степени неравномерности структуры. В нашем случае неравномерность структуры графа отражает степень хорологического разнообразия растительного покрова Сохондинского заповедника. В равномерном графе, для которого характерно случайное распределение ребер по вершинам количество вершин с определенным числом исходящих из них ребер, можно рассчитать как произведение вероятностей элементарных событий – вероятность ребра занять определенное положение (Галанин, 1989). На рис. 3 изображены кривые статистических распределений количеств вершин графов в |
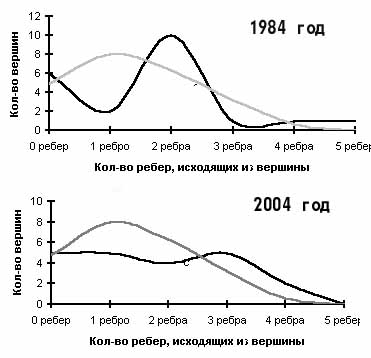
Рис. 3. Сравнение уровня хорологического флористического разнообразия растительности Сохондинского заповедника с помощью графовых моделей структуры растительного покрова в 1984 и 2004 гг.
|
зависимости от количества исходящих из них ребер. Одна кривая получена для равномерного (случайное распределение ребер по вершинам) графа. Вторая кривая – это распределение числа вершин в зависимости от количества исходящих ребер в реальном графе. Степень несовпадения кривых случайного и реального графов можно оценить с помощью критерия Пирсона (Василевич, 1969). Величина критерия Пирсона для графа 1984 г. равна 56,6, а для графа 2004 г. только 9,0. Это говорит о существенном снижении хорологического разнообразия растительного покрова заповедника, произошедшего за 20 лет. Мы считаем, что такое снижение структурного разнообразия растительного покрова ландшафтного района есть явный признак природной катастрофы.
Природная катастрофа – это такое неравновесное, нестационарное преобразование системы, следствием которого является потеря ею устойчивости (утрата динамического равновесия) в результате изменения собственных параметров и/или быстрого изменения внешней среды. На фото 1 и 2 можно видеть, насколько существенны изменения во внешней среде, произошедшие за 20 лет на территории Сохондинского заповедника.
Потепление привело к деградации многолетней мерзлоты, в результате чего понизился уровень грунтовых вод, дренировались многие озера на мировом водоразделе в верхнем лесном поясе, опустилось (просело) дно Букукунского озера, расположенного в ледниковом каре на высоте 1850 м н.у.м., причиной чего могли стать как тектонические движения земной коры, так и вытаивание погребенного под слоем обломков на дне озера реликтового ледника. Последнее, на наш взгляд, более вероятно, так как вода в Букукунском озере в июле стала прогреваться до +18 градусов, чего не наблюдалось 20 лет назад. |

Фото 1. Дренированное за 20 лет моренное озеро на Джермалтай-Ингодинской депрессии (1750 м н.у.м.). Произошло понижение уровня грунтовых вод в результате более глубокой протайки многолетней мерзлоты. В 1982–1984 гг. эта ложбина была непересыхающим озером. Фото А.В. Галанина
|

Фото 2. Оседание дна карового Букукунского озера за 20 лет. Сухие деревья в воде 30 лет назад были живыми и росли на берегу. Фото А.В. Галанина
|
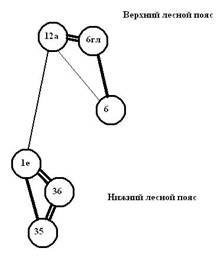 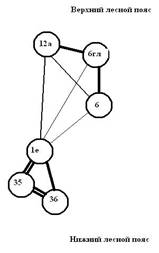
Рис. 4. Графы флористической ординации растительности 6 пп, заложенных в Сохондинском заповеднике: слева – при описании в 2000–2004 гг. и справа – при последующей ревизии в 2008–2010 гг. (По И.В. Козырь, 2010)
|
На рис. 4 приведены два графа флористического сходства растительности шести пробных площадей, один по результатам учета 2000–2004 гг., второй по данным учета 2008–2010 гг.
На графе первичного описания (2000–2004 гг.) наибольший уровень сходства (К от 0,15 до 0,16) наблюдается в кластере «нижний лесной пояс» среди пробных площадей нижнего лесного пояса пп-35, пп-36, а также между пп-35 и пробной площадью пп-1Е. Несколько меньший уровень сходства (К от 0,11 до 0,14) прослеживается в кластере «верхний лесной пояс» между пробными площадями верхнего лесного пп-6, пп-6Гл, а также между пп-6Гл и пробной площадью подгольцового пояса пп-12а. Эти 2 кластера оказываются связанными при уровне сходства (К от 0,07 до 0,10) через пробные площади пп-1Е, пп-12а (подгольцовый пояс) и пп-6Гл (верхний лесной пояс). Пробная площадь, заложенная в переходной зоне между нижним и верхним лесными поясами (пп-1Е), в большей степени сходна с лесной растительностью из нижнего лесного пояса, чем с растительностью верхнего лесного пояса. Наблюдаемое сходство пп-1Е с площадями верхнего лесного и подгольцового поясов (но уже в меньшей степени) объясняется присутствием на ее площади нескольких видов – представителей из верхнего лесного и подгольцового поясов: единичное возобновление Pinus sibirica, присутствие Bistorta vivipara, Valeriana turczaninowii, Viola biflora и др.
На графе (2008–2010 гг.) в кластере «нижний лесной пояс» уровень сходства повысился у пп-1Е с пп-35, но понизился у пп-1Е с пп-36; в кластере «верхний лесной пояс» понизился уровень сходства между пп-12а и пп-6гл. Кроме того, повысилось сходство между кластерами «нижний лесной пояс» и «верхний лесной пояс» – появились две значимые межкластерные связи. Такая перестройка флористического сходства между пробными площадями стала следствием проникновения ряда видов нижнего лесного пояса в верхний. Однако при этом субальпийский кедровый лес в Букукунском каре (пп-12а) несколько флористически «удалился» от верхнего лесного пояса. Таким образом, проведенное исследование позволяет сделать следующие общие выводы.
Ценотический уровень
Анализ структуры древостоев на постоянных пробных площадях позволил выявить явление ценотического маятника в двувидовых древостоях. Это явление отчетливо регистрируется на рис. 5, на котором приведены кривые частотных распределений особей кедра сибирского и лиственницы даурской в зависимости от толщины ствола на пробной площади в кедрово-лиственничном приручьевом лесу на пп-5 в Сохондинском заповеднике. На рис. 5 видно, что кривая кедра и кривая лиственницы находятся в противофазе относительно друг друга: там, где у кедра положительная деформация кривой, там, у лиственницы отрицательная и наоборот. Это можно объяснить тем, что кедр и лиственница активно возобновлялись в разные отрезки климатического цикла. В |
период, когда интенсивно возобновлялся кедр, лиственница возобновлялась плохо, и наоборот. Именно цикличность изменения климата является причиной того, что в древостое сосуществуют два вида, несколько отличающиеся друг от друга по требованиям к условиям внешней среды. По всей вероятности кедр активизировался здесь в более теплые фазы 11-ти летнего климатического цикла (11-летний цикл солнечной активности), а лиственница в более холодные фазы.
Мониторинг древостоя на постоянных пробных площадях позволяет выявить не только динамику ценопопуляций каждого вида, но и проследить развитие ценотических отношений между видами деревьев (рис. 6, 7).
Такие исследования были проведены нами в Сихотэ-Алинском заповеднике. Постоянные пробные площади здесь были заложены в конце 50-х – начале 60-х годов ХХ в. И.Н. Флягиной (1982). Древостой на них подвергался многократной ревизии, последняя ревизия была проведена нами в 1997–2000 гг. В качестве примера приведем результаты мониторинга на пп-М1, заложенной в 1967 г., во вторичном березовом лесу, возникшем после повального пожара на месте кедрово-широколиственного леса, случившегося здесь в начале 50-х годов ХХ в. (рис. 6).
На этом рисунке видно, как на пп изменялась численность и размерная структура ценопопуляции березы с 1967 по 1999 гг. Численность особей за этот период уменьшилась почти втрое. В 1967 и 1977 гг. ценопопуляция была неоднородна, отчетливо просматриваются две деформации кривых распределений с двумя модальными значениями толщина ствола, которые можно назвать «лидерами» и «угнетенными. Кривая 1987 г. имеет только одну четкую деформацию, а вот в 1992 г. деформации кривой частотного распределения березы незначительны. По нашему мнению, в период с 1987 по 1992 гг. береза в данном сообществе перестала быть лидером. Такие изменения в структуре ценопопуляции березы полностью соответствуют теории сукцессии. Пионерный вид постепенно теряет свои позиции в растительном сообществе, происходит смена пород.
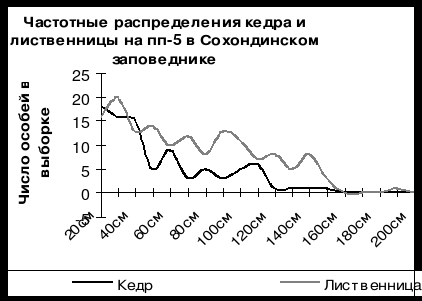
Рис. 5. Статистические распределения особей кедра сибирского и лиственницы даурской в зависимости от толщины ствола на постоянной пробной площади в кедрово-лиственничном лесу в Сохондинском заповеднике в 2001 г. По оси абсцисс – длина окружности ствола в см, по оси ординат – число особей на пп-5
|
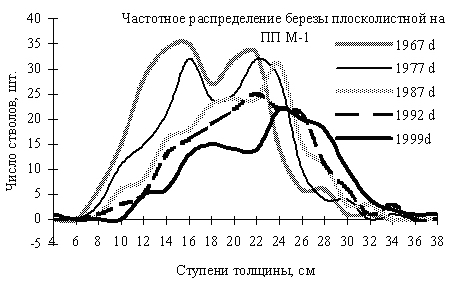
Рис. 6. Частотные распределения березы плосколистной в разные годы, по 2-х сантиметровым ступеням толщины в Сихотэ-Алинском заповеднике. По оси ординат – диаметр стволов в см, по оси абсцисс – число особей на пп. (По Н.А. Василенко, 2008)
|
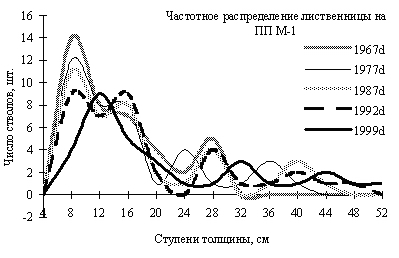
Рис. 7. Частотные распределения лиственницы даурской в разные годы, по 4-х сантиметровым ступеням толщины. По оси ординат – диаметр стволов в см, по оси абсцисс – число особей на пп. (По Н.А. Василенко, 2008) |
Кому же уступает свои позиции в фитоценозе по мере сукцессии береза на пп-М1? На рис. 7 приведены кривые частотного распределения особей лиственницы в зависимости от диаметра ствола. Видно, что в 1967 г. у ценопопуляции лиственницы было 2 четких деформации кривой и одна нечеткая, при этом преобладали тонкие особи. Однако уже тогда существовала немногочисленная группа толстых особей лиственницы, которые смогли пережить пожар в начале 50-х гг. В 1977 г. эти толстые деревья лиственницы резко увеличили диаметр с 28 см до 36 см, что хорошо видно на рис. 7. Однако из группы «средних» особей лиственницы с диаметром ствола 16 см к этому времени «вырвались» немногочисленные молодые особи сублидеры с диаметром ствола 24 см. Кривая распределения лиственницы 1987 г. существенно сместилась вправо, что говорит об интенсивном росте особей этого вида, при этом пропорционально росли в особи всех групп толщины, переходя при этом из одной группы в другую. Пополнение группы тонкомера с диаметром ствола 8 см к этому времени заметно снизилось, что можно объяснить только возросшей конкуренцией особей за ресурсы. В период с 1987 по 1992 гг. структура популяции лиственницы на пп-М1 изменилась мало, хотя количество особей тонкомера заметно сократилось. Очень существенная перестройка размерной структуры ценопопуляции этого вида произошла в период с 1992 по 1999 гг. Резко сократилось количество тонкомера в результате изреживания, зато сформировались две малочисленные группы «лидеров» с диаметром ствола 32 см и 44 см.
|
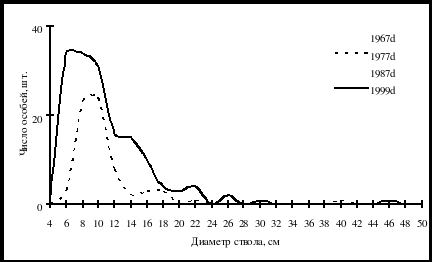
Рис. 8. Частотные распределения кедра корейского в березняке разнотравно-осоково-папоротниковом (пп М-1) на территории Сихотэ-Алинского биосферного заповедника. По оси ординат – диаметр стволов в см, по оси абсцисс – число особей на пп. (По Н.А. Василенко, 2008) |
На рис. 8. приведены кривые частотных распределений особей кедра корейского в зависимости от толщины ствола в разные годы на пп-М1. Все кривые имеют достоверные деформации, причем деформации в диапазонах толщины деревьев более 20 см по диаметру у кедра, как и у лиственницы, образуют деревья, выжившие после пожара, а деревья тоньше 20 см по диаметру – это послепожарное возобновление. У березы «допожарными» являются деревья с диаметром ствола больше 25 см, а все деревья тоньше относятся к послепожарной генерации. В отличие от лиственницы ценопопуляция кедра весьма интенсивно пополнялась в период с 1987 г. по 1999 г. Если до 1987 г. лидером в подросте была лиственница, то после 1987 г. активность в подросте явно перешла к кедру. По всей вероятности в 1877–1987 гг. кедр на пп-М1 сильнее лиственницы пострадал от низового пожара, но после 1990 г. он в группе подроста вновь стал лидировать.
Уже в 1967 г. пополнение ценопопуляции березы из подроста на пп-М1 прекратилось, все особи этого вида разделились на две совокупности: 1 – с модальным значением диаметра ствола 14–16 см и 2 – с модальным значением 21–23 см. Эти две совокупности образовались в результате внутривидовой конкуренции между особями березы. Толстые стали лидировать и вышли в верхний ярус, тонкие сформировали второй ярус. В 1977 г. обе совокупности сохранились, деревья, входящие в них выросли в толщину примерно на 1 см по диаметру. Количество особей в «тонкой» совокупности заметно сократилось, в «толстой» осталось практически прежним. В 1987 г. кривая |
распределения березы имела одну явную деформацию, при этом деформация совокупности «тонких» деревьев исчезла, – они не выдержали внутривидовой конкуренции. В 1992 г. исчезла и деформация совокупности «толстых» особей березы, кривая ее распределения приобрела колоколоподобный одновершинный вид. А вот в 1999 г. кривая вновь становится достоверно деформированной с двумя модальными значениями диаметра ствола – 17–19 см и 24–28 см. На этот раз дифференциация особей березы по толщине связана не только с внутривидовой конкуренцией, сколько с конкуренцией особей березы с особями лиственницы и кедра, которые стали заметно теснить березу.
Ценопопуляция лиственницы в 1967 г. имела две достоверные деформации с модальными значениями диаметра 8–9 см и 28 см и слабо выраженную недостоверную деформацию с модальным значением 15–16 см. Деформация «толстых» деревьев здесь связана с лиственницами допожарной генерации, которые смогли выжить после пожара. В 1977 г. кривая лиственницы имеет три достоверных деформации, причем модальные диаметры первых двух деформаций остались с 1967 г. неизменными, а третья деформация с модальным диаметром ствола 24 см явно возникла за счет опережающего роста части особей лиственницы, которые стали лидерами в конкуренции. Среди тонкомера лиственницы произошел незначительный отпад, но большая часть тонких особей перешла в класс большей толщины. В 1987 г. кривая распределения лиственницы имела четыре деформации с модальными значениями диаметра 9 см, 16 см, 28 см и 40 см. Две правые деформация образована «допожарными» особями, а вот три левых – это результат дифференциации уже «послепожарных» особей, которые разошлись по трем ярусам древостоя. В 1992 г. четыре деформации кривой распределения лиственницы сохранились с прежними модальными значениями толщины ствола. Это значит, что в период с 1987 по 1992 гг. лиственница росла не слишком интенсивно. Но за последующие 5 лет в период с 1992 по 1999 гг. в ценопопуляции лиственницы произошли существенные изменения. Ее кривая распределения за это время стала трехвершинной с явным сдвигом модальных значений диаметра вправо. Такая перестройка была связана с бурным ростом особей лиственницы в этот период.
Динамика ценопопуляции кедра корейского за период с 1967 по 1999 гг. чрезвычайно интересна. В 1967 г. на площади было много тонкомера кедра и незначительное количество особей с диаметром ствола больше 14 см. В 1977 г. количество особей кедра резко сократилось, что можно объяснить гибелью части их в результате низового пожара. Уцелевшие от пожара особи кедра за эти 10 лет несколько увеличили свой диаметр. В 1987 г. кривая распределения особей кедра заметно сдвинулась вправо, что можно объяснить только интенсивным ростом особей кедра и их активным внедревнием во второй ярус древостоя. К 1987 г. особи кедра и лиственницы в древостое становятся лидерами над особями березы. К 1999 г. продолжается интенсивный рост особей кедра в толщину, но самое главное – то, что к этому году ценопопуляция кедра существенно пополняется за счет возобновления и подроста, урон, нанесенный ценопопуляции кедра низовым пожаром, к этому времени полностью компенсирован.
Дальнейшую динамику древостоя на этой площади несложно предсказать. Будет происходить дальнейшая деградация ценопопуляции березы, прогрессировать ценопопуляция кедра, а лиственница будет входить в состав древостоя в качестве примеси. При восстановлении кедрово-широколиственного леса после верхового пожара в среднем Сихотэ-Алине стадия березового леса, в которой береза является доминантом, длится примерно 50 лет, за это время под пологом березового леса происходит возобновление лиственницы и кедра, которые через 60 лет после начала сукцессии становятся лидерами в древостое, а через 100 лет полностью вытесняют березу, кедр при этом постепенно теснит и лиственницу.
Самая длинная цепь ревизий древостоя на пп в нашем распоряжении имеется в Сихотэ-Алинском заповеднике. Пробная площадь пп-З4 заложена в 1953 г. И.Н. Флягиной (1982), ею проведены 3 ревизии: в 1963, 1973 и 1984 гг., последняя ревизия проведена нами в 1998 г. По результатам пяти таксаций древостоя нами были построены частотные распределения особей кедра корейского (Pinus korajensis), которые приведены на рис. 9. На этих диаграммах видно, как изменялась размерная структура ценопопуляции этого вида за почти полувековой период. В течение всего периода сохранялась многовершинность кривых распределений, если в 1953 г. кривая имела две достоверные положительные деформации с модальными значениями диаметра 8–12 см и 20–28 см, то в 1998 г. их было четыре с модальными значениями диаметра 8–10 см, 23–25 см, 31–35 см и 52 см, следовательно, гетерогенность размерной структуры данной ценопопуляции усилилась. Примечательно, что с 1953 г. по 1984 г. количество тонкомера закономерно снижалось, а к 1998 г. вдруг существенно увеличилось. Причиной этого была выборочная рубка нескольких самых толстых деревьев, что видно при сравнении кривых распределений 1973 и 1984 гг. По всей вероятности, это стало причиной существенного прироста у деревьев, диаметр которых в 1984 г. составлял 38–40 см, эти деревья в 1998 г. образовали деформацию кривой в области значений диаметра ствола 52 см. Всего за 5 лет система (древостой) «устранила» возникшую в результате выборочной рубки лакуну. Осветление полога в результате удаления самых мощных деревьев и освобождение ресурсов местообитания вызвало интенсивное пополнение тонкомера. Существенно сместились вправо положительные деформации кривой 1984 г. в области значений диаметра 20 см, 26–30 см. Таким образом, мы видим, как «чувствительна» ценотически хорошо сформированная система древостоя к внешним воздействиям и как быстро (всего в течение 5 лет) в условиях Приморского края она способна восстановить утраченное в результате воздействия равновесие.
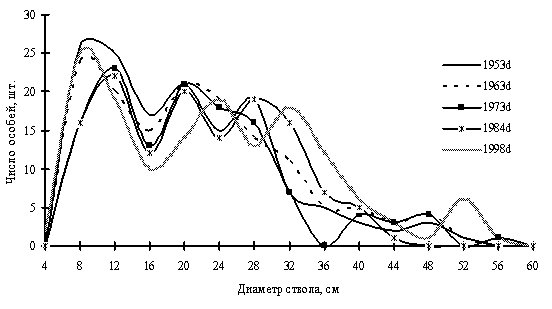
Рис. 9. Частотные распределения кедра корейского в кедрово-пихтовом (Pinus korajensis + Abies nephrolepis) с елью (Picea jesoensis) и дубом (Quercus mongolica) в мелкотравном лесу (пп-З4) на территории Сихотэ-Алинского биосферного заповедника. (По Н.А. Василенко, 2008) |
На рис. 10 и 11 приведены вариационные ряды лиственницы даурской и сосны обыкновенной на пп-А1 в Алханайском национальном парке в северной части Хэнтей-Чикойского нагорья в Читинской области. Хорошо видна разница в динамике ценопопуляций этих видов в лиственнично-сосновом лесу в Алханайском национальном парке в Забайкальском крае на пп-А1. Количество живых особей лиственницы в 2010 г. немного меньше особей сосны, но вот количество сухих особей сосны в этом году значительно больше, чем особей лиственницы. В каждом классе толщины особей лиственницы здесь меньше, чем особей сосны, но, тем не менее, отпад лиственницы «отстает» от отпада сосны весьма существенно. Это позволяет предполагать, что в данном типе леса началась постепенная смена лидера сосны обыкновенной лиственницей даурской.
|
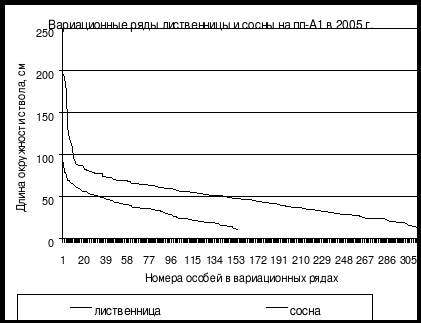
Рис. 10. Сравнение вариационных рядов лиственницы даурской и сосны обыкновеннной на пп-А1 в Алханайском национальном парке в лиственично-сосновом мертвопокровном лесу в 2005 г. (По Л.М. Долгалевой) |
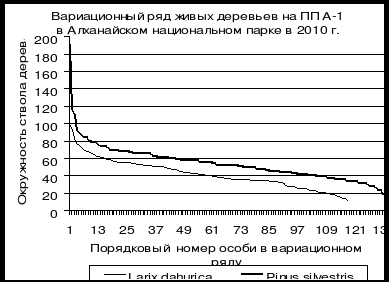
Рис. 11. Сравнение вариационных рядов лиственницы даурской и сосны обыкновеннной в Алханайском национальном парке в лиственично-сосновом мертвопокровном лесу в 2010 г. (По Л.М. Долгалевой) |
| |
Таблица 2
Динамика древостоя лиственично-соснового мертвопокровного леса в Алханайском национальном парке в 2005 и 2010 гг. (По Л.М. Долгалевой).
Динамика фаутности древостоя на ПП А-1 в Алханае |
Вид |
Деревьев в 2005 г. (всего учтенных 471) |
Деревьев в 2010 г. (всего учтенных 481) |
|
сухостойных |
живых |
фаутных |
|
сухостойных |
живых |
фаутных |
выпало |
Всего |
121 |
338 |
12 |
471 |
95 |
130 |
193 |
63 |
481 |
из них: |
|
|
|
|
|
|
|
|
|
Pinus sylvestris |
81 |
219 |
9 |
309 |
59 |
70 |
133 |
47 |
309 |
Larix dahurica |
38 |
113 |
3 |
154 |
29 |
58 |
60 |
16 |
163 |
% |
|
|
|
|
|
|
|
|
|
Pinus sylvestris |
26,3 |
70,8 |
2,9 |
100 |
19,1 |
22,6 |
43,1 |
15,2 |
100 |
Larix dahurica |
24,7 |
73,4 |
1,9 |
100 |
17,8 |
35,6 |
36,8 |
9,8 |
100 |
|

Фото 3. Закладка постоянной пробной площади пп-А1 в Алханайском национальном парке в 2005 г. Лиственично-сосновый редкотравный лес. Фото А.В. Галанина |
Проанализированные данные по структуре древостоя на пп-А1 показывают, что в результате произошедшего изменения климата увеличилась вероятность лесных пожаров, в результате которых более уязвимым видом оказывается сосна обыкновенная, которая в условиях аридизации климата уступает позиции в сообществах лиственнице даурской. Даже выжившие в результате пожара толстые особи сосны резко замедляют в росте и со временем погибают, уступая особям лиственницы в конкуренции за ресурсы воды и минеральных веществ. На фото 3 видно, в каком состоянии находится лиственично-сосновый лес на пп-А1
Проведенный анализ показал, что горизонтальная флористическая неоднородность лиственично-соснового рододендронового леса в нижнем лесном поясе в Сохондинском заповеднике на пп-35 очень неустойчива во времени. В 2001 г. при ревизии пп-35 она была разделена на 25 постоянных квадратов размером 10х10 м, было составлено подробное геоботаническое описание каждого квадрата, выявлены все виды сосудистых растений и составлена матрица распределения видов и основных синузий по квадратам с указанием их проективного покрытия. В 2008 г. при очередной ревизии перечет видов и синузий на этих квадратах был проведен снова, и получена вторая матрица. Были построены два графа (рис. 12) флористического сходства квадратов. За 7 лет распределение видов по квадратам 10х10 м в пределах пробной площади изменилось весьма существенно. На площади за 7 лет «исчезли» 13 видов, вновь зарегистрированы 14 видов, сохранились 30 видов. Были построены графы |
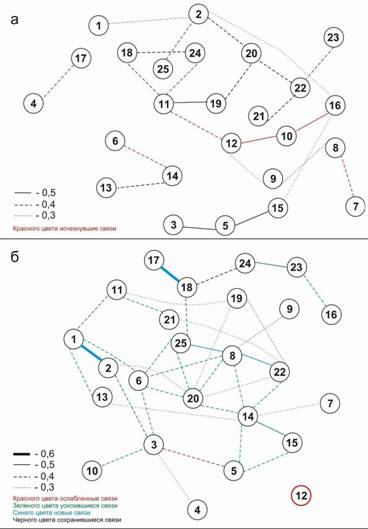
Рис. 12. Графы флористического сходства постоянных квадратов пробной площади пп-35 в 2001 г. (а) и в 2008 г. (б) (По И.В. Козырь, 2010) |
флористического сходства квадратов, сравнение которых показало, что сходство графа 2001 г. (а) и графа 2008 г. (б) по Жаккару составляет 52,6%. Случайность в распределении видов по квадратам в лесном фитоценозе столь велика, что ставит под сомнение использование видов травянистых растений в качестве индикаторов при флористической классификации лесных фитоценозов. Больше всего изменился флористический состав на квадратах.
Наши исследования показали, что в результате лесообразовательного процесса в лесном сообществе происходит не простое отставание в росте одних деревьев от других, а увеличение функционального разнообразия компонентов древостоя. В процессе конкуренции формируются естественные группы особей разной толщины, которые располагаются в разных ярусах древостоя и выполняют в сообществе биогеохимическую работу по-разному. Это хорошо иллюстрирует диаграмма на рис. 13.
Видно, что в данном фитоценозе интенсивно росли в толщину самые толстые и самые тонкие деревья, а деревья средней толщины и толстые росли значительно медленнее. Это можно объяснить только тем, что толстые деревья не выдерживали конкуренции с самыми толстыми, а деревья средней толщины не выдерживали конкуренции с тонкими. Ресурсов местообитания для тонких деревьев, чтобы они росли хорошо, хватало, но их было недостаточно для роста деревьев средней толщины. Следовательно, самые толстые и самые тонкие деревья на пп-12А занимают разные экологические ниши.
Морфологическая структура древостоя изучалась дополнительно методами кластерного анализа, отражающего ключевые «узлы» по толщине стволов, где идет дифференциация особей, занимающих одну экологическую нишу. Кластерный анализ позволяет классифицировать многомерные наблюдения, каждое из которых описывается набором исходных переменных x1, x2,…, xn. Преимущество кластерного анализа – это возможность разбивать совокупность на группы с учетом всех группировочных признаков одновременно. |
В кластерном анализе используется политетический подход как принцип образования групп. Все группировочные признаки одновременно участвуют в группировке, т. е. они учитываются все сразу при отнесении наблюдения в ту или иную группу. При этом, как правило, не указаны четкие границы каждой группы, а также неизвестно заранее, сколько же групп целесообразно выделить в исследуемой совокупности. Кластерные методы классификации как нельзя удачно подходят для целей мониторинга, которые связаны с изучением массовых явлений и процессов. Методы кластерного анализа позволяют решать задачи проведения классификации объектов с учетом признаков, отражающих сущность структуры изучаемой совокупности объектов, т.е. проводить поиск существующей структуры, прогноз изменений; строить новые классификации для слабодифференцированных ценозов, когда необходимо установить наличие связей в некой совокупности и начало процессов дифференцирования в экосистеме; унификацию данных в выборке с большим числом данных (рис. 14).
Рис. 13. Прирост деревьев кедра сибирского на пп-12А в субальпийском поясе Сохондинского биосферного заповедника разной исходной толщины за период с 1976 по 2000 гг. |
Рис. 14. Диаграмма кластеризации древостоя по обхвату стволов, мм на пп А-3 в национальном парке «Алханай» в 2006 г. |
В диаграмме кластеров показаны «расхождение» по обхвату ствола пород сходных экологических ниш. Первая группа – особи с толщиной более 382 мм (лиственница) и менее 137,5 мм – береза. Среднетолщинные группы, состоящие из лиственницы и ивы росистой – более 137, 5 разбиваются на 4 ветви более 315; 242,5; 222,5; 157,5 мм. Учитывая, что лиственница имеет длительный срок жизни, а ива – короткоживущий вид, прогноз развития ценоза определен последующим доминированием лиственницы. Анализируя фаутность и динамику структуру усыхания ивы росистой с помощью графика бокс-плот («ящик с усами»), прогноз динамики ценоза можно сделать с достаточной степенью вероятности (рис. 15).
Средняя линия – медиана показывает тенденцию развития совокупности. Выборка из фаутных особей имеет медиану ниже середины ящика, что показывает отмирание молодых деревьев. Ящик совокупности живых ив имеет медиану выше середины, что показывает на параллельные процессы изреживания среди молодых деревьев, и рост в толщину только отдельных деревьев. Такая же картина встречается в перестойных березовых и ивовых лесах с возобновлением от пня.
Нахождение «боксов» сухостойных и живых деревьев почти на одном уровне и короткие «усы» у живых деревьев свидетельствуют о том, что популяция не полностью восполняется подростом, а, следовательно, при дальнейшем изменении экологических условий сменится на лиственничник. Наши исследования показали, что распределение деревьев по толщине подчинено определенным принципам. В основе такого принципа – максимальная морфологическая дифференциация, способствующая равномерному распределению ресурсов влаги и питания.
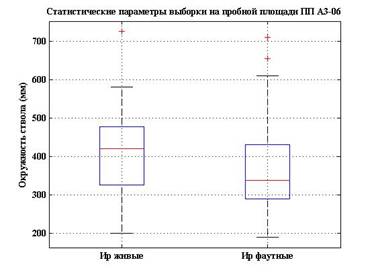
Рис.15. Бокс-плот совокупности данных по толщине для ивы росистой на пп А-3 в национальном парке «Алханай» в 2006 г.
|
При изменении экологических условий происходит сукцессия, в основе которой – смена доминантов. Так, на пп А-4 кедрово-лиственничный лес через 4 года уже заметны изменения. На пп А-4, заложенной в 2006 г. в национальном парке «Алханай» прослеживается изменение вариации живых особей – увеличение числа живых и переход в стадию «сухостой» крупных фаутных деревьев. Популяция лиственницы, судя по кривой вариационного ряда, незначительно изменилась морфологически (рис. 16). Живая лиственница в 2010 г. испытывает дефицит молоди (обрывистая кривая в последней трети вариации). Крупномерные особи перешли в статус фаутных, ослабив ценопопуляцию этого вида. Кривая сухих деревьев за 4 года практически не изменилась. Кедр, немногочисленный в 2006 г., сократил свое участие в ценозе в 2010 г. Доля фаутных особей возросла (рис. 17). Между тем, доля тонких деревьев недостаточна для пополнения популяции. Это говорит о том, что полноценный древесный ценоз сменится ксерофитным редкостойным лиственничником.
Изменения древостоя за 4 года обусловлены явлениями гидрологического и климатического характера. Климатические колебания: аномально жаркие летние месяцы и бесснежные зимы привели к иссушению Аршана, горного ручья снегового ледникового питания. Небольшая наледь, находящаяся в гроте с наветренной стороны ручья у каменной россыпи и питающая ручей, и каменный глетчер в толще каменной россыпи за 4 года растаял. Вода верховьев Аршана ушла под русло и лишь местами выходит на поверхность. Недостаток влаги вызвал смену растительного покрова: мохово-брусничные мезо- и гигрофитные участки пробной площади, стабилизирующие режим влажности в прикорневом слое, сменились мезо- и ксерофитными травяными сообществами. |
А
В
Рис. 16. Вариационные ряды лиственницы даурской (Л) в 2006 г. (А) и в 2010 г. (В) в Национальном парке «Алханай»
А

В
Рис. 17. Вариационные ряды кедра сибирского (К) в 2006 г. (А) и в 2010 г. (В) в Национальном парке «Алханай»
|
Такие сукцессионные процессы связаны с высокой экологической валентностью лиственницы. Лиственница даурская растёт в низинах, на заболоченных и торфянистых марях , вблизи неглубокого залегания многолетней мерзлоты , на каменистых россыпях и других довольно сухих местообитаниях. Проточное увлажнение неблагоприятно для нее. Лиственница – мезотроф, но в пределах своего ареала она широко распространена на территориях со сравнительно бедными почвами. Это и определяет сукцессионные процессы с заменой кедра на лиственницу. Поскольку дерево светолюбиво и пропускает свет, то это создает благоприятные условия для развития травяного покрова на месте мохово-брусничного. Поэтому в условиях ксерофикации биотопа лиственница становится доминантом в ценозе.
Кедр сибирский предпочитает супесчаные и суглинистые, средне увлажненные, дренированные плодородные почвы. На сухих песчаных и болотистых почвах он растет плохо (Игнатенко, 1988). Корневая система кедра требует аэрируемых почв. Кедр сибирский – микотрофное дерево. В условиях застойного переувлажнения на его корнях не развивается свойственная ему микориза, поэтому он здесь плохо растет. В условиях проточного увлажнения кедр хорошо идет в рост и неплохо возобновляется (Кирсанов, 1981). В горных условиях у деревьев формируется поверхностная корневая система. В период засухи кедр ослабевает, засыхает, а затем выпадает из древостоя. Поэтому засушливые годы способствуют усыханию кедра не только в нижнем лесном поясе, но и в высокогорных лесах.
Дифференциация деревьев в ходе их роста в сообществе на функциональные группы была изучена нами на пп-1Сах, заложенной в южной части острова Сахалин в лиственничнике (Larix cajanderi) бамбуковом (Sasa curilensis), посаженом в 30-е годы (рис. 14). На диаграмме частотного распределения особей лиственницы в зависимости от толщины видно, что в древостое сформировались две группы деревьев, между которыми имеет место достоверный хиатус (провал на диаграмме). Первую группу «лидеров» образуют особи с длиной окружности ствола 400 – 750 мм, вторую группу «аутсайдеров» особи с длиной окружности ствола 150 – 300 мм. Поскольку все деревья в посадке одного возраста, то дифференциация особей на две дискретные группы произошла в процессе их конкуренции за ресурсы местообитания. |

Фото 4. Постоянная пробная площадь пп-1Сах в окрестностях г. Южно-Сахалинска, заложенная в лиственничнике бамбуковом в 2005 г. Фото А.В. Галанина |
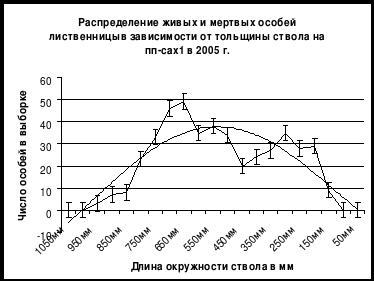
Рис. 18. Кривая частотного распределения особей лиственницы Каяндера в зависимости от длины окружности ствола на высоте 1,3 м. Дифференциация особей лиственницы на пп-1Сах по толщине ствола. Все особи одного возраста, они были высажены в 30-е годы ХХ века |
Рис. 19. Распределение толстых особей лиственницы Каяндера (условно подчиненных) в фитогенном поле толстых особей лиственницы Каяндера (условно лидеров) на пп-1Сах. Распределение получено методом геометрического суммирования. Учитывались только толстые особи лиственницы. Интегральная «действующая» особь лиственницы расположена в центре круга. Диаметр круга 5 м. (По Н.А. Василенко, 2008)
|
На диаграмме (рис. 15) хорошо видно, что при посадке все особи лиственницы на пп-1Сах были расположены друг от друга на расстоянии 1 м. Следует сказать, что, когда мы проводили закладку пп и ее описание, то мы не знали, что древостой на пп посаженный, настолько хорошо он был сформирован (фото 4). Однако построение круговых моделей распределения особей в фитогенном поле особи лидера методом геометрического суммирования (Зимов, Чупрынин, 1991; Василенко, 2008) выявило, что метровое удаление особей друг от друга здесь налицо. Обращают на себя внимание сгущения и разрежения точек (особей лиственницы) в фитогенном поле толстых особей лиственницы, полученные при геометрическом суммировании распределения особей в конкретных фитогенных полях конкретных особей лидеров. Достоверность неравномерности распределения точек в пределах круга доказана статистически при уровне значимости Р=0,5 (вероятность ошибки 5%). Такое суммированное распределение условно «подчиненных» особей лиственницы в фитогенном поле особей этого же вида условных «лидеров» есть следствие формирования в древостое регулярной структуры размещения деревьев в узлах шестиугольника вокруг особи «лидера». «Расталкивание» деревьев в лесу по мере их роста происходит в связи с конкуренцией за ресурсы местообитания, а вот формирование регулярного распределения по «закону шестиугольника» есть следствие оптимизации структуры фитоценоза, следствие его самоорганизации.
Мониторинг растительности и повторные описания древостоя на данной пп позволят выяснить, что происходит с распределением особей «лидеров» и «подчиненных» особей относительно друг друга во времени: становится ли распределение более регулярным и организованным, или в этом распределении со временем нарастает хаос. |
Изменения климата и растительности
Климат Даурии в плиоцене был значительно теплее современного, он был похож на современный климат северо-восточного Китая (Караваев, 1955). По данным М.Н. Алексеева (1978) в плиоцене на территории Восточного Забайкалья росли темнохвойные и хвойно-широколиственные леса с элементами тургайской флоры, а также ксерофильные и лесостепные ассоциации, площадь которых в ландшафте увеличивалась при движении к югу. В Маньчжурии в плиоцене господствовала неморально-субтропическая полуаридная растительность. Морская регрессия в плиоцене отодвинула морское побережье на восток, в связи с чем растения ксероморфного облика проникли даже в Приморье и Японию (Криштофович, 1958). В то время плиоценовые саванны и прерии были широко распространены в континентальной части Восточной Азии – в Маньчжурии и прилегающей к ней Даурии (Малышев, Пешкова, 1984). В плейстоцене произошло похолодание, несколько раз прерывалась связь Азии с Америкой через Берингийский мост суши. Климат и в плиоцене изменялся циклически (Величко, 1973). Однако в отложениях плейстоцена регулярно регистрируется пыльца таких представителей неморальной флоры, как дуб, ильм, липа, а некоторые авторы (Волкова, Белова, 1980) приводят эти деревья и для голоценовых отложений Забайкалья. Плейстоценовые оледенения в Даурии не были слишком мощными и носили либо каровый, либо горно-долинный характер. В сухие эпохи потеплений растительность Даурии носила степной или даже полупустынный характер, в это время здесь были широко распространены сообщества с доминированием эфедры (Ephedra dahurica, E. fruticosa, E. hinensis, E. monosperma).
Последняя вюрмская ледниковая эпоха закончилась 11 тыс. лет назад, начался очередной межледниковый голоценовый период. Но и в течение голоцена климат Земли не был постоянным, теплые засушливые периоды были во 2-м тысячелетии до новой эры и в 1-м тысячелетии новой эры, а в промежутке между ними было тысячелетие более холодного и влажного климата (Кобышева, Костин, Струнникова, 1980). В начале 20-го столетия началось потепление вод Арктики, которое было обнаружено в 1919 г. С той поры происходило уменьшение количества льдов в Северном ледовитом океане – процесс, который мы наблюдаем и в начале XX века. Совершенно точно установлено, что в последние 60–70 лет в самых разных частях Земли происходит деградация материкового оледенения. Сокращаются ледники в Скандинавии, в Альпах, в Аргентинских Андах, на Памире и Тянь-Шане, на Алтае, на Кавказе, на хребте Черского в Восточной Сибири, в Гренландии и других арктических островах, в Антарктиде (Поповкин, 2006). Данные метеорологических наблюдений показывают, что в последние 2 десятилетия аридность климата Даурии усилилась, произошло иссушение Гоби. Так, например, в Долине озер в Монголии с 1981 г. по 1990 г. сумма годовых осадков снизилась с 105,9 мм до 67,5 мм, а на южном макросклоне Хангая с 505,7 мм до 152,2 мм (Береснева, 2006).
Атлантический воздух весной и в первой половине лета приносит на европейскую и западноазиатскую части Евразии существенные осадки, однако в Даурию он приходит уже весьма сухим, и осадки здесь в это время редки и малообильны. Однако этот сухой атлантический воздух препятствует проникновению в Даурию весной и в начале лета влажных тихоокеанских воздушных масс. В Даурии, над которой располагается центральная часть Сибирского антициклона, зимой снежный покров очень маломощный, а местами снег вообще сдувается ветром. При длительных сильных морозах почва промерзает на несколько метров в глубину. На дне широких заболоченных речных долин имеется многолетняя мерзлота, по всей вероятности, реликтового характера. Весной над Даурией сухой холодный воздух держится до середины мая, в конце апреля среднесуточная температура здесь переходит через 0 градусов, относительная влажность воздуха в это время днем падает так низко, как в пустынях Средней Азии. В это время дуют сильные сухие ветры. Весна короткая, сухая и ветреная, что способствует возникновению и быстрому распространению лесных пожаров. В летнее время тихоокеанские воздушные массы проникают гораздо дальше на запад, однако в летнее время и арктические воздушные массы проникают в Восточную Сибирь и в Даурию. Благодаря этому в суммарном количестве выпадающих осадков доля летних циклонов составляет 70%, в Даурии это в основном тихоокеанская вода в виде дождей. Лето короткое, в июне бывают снегопады и заморозки. Вторая половина лета – с затяжными дождями, которые вызывают паводки.
Осенью уже в октябре в Даурии происходит перестройка полей давления и циркуляции, и над всей ее территорией устанавливается область повышенного давления. Сентябрьские дожди промачивают почву, питают грунтовые воды, и эта влага обеспечивает жизнь растений весной и в начале лета. Переход среднесуточной температуры через 0 градусов происходит в начале октября, а в конце октября и днем устанавливается отрицательная температура. Осень теплая и сухая, первые заморозки приходятся на конец августа – начало сентября.
В аридные эпохи в Даурии происходило опустынивание, в Маньчжурии прерии наступали на дубовые, сосновые и черноберезовые (Betula dahurica) леса, в это время северная граница пустыни Гоби проходила по югу теперешнего Забайкальского края. Маньчжурская прерия в аридные эпохи доходила до озера Ханка, а в Хасанском районе отдельные ее фрагменты встречались вплоть до побережья Японского моря. В такие эпохи здесь активизировались сосна густоцветковая (Pinus sylvestris ssp. densiflora), дуб зубчатый (Quercus dentata), ковыль байкальский (Stipa baicalense), нителистник сибирский (Filifolium sibiricum), володушка скорзонеролистная (Bupleurum scorzonerifolium), реликтовые популяции которых встречаются в Приханковье, в горах, на побережьи Японского моря и в настоящее время. Колебания климата между аридностью и гумидностью в пределах Восточно-Азиатской области, вероятно, приводили к значительно большим перестройкам в растительном покрове, чем потепления и похолодания. Важным фактором в динамике флоры и растительности на востоке области стало изменение уровня моря и связанные с этим осушения шельфовой зоны между Сахалином и материком, а также между Сахалином и Хоккайдо. Эти мосты суши способствовали миграции видов и их сообществ с материка на Сахалин, с Сахалина на Хоккайдо и Южные Курилы. Встречные миграции играли меньшую роль, так как материковые популяции видов, занимающие обширные территории, имеют более высокое генетическое разнообразие, их генофонд богаче, и это позволяет им быть ценотически более «агрессивными».
Нами получены данные, которые позволяют предположить, что в голоцене климат Даурии изменялся циклически с периодом в 600 лет (Галанин и др., 2005). В течение этого периода 200 лет бывает очень сухо и 200 лет весьма влажно, еще 100 лет приходится на переход от сухости к влажности, и 100 лет, наоборот, от влажности к сухости. В сухую часть климатического цикла степь наступает на леса и луга, происходит остепнение даже в тундровых сообществах на гольцах Хэнтея. Возможно, реликтом такой аридной эпохи являются песчаные дюны Чарской котловины на севере Читинской области (фото 5). Степные виды растений в эти эпохи внедряются на луга и в леса. В это время учащаются лесные и степные пожары, они становятся более опустошительными, так как в сухом климате процессы гниения замедляются, в лесу и степи накапливается много сухого материала. Дожди же в сухие эпохи становятся более редкими и мало обильными и не могут потушить бушующие лесные и степные пожары (фото 6). С подобными явлениями в последние годы столкнулись не только жители Даурии, но и Маньчжурии и Приморского края.
Один из важных результатов наших геоботанических и флористических исследований – это вывод об изменении растительного покрова юга Читинской области в связи с устойчивым иссушением климата. Последний раз длительная засуха в Центральной Азии разразилась в конце Х II и в Х III вв. В предшествовавшую этой длительной засухе влажную эпоху на продуктивных пастбищах в Даурии размножились стада домашних животных, увеличилась численность населения. В Х II в. климат резко ухудшился, продуктивность пастбищ снизилась, начался падеж скота. Именно бескормица толкнула монголов и татар под предводительством Чингисхана, а затем и его сыновей начать экспансию в регионы Восточной и Западной Азии и в Европу – в районы с более влажным климатом (Галанин и др., 2008).
Последний влажный климатический период в Центральной Азии был в Х VII и XVIII вв. В этот период на гольце Сохондо побывал участник ботанической экспедиции Петра Симона Палласа Никита Петрович Соколов. Он оставил нам довольно подробное и точное описание растительности района Сохондо в то время. Сравнивая его описания с состоянием растительности сегодня, мы можем однозначно сказать, что верхняя граница леса тогда проходила на 100–150 м выше, а нижняя на 100–150 м ниже, чем сегодня. Луга тогда занимали участки, на которых сегодня растет только степная растительность. И не случайно, что именно в это время была столь интенсивная экспансия русских в Сибирь и в Ононскую Даурию в частности. Климат здесь тогда позволял заниматься земледелием, на лугах можно было ежегодно заготавливать много сена.

Фото 5. Пески в долине р. Чара на севере Забайкальского края. Колебания климата в Забайкалье в прошлом были столь мощными, что пустынные «гобийские» ландшафты в аридные эпохи формировались даже здесь. В последние десятилетия здесь вновь происходит активизация песков. (Фото А. Леснянского, интернет-ресурс по адресу: http://www.photosight.ru/photos/870164/) |

Фото 6. Лесной пожар в окрестностях с. Букукун на юге Читинской области в 2008 г. Аридизация климата способствует возникновению лесных пожаров. Фото И. Маврина |
В XX веке в Центральной Азии в связи с глобальным изменением климата прогрессировало иссушение климата, особенно заметно оно стало в конце этого века и в начале ХХ I столетия. При этом среднегодовая температура воздуха с 1936 г. по 2010 г. увеличивалась с -3,5 до -1,5 градуса (рис. 16). При этом среднесуточное среднегодовое количество осадков изменялось нелинейно, что хорошо видно на полиномиальной линии тренда (рис. 17). На этой диаграмме видны два 25–26-летних цикла изменения среднегодового количества осадков. Минимальное количество осадков в этих циклах имело место в 1946–1947 гг., 1975–1976 гг. и в 2006–2007 гг., а максимальные – в 1961–1964 гг., в 1987–1990 гг. При этом за период с 1949 по 1979 гг. осадков в среднем за год выпадало больше, чем за период с 1980 по 2010 гг. (рис. 18). Если учесть, что за это время происходило повышение среднегодовой температуры воздуха, то станет понятно, что испарение с поверхности почвы увеличилось, что и вызвало усиление аридизации климата, что катастрофически отразилось на растительном покрове.
Рис. 20. Динамика среднегодовых среднесуточных температур воздуха в периоды с 1936 по 2010 гг. по данным ГМС «Букукун» (юг Читинской области) с линией полиномиального линейного тренда. (Материалы И. Маврина) |
Рис. 21. Динамика количества осадков за период с 1949 по 2010 гг. по данным ГМС «Букукун» (юг Читинской области). Гладкая линия – тренд, полученный методом экстраполяции с помощью полинома 6-й степени. (Материалы И. Маврина) |
Но произошло не простое равномерное уменьшение: по месяцам уменьшалось количество осадков в регионе, изменялся характер их распределения по месяцам и декадам в течение года, – изменился погодный ритм. Это хорошо видно на рис. 19. Произошло не просто снижение количества осадков, выпадающих в течение года, а стали значительно суше весна и первая половина лета, суше стали июль и август. А вот в июне осадков стало выпадать немного больше. Сложившаяся ситуация отрицательно сказалась как на видах растений степного склада, так и на видах мезофитах. Весной и в начале лета вероятность возникновения лесных и степных пожаров резко увеличилась.
Похоже, что не только для земледельцев, но и для кочевников-скотоводов здесь настали совсем не лучшие времена. Как долго продлится очередной сухой период? По нашему мнению, он продлится не более 100, но и не менее 50 лет. При этом в течение 2015–2018 гг. количество осадков увеличится, но затем последует еще более сухой период, чем период 1972–1982 гг., после чего гумидность климата снова немного возрастет.
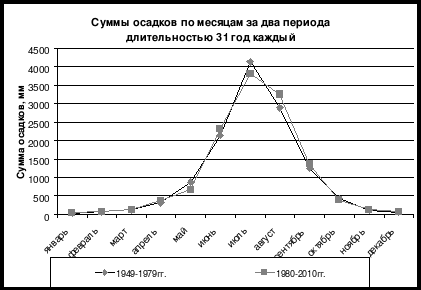
Рис. 22. Сравнение количества осадков по месяцам за два периода наблюдений по данным ГМС «Букукун». (Материалы И. Маврина) |
Рис. 23. Среднедекадное количество осадков за два периода по многолетним данным ГМС «Букукун» (юг Читинской области). 1 – за период с 1949 по 2010 гг.; 2 – за период с 1980 по 2010 гг. На диаграмме видны два периода: 1– засушливый (сухое лето) и 2 – влажный (сырое лето). (Материалы И. Маврина)
|
Как же пережить в Даурии эпоху современного иссушения климата? Что делать жителям края? Вопросов много, и на большинство из них у нас нет ответа. Ясно одно – надо прекратить распашку степи, и отказаться от земледелия, надо усиленно бороться с лесными и степными пожарами, необходимо прекратить устраивать весенние палы, ужесточить наказания за поджоги, в пожароопасные периоды запретить выезд на пикники, охоту и сбор грибов и ягод. Только лес может существенно смягчить последствия иссушения климата. Ни в коей мере нельзя допускать перевыпас скота и пастбищную дигрессию степных растительных сообществ.
О том, что опустынивание в прошлом в Даурии случалось, говорят гранитные глыбы в окрестностях Тыринских озер и в окрестностях с. Шумунда на юге Забайкальского края в Кыринском районе, На них видны следы былой мощной ветровой эрозии – выдува (фото 7). Такая эрозия характерна для пустынь. Значит, когда-то в этих местах задернение почвы было таким же слабым, какое мы сегодня видим в пустынных регионах Монголии (фото 8). Тогда существенным фактором формирования рельефа в Даурии были эоловые процессы.

Фото 7. Тыринское озеро на юге Читинской области. На гранитных скалах видны следы ветровой эрозии. Когда-то климат здесь был более аридный. Фото А.В. Галанина
|

Фото 8. Пустыня Гоби отвоевывает у Даурии все новые и новые территории. Фото И.А. Галаниной
|
Даурская антилопа – дзерен – в гумидные периоды климатического цикла обитала в степях и прериях северо-восточной Монголии, но в эпохи аридизации климата, когда степи и прерии Монголии превращались в пустыню, дзерены мигрировали на север. Даже в сильно потравленных сегодня степях Монголии дзерены находят для себя пропитание, но в последние 20 лет от бескормицы они мигрируют на юг Забайкальского края, в российской части Даурии орнитологи в последние 20 лет все чаще отмечают грифов. Нам известны заходы дзеренов в Сохондинский и Даурский биосферные заповедники, а популяция грифов постоянно обитает в Алтано-Кыринской котловине.
Эти явления, несомненно, связаны с длительной засухой, степными пожарами, с деградацией пастбищ от засухи и с перевыпаса домашнего скота в Монголии. Численность домашнего скота в Монголии в настоящее время регулируется только естественным путем, а именно, падежом его в конце зимы и весной от бескормицы. Никаких нормативных актов на предельно допустимое количество скота на единицу площади пастбищных угодий в Монголии нет.
Ландшафт северо-восточной Монголии – это сухая выбитая от перевыпаса степь-прерия (фото 9). Всего в 100 км к югу от Улан-Батора растительность в виде сухой кустарниковой карагановой (Caragana microphylla) степи с явными элементами опустынивания. Карагана мелколистная в условиях перевыпаса и аридизации климата резко усиливает ценотические позиции в растительном покрове. Таким образом, Caragana microphylla в Даурии – это защитный механизм степных экосистем, обеспечивающий сохранение их биоразнообразия.
Для Даурии характерен своеобразный ритм погоды, здесь уместно выделять не четыре, как обычно, а пять фенологических сезонов года. Это первая половина лета (с 20 апреля по 20 июля), отличающаяся крайней сухостью, особый сезон года – сухое лето; вторая половина лета (с 20 июля по 10 сентября) – период, когда количество осадков значительно и достаточно для роста и развития мезофитов, также особый сезон года – влажное лето. Подобный ритм погоды сохраняется в Даурии на протяжении многих десятков тысяч (а может быть, и сотен) лет благодаря тихоокеанскому муссону. За столь длительный период в здешних ландшафтах сформировались, по крайней мере, два своеобразных типа растительных сообществ, которым нет аналогов в других регионах северной Евразии – даурская прерия и харганат (Галани, Беликович, 2005). Даурская прерия представляет собой оригинальное сочетание настоящих травянистых ксерофитов, мезофитов и мезо-ксерофитов. В период сухого лета в экосистеме активны ксерофиты, рост и развитие которых осуществляется за счет запаса питательных веществ, накопленных в корневищах. В период влажного лета сильно активизируются мезофиты. Смесь ксерофитов и мезофитов в одном сообществе на одном участке – характерная особенность даурской прерии. В мае – июне эта прерия выглядит как сухая степь или даже полупустыня. Облик этой прерии в начале лета бурый, зелень едва пробивается и плохо видна среди прошлогодней ветоши, в это время цветут растения ксерофиты, но в конце июля – августе даурская прерия превращается в цветущий луг – активизируются растения мезофиты.

Фото 9. Типичный ландшафт в 100 км южнее Улан-Батора. Карагановая (Caragana microphylla) степь. Вдали убегающее стадо дзеренов. Фото А.В. Галанина |
Нестабильность погодных условий от года к году и вековые колебания аридности-гумидности климата в Центральной Азии приводят к тому, что процессы конкуренции никогда не успевают привести к вытеснению из растительных сообществ многих видов, как ксерофитов, так и мезофитов, при этом сообщества остаются многовидовыми. Действительно, в сырые годы конкурентно способными оказываются растения мезофиты, а в сухие – растения ксерофиты. Высокое таксономическое разнообразие даурской прерии говорит о том, что условия здесь в высшей мере нестабильны на протяжении длительного времени.
При мониторинге растительности следует выявлять ведущие факторы, вызывающие периодические изменения конкретных растительных сообществ. Например, при потеплении и явной аридизации климата в Даурии за последние 40 лет, в некоторых экосистемах верхнего лесного пояса в поймах рек влажность почвы резко возросла за счет более интенсивной оттайки многолетней мерзлоты в гольцовом и подгольцовом поясах. В альпийском поясе Хентея в результате такого потепления многие относительно холодолюбивые виды стали деградировать, так как снега стало выпадать меньше, и снеговое укрытие в местах перегибов склонов, где перезимовывали эти растения, стало незначительным. Стали гибнуть кусты кедрового стланика, сокращаться реликтовые ценопопуляции пихты сибирской. При климатических пертурбациях растительность на разных местообитаниях изменяется с разной скоростью и не синхронно. |
Литература
• Алексеева М.Н. Антропоген Восточной Азии. – М.: Наука, 1978. – 207 с.
• Арманд А.Д. Ландшафтная граница как объект мониторинга // Проблемы экологического мониторинга и моделирование экосистем. – Л.: Гидромстсоиздат, 1987. – С. 36–46.
• Беликович А.В. Сенсорные элементы в растительном покрове и их использование при выявлении природных катастроф // Ритмы и катастрофы в растительном покрове Дальнего Востока. – Владивосток: БСИ ДВО РАН, 2005. – С. 142–150.
• Беликович А.В. Ландшафтная флористическая неоднородность растительного покрова. – Владивосток: БПИ ДВО РАН, 2001. – 248 с.
• Беликович А.В., Галанин А.В. Забайкальский харганат как тип растительности // Комаровские чтения. – Владивосток: Дальнаука, 2005. – Вып.52. – С. 98–126.
• Береснева И.А. Климаты Аридной зоны Азии. – М.: Наука, 2006. – 286 с.
• Василевич В.И. Статистические методы в геоботанике. – Л.: Наука, 1969. – 232 с.
• Василенко Н.А. Самоорганизация древесных ценозов. – Владивосток: Дальнаука, 2008. – 171 с.
• Величко А.А. Природный процесс в плейстоцене. – М.: Наука, 1973. – 256 с.
• Волкова В.С., Белова В.А. О роли широколиственных пород в растительности голоцена Сибири // Палинология Сибири. – М.: Наука, 1980. – С. 112–117.
• Галанин А.В. Экологический потенциал флоры Полярного Урала и его использование в народном хозяйстве // Биологические проблемы Севера: Тез. Докл. 9-го симпозиума. 1981. Ч. 1. С. 18.
• ГаланинА.В. Перспективы ботанических исследований в Сохондинском заповеднике. // Ботанические исследования в заповедниках. РСФСР. – М., 1984. – С. 39–45.
• Галанин А.В. Анализ графов флористической структуры растительного покрова двух ландшафтных районов Чукотки // Метод графов в экологии / Биологические проблемы Севера. – Владивосток: ДВО РАН, 1989. – С. 6–24.
• Галанин А.В. Флора и ландшафтно-экологическая структура растительного покрова. – Владивосток: ДВО АН СССР, 1991. – 272 с.
• Галанин А.В. Флора и ландшафтно-экологическая структура растительного покрова. – Владивосток: Дальнаука, 2005. – 272 с. Режим доступа: http://geobotany.narod.ru/galanin/m1.htm
• Галанин А.В. Ландшафтно-экологический мониторинг на стационаре «Контакт» (Верхнеколымское нагорье) // Комплексные экологические исследования на стационаре «Контакт». – Владивосток: Дальнаука, 1993. – С. 3–19.
• Галанин А.В. Лекции по экологии / Мониторинг растительного покрова: состояние проблемы, основные понятия и элементы теории. Интернетресурс. 2006. Адрес доступа: http://www.botsad.ru/papers5.htm
• Галанин А.В., Беликович А.В., Галанина И.А., Роенко Е.Н., Яковченко Л.С. Растительность Даурии и великие переселения народов // Вестник СВНЦ ДВО РАН, 2008. – № 4. – С. 41–53.
• Галанин А.В. Некоторые мысли о биосфере и растительном покрове // Бюллетень Ботанического сада-института ДВО РАН, 2009. Вып. 4. Режим доступа: http:// botsad.ru/journal/number 4/4-18.pdf
• Галанин А.В., А.В. Беликович. Эколого-флористическая структура растительного покрова Сохондинского заповедника // Теоретико-графовые методы в биогеографии. – Владивосток: ДВО АН СССР, 1988.
• Галанин А.В., А.В. Беликович. Динамика древостоя в субальпийском кедровнике в Сохондинском заповеднике за 24 года // Классификация и динамика лесов Дальнего Востока. – Владивосток: Дальнаука, 2001.
• Галанин А.В., А.В. Беликович. Изменения в растительном покрове Сохондинского заповедника по результатам ревизии геоботанических пробных площадей (1983–2001) // Растительный и животный мир Сохондинского биосферного заповедника. Чита-Владивосток: БПИ ДВО РАН, БСИ ДВО РАН, СГБЗ, 2002. – С. 14–33.
• Галанин А.В., Беликович А.В. Даурия как подобласть Манчжурской ботанико-географической области // Комаровские чтения. – Владивосток: Дальнаука, 2006. Вып. LIII . – С.7–38.
• Галанин А.В., Беликович А.В., Галанина И.А. Отражение векового климатического ритма в растительности охраняемых территорий российского Дальнего Востока. (На англ. яз.) // Доклады международного APN-START Симпозиума по изучению глобальных изменений в Северо-Восточной Азии. – Владивосток: Дальнаука, 2005. – С. 25–36.
• Галанина И.А. Анализ распределения видов лишайников по постоянным пробным площадям в Сихотэ-Алинском заповеднике // Комаровские чтения. – Владивосток, 2005. Вып. 56. – С. 134–142.
• Галанина И.А. Синузии эпифитных лишайников в дубовых лесах юга Приморского края. – Владивосток: Дальнаука, 2008. – 238 с.
• Зимов С.А.Б Чупрынин В.И. Экосистемы: устойчивость, конкуренция, целенаправленное преобразование. – М.: Наука, 1991. – 159 с.
• Игнатенко, М.М. Сибирский кедр/ М.М. Игнатенко. – М.: Наука, 1988. – 162 с Ипатов B.C. Дифференциация древостоя II // Вести. ЛГУ. Биология. 1969. – Вып. З. – С. 43–54.
• Караваев М.Н. Новые находки серого ореха ( JuglanscinereaL .) в древних аллювиальных отложениях бассейна Алдана // Вопросы геологии Азии. Вып. 2. – М.: Изд-во АН СССР, 1955. – С. 815–823.
• Кирсанов В.А. Биолого-экологическая характеристика кедра сибирского как главного лесообразователя кедровых лесов / В.А. Кирсанов // Воспроизводство кедровых лесов на Урале и в Западной Сибири. – Свердловск: УНЦ АН СССР, 1981. – С. 3 –12.
• Кобышева Н.В., Костин С.И., Струнникова Э.А., Климатология. – Л.: Гидрометеоиздат, 1980. – 344 с.
• Кожевников Ю.П. Геосистемные аспекты растительного покрова Чукотки. – Владивосток: ДВО АН СССР, 1989. – 308 с.
• Козырь И.В. Мониторинг лесной растительности Сохондинского биосферного заповедника / Автореф. дисс. канд. биол. наук. – Владивосток, 2010. – 18 с.
• Криштофович А.Н. Происхождение Ангарской суши // Материалы по истории флоры и растительности СССР. – М.–Л.: Изд-во АН СССР, 1958. – С. 7–41.
• Любищев А.А. Проблемы формы, систематики и эволюции организмов. – М.: Наука, 1982. – 280 с.
• Мазуренко М.Т., Хохряков А.П. Структура и морфогенез кустарников. – М., Наука, 1977, – 160 с.
• МалышевЛ.И., Пешкова Г.А. Особенности и генезис флоры Сибири. Предбайкалье и Забайкалье. – Новосибирск: Наука, 1984. – 265с.
• Миркин Б.М. Теоретические основы современной фитоценологии. – М.: Наука, 1985. – 137с.
• Норин Б.Н. Фитоценотическая организация некоторых типов растительного покрова Крайнего Севера // Ботан. Журн. – 1980. – Т. 65, N 11. – С. 1531–1542.
• Норин Б.Н. Некоторые вопросы теории фитоценологии, ценотическая система, ценотические отношения, фитогенное поле // Ботан. Журн. – 1987. – Т.72, № 9. – С. 1116 –1174.
• Паллас П.-С. Путешествие по разным провинциям Российского государства. – СПб., 1773 – 1788.
• Пузаченко Ю.Г. Принципы информационного анализа // Статистические методы исследования геосистем. – Владивосток, 1976. – C . 5–37.
• Поповкин В.В. Современная эволюция ледников Земли // Современные глобальные изменения природной среды. Т. 1. – М.: Научный мир, 2006. – С. 507–577.
• Рафес П. М. О влиянии листогрызущих насекомых на биологический круговорот в лесах // Средообразующая деятельность животных. – М.: Наука, 1980. – С. 43–44.
• Флягина И.А. Лесовозобновление в кедровых лесах на восточных склонах Сихотэ-Алиня. – Владивосток: ДВНЦАНСССР, 1982. – 180 с.
• Breymeyer A.I. Ecosiste homeostatic-search for a definition // Mem. Zool. 1979. Vol. 32. N 1. P. 3-11.
• Galanin A.V., N.A. Vasilenko. Analisis of the thickness growth of Korean Cedar and Mongolian Oak in cedar – deciduous forest, Sikhote-Alin biosphere reserve // Journal of International Scientific Publications: Ecology & Safety, Volume 3, Part 2. 2009. P. 355-362.
• Galanin A.V., Galanina I.A. Peculiarities of flora, vegetation and lichen biota of the Changbaishan Volcano (China, East Asia) // Journal of International Scientific Publications: Ecology & Safety, Volume 4, Part 3. 2010. С. 457-470.
• May R.M. Patterns in multispecies communities // Theoretical ecology: principls and applitations. Blackwell ; Oxford, 1976. P. 147-162.

