 |
УДК 582.28 (581.9) (571.66)
Галанина И.А. Синузии эпифитных лишайников в дубовых лесах юга Приморского края. – Владивосток: Дальнаука, 2008. – 135 с.
Монография посвящена биологическому разнообразию синузий лишайников-эпифитов в дубовых лесах на юге Приморского края. Проведено сравнение биоты лишайников на дубе монгольском и дубе зубчатом в разных ландшафтах региона. В аннотированном списке лишайников эпифитов приводится 201 вид из 26 семейств и 61 рода. Сравнение показало, что видовой состав синузий лишайников-эпифитов на дубе монгольском и дубе зубчатом существенно различается. Установлен ботанико-географический градиент при переходе от флоры южного Сихотэ-Алиня к флоре Черных гор в Хасанском районе.
Ответственный редактор д.б.н. Л.Н. Васильева.
Рецензенты: д.б.н. З.М. Азбукина, д.б.н. А.В. Беликович, д.б.н. Осипов С.В.
Печатается по решению Ученого совета Ботанического сада-института ДВО РАН
ISBN 978-5-8044-0938-9
|
Глава 6. Анализ лихенобиоты дубняков юга Приморского края
6.1. Систематический анализ
В лихенобиоте дубняков юга Приморского края выявлен 201 вид, относящиеся к 61 роду, 26 семействам, 8 порядкам: Arthoniales, Gyalectales, Dothideales, Lecanorales, Ostropales, Pertusariales, Pyrenulales, Teloschistales. Наиболее значительно представлен порядок Lecanorales (17 семейств, 46 родов, 158 видов, или 79% от общего числа видов). Преобладание видов в лихенобиоте из этого порядка является характерной чертой голарктических флор (Харпухаева, 2004). В своих работах многие авторы использовали понятие лихенофлоры, но так как мы принимаем в данной работе понятие лихенобиоты, то здесь и далее при ссылках на работы авторов, использующих понятие флоры лишайников, считаем их эквивалентами. На остальные семь порядков приходится 42 вида из 15 родов и 9 семейств. Среди них наиболее богаты по числу видов порядки Pertusariales (1 семейство, 2 рода, 18 видов), Teloschistales (1 семейство, 2 рода, 9 видов).
К девяти наиболее крупным семействам относятся Parmeliaceae (54 вида), Physciaceae (43), Pertusariaceae (18), Lecanoraceae (17), Collemataceae (12), Teloschistaceae (9), Bacidiaceae (8), Lobariaceae (6), Ramalinaceae (6) (табл. 5). Семейства лишайников, отмеченных на дубе монгольском, располагаются следующим образом в порядке убывания видов: Parmeliaceae (54 вида), Physciaceae (43), Pertusariaceae (18), Lecanoraceae (17), Collemataceae (12), Teloschistaceae (9), Bacidiaceae (6), Lobariaceae (6), Ramalinaceae (6); на дубе зубчатом: Physciaceae (30 видов), Parmeliaceae (13), Lecanoraceae (8), Collemataceae (7), Pertusariaceae (5), Teloschistaceae (5), Ramalinaceae (5), Bacidiaceae (2).
На девять ведущих семейств приходится 173 вида, или 85.9 % от всей лихенобиоты, и 39 родов или 65% от общего числа родов лихенофлоры. На остальные 17 семейств приходится 26 видов, или 12.9% видового состава. Одно из этих семейств ( Roccelaceae ) содержит четыре вида. Шесть семейств (Brigantiaeaceae, Chrysothricaceae, Coccocarpiaceae, Coniocybaceae, Graphidaceae, Pannariaceae) представлены двумя видами. Одиннадцать семейств (Agyriaceae, Arthopyreniaceae, Candelariaceae, Catillariaceae, Gyalectaceae, Lecideaceae, Loxosporaceae, Megalosporaceae, Pyrenulaceae, Rocellaceae, Shinctrinaceae) включают по одному виду. Два вида имеют неясное положение в системе.
Таблица 5
Таксономический состав многовидовых семейств лишайников
Место во флоре |
Семейство |
Число родов |
Число видов |
Процент видов |
| |
|
|
1 |
Parmeliaceae |
18 |
54 |
26.9 |
2 |
Physciaceae |
8 |
43 |
21.4 |
3 |
Pertusariaceae |
2 |
18 |
8 .9 |
4 |
Lecanoraceae |
2 |
17 |
8.5 |
5 |
Collemataceae |
2 |
1 2 |
6.5 |
6 |
Teloschistaceae |
2 |
9 |
4 |
7 |
Bacidiaceae |
3 |
8 |
3.9 |
8 |
Lobariaceae |
1 |
6 |
2.9 |
9 |
Ramalinaceae |
1 |
6 |
2.9 |
|
Итого |
39 |
173 |
8 5.9 |
Лидирующим в лихенобиоте является семейство Parmeliaceae , насчитывающее 18 родов, 54 вида (26.9 % от общего числа видов). Виды этого семейства играют значительную роль в формировании лишайниковых группировок на дубах в южном Приморье. 43 вида из этого семейства более характерны для хвойношироколиственных лесов, в частности, долинных кедрово-широколиственных и кедрово-широколиственных с липой, в составе которых встречается дуб, а также высокогорных, сравнительно влажных, дубняков на границе с поясом темно-хвойных лесов. Большинство видов данного семейства встречается редко в Хасанском районе или их совсем нет, только с приближением к южным отрогам Сихотэ-Алиня они обнаруживаются чаще. Несмотря на это, большая часть видов семейства Parmeliaceae относится к неморальному элементу - 32, а к бореальному - только 15 видов. Остальные виды распределяются следующим образом: бореально-неморальные - 3, неморально-бореальные - 2.
Вторым семейством в лихенобиоте по числу видов является Physciaceae , оно включает 8 родов, 43 вида (21.4 %). Большинство видов относится к неморальному элементу - 35, к бореальному -5, к неморально-бореальному - 2, к мультизональному - 1. Виды этого семейства активно участвуют в формировании лишайникового покрова на дубах на юге Приморья. Здесь в южных дубняках они конкурируют за первое место с видами из семейства Parmeliaceae , а на сухих южных и юго-восточных склонах, особенно на дубе зубчатом на юге Хасанского района и в Лазовском заповеднике доминируют с большим проективным покрытием.
Третье семейство Pertusariaceae включает 2 рода, 18 видов (8.9 %). Виды данного семейства часто встречаются, с небольшим проективным покрытием, во всех изученных местообитаниях, но в основном на дубе монгольском. Большая часть видов этого семейства относится к неморальному элементу - 15, из них к неморальному горному элементу - 2 вида, к мультизональному - 1.
Четвертое семейство Lecanoraceae включает 2 рода, 17 видов (8.5 %). Как и в предыдущих семействах, ведущее место занимают виды неморального элемента - 12, к бореальному элементу относятся 3 вида, к мультизональному - 2.
Следующее семейство Collemataceae включает 2 рода, 12 видов (6.5 %). Большинство видов относится к неморальному элементу - 11 (из них один вид неморальный горный, один - неморальный океанический), к бореальному горному элементу относится 1 вид.
Ведущее положение семейств Parmeliaceae , Physciaceae , Pertusariaceae , Lecanoraceae (вместе составляют 132 вида и 65.7 % от всей лихенобиоты) типично для лихенобиоты умеренных широт Голарктики (Голубкова 1983; Урбанавичене, 1997; Катаева, 2002). По составу ведущих семейств и их количественным показателям видно, что лихенофлора дубняков юга Приморского края насыщена неморальными видами, характерными для пояса широколиственных лесов. При приближении к отрогам Сихотэ-Алиньского хребта лихенофлора дубняков Приморского края насыщается бореальными видами и видами, тяготеющими к хвойным породам деревьев.
К числу ведущих родов (табл. 6) относятся Lecanora - 15 видов (7.5 %), Pertusaria - 15 видов (7.5 %), Heterodermia - 10 (5 %), Phaeophyscia - 9 (4.5 %), Usnea - 9 (4.5 %), Physconia - 8 (4 %). Из них на дубе монгольском преобладают Lecanora - 15 видов, Pertusaria - 15, Heterodermia - 10, Phaeophyscia - 9, Usnea - 9, Physconia - 8 ; на дубе зубчатом: Lecanora - 7 видов, Collema - 7, Phaeophyscia - 7, Physconia - 6, Rinodina - 6, Heterodermia - 5 .
Первые 12 крупных родов содержат 105 видов, или 52.5 % от всей лихенофлоры. 17 родов (27.9 %) содержат по два вида, 23 рода (37.7 %) - по одному виду, оставшиеся 9 родов (14.8 %) - от 3-х до 5-ти видов.
Доминирующие виды из рода Heterodermia (H. hypoleuca, H. speciosa, H. diademata, H. japonica, H. micrphylla) тяготеют к более влажным, но открытым местообитаниям, это тропические и субтропические виды, заходящие в умеренный пояс ( Moberg , Nash , 1999; Trass , 2000).
Род Phaeophyscia включает следующие доминирующие виды: Ph. hispidula - обычный вид в горных районах в тропических и субтропических областях на всех континентах, реже встречается в умеренных областях. Виды Phaeophyscia hirtuosa и Ph . rubropulchra быстро возобновляются после пожаров, а в регулярно горящих лесах, особенно на дубе зубчатом, первый из них имеет большое проективное покрытие. Ph . sciastra - один из наиболее широко распространенных видов данного рода, но мы сомневаемся в достоверности его нахождения в районе исследования, так как он встречается в тропических и субтропических областях, где является высокогорным видом ( Moberg , 1993).
Доминирующие виды из рода Physconia ( Ph . detersa , Ph . subpulverulenta ) предпочитают открытые, хорошо освещенные участки ( Moberg , 1987). В исследованных нами местообитаниях это прежде всего Physconia detersa и Ph . subpulverulenta, немного реже встречается Ph . kurokawae .
Таблица 6.
Таксономический состав многовидовых родов лишайников
Род |
Число видов Процент видов |
| |
|
Lecanora |
15 |
7.5 |
Pertusaria |
13 |
6.5 |
Heterodermia |
10 |
5 |
Usnea |
10 |
5 |
Phaeophyscia |
9 |
4.5 |
Physconia |
8 |
4 |
Collema |
7 |
3.5 |
Parmelia |
7 |
3.5 |
Rinodina |
7 |
3.5 |
Caloplaca |
7 |
3.5 |
Lobaria |
6 |
3 |
Ramalina |
6 |
3 |
Итого |
105 |
52.5 |
6.2. Биоморфологический анализ
При биоморфологическом анализе за основу были взяты жизненные формы, разработанные Н.С. Голубковой (1983). Анализ жизненных форм показал, что все виды относятся к отделу эпигейных лишайников, к двум типам плагиотропных и ортотропных лишайников, это соответственно 193 и 8 видов.
Плагиотропные лишайники относятся к двум классам - накипных и листоватых лишайников, это соответственно 92 и 101 вид. Ортотропные лишайники представлены классом кустистых (8 видов). Наибольшее количество видов лишайников принадлежит к группе с листоватой рассечено-лопастной ризоидальной формой слоевища (89 видов, 44.3%) (табл. 7). На втором месте находится накипная зернисто-бородавчатая форма (38 видов, 18.9%) и на третьем месте - лишайники с накипной плотно-коркововой формой слоевища (33 вида, 16.4%).
6.3. Географический анализ
В основу выделения географических элементов в лихенобиоте юга Приморского края положен принцип поясности-зональности, принятый А.Н. Окснером (1944, 1946, 1948). Кроме того, использовались системы элементов Х.Х. Трасса (1970а) для лихенобиоты Эстонии, Н. В. Седельниковой (1985) для лихенобиоты нагорья Сангилен, с учетом современных ареалогических данных.
В лихенобиоте дубняков юга Приморского края выделено 9 географических элементов: неморальный, бореальный, мультизональный, арктобореальный, неморально-бореальный, бореально-неморальный, бореальный горный, неморальный горный и неморальный океанический (табл. 8).
Таблица 7.
Число видов различных жизненных форм лишайников
Жизненная форма |
Число видов Процент видов |
|
|
листоватая рассеченно-лопастная ризоидальная |
89 |
44.3 |
накипная зернисто-бородавчатая |
38 |
18.9 |
накипная плотно-корковая |
33 |
16.4 |
кустистая повисающая радиально-лопастная |
10 |
4.9 |
листоватая широко-лопастная |
8 |
3.9 |
кустистая повисающая плоско-лопастная |
8 |
3.9 |
накипная лепрозная |
7 |
3.5 |
листоватая вздуто-лопастная неризоидальная |
6 |
2.9 |
накипная однообразно-чешуйчатая |
2 |
0.9 |
Ведущее положение в географическом спектре принадлежит неморальному элементу, включающему 140 видов (69.6%). Преобладание неморальных лишайников закономерно, так как изучаемая территория находится в зоне хвойно-широколиственных лесов, южной подзоне лиановых хвойно-широколиственных лесов и подзоне неморально-хвойных лесов той же зоны (Ливеровский, Колесников, 1949; Колесников, 1955, 1961; Добрынин, 2000). Неморальные лишайники определяют облик лихенофлоры в приморских широколиственных лесах и относятся к 41 роду.
Таблица 8.
Распределение числа видов лишайников по географическим элементам
Географический элемент |
Число видов |
Процент видов |
неморальный |
140 |
69.6 |
бореальный |
37 |
18.5 |
мультизональный |
14 |
7 |
неморально-бореальный |
4 |
2 |
бореально-неморальный |
5 |
2.4 |
бореальный горный |
2 |
0.9 |
неморальный горный |
3 |
1.5 |
неморальный океанический |
6 |
3 |
арктобореальный |
1 |
0.5 |
Наибольшее число видов отмечено в родах Pertusaria (13 видов), Lecanora (12), Phaeophyscia (9), Physconia (8), Heterodermia (8), Rinodina (7), Collema (7), Lobaria (6), Ramalina (6), остальные роды содержат от 1 до 5 видов. Преобладают виды с мультирегиональным (52 вида, 26%) и восточноазиатским (32 вида, 16%) типами ареалов. К евразиатско-американскому относится 26 неморальных видов (11.9% от всех видов), к голарктическому типу ареала - 10 неморальных видов (5%). Остальные типы ареалов распределились таким образом: азиатско-американский - 5 видов (2.5%), евразиатский - 4 (2%), азиатский - 4 (2%), и 6 видов с неопределенным типом ареала (3%).
Как показывает ареалогический анализ, 26% неморальных лишайников относятся к видам с широким ареалом. По А. Н. Окснеру (1946), в составе неморальной лихенобиоты определенное место занимают виды автохтонных тургайских родов, но ведущая роль принадлежит поглощенным тургайской флорой видам тропогенного происхождения, основной ареал которых связан с тропическими и субтропическими районами земного шара.
Бореальный элемент стоит на втором месте в спектре географических элементов и включает 37 видов (18.5 %). Среди видов бореального элемента преобладают лишайники с евразиатско-американским ареалом - 14 видов (7%) и мультирегиональным ареалом - 15 (7.5%). Остальные виды бореального элемента распределились таким образом: голарктический ареал - 3 вида (1.5%), азиатско-американский - 2 (1), восточноазиатский - 1 (0.5) и 2 вида с неопределенным типом ареала. Это говорит о том, что формирование бореального элемента на юге Приморья происходило в составе бореальной лихенобиоты Голарктики (Чабаненко, 1990). Малое количество видов восточноазиатского ареала (1 вид, 0.5 %) указывает на слабую специфичность бореального элемента.
Мультизональный элемент находится на третьем месте географического спектра и включает 14 видов (7%), которые широко распространенны в Голарктике, а также в других флористических царствах. 10 видов (4.9%) этого элемента имеют мультирегиональный ареал распространения, 3 вида (1.5%), - голарктический ареал, 1 вид - евразиатско-американский. Большинство лишайников мультизонального элемента имеют широкую экологическую амплитуду, что позволяет им расселяться от арктических районов до областей теплоумеренной зоны, в аридных и даже в тропических районах земного шара. Среди них немало эвритопных видов, способных поселяться на самых различных субстратах и в самых разных местообитаниях (Голубкова, 1983).
Бореально-неморальных видов - 4, из них 2 вида имеют восточноазиатский ареал, 1 - мультирегиональный ареал и 1 вид с неясным типом ареала. Неморально-бореальных видов - 4, из них 2 вида имеют евразиатско-американский ареал, 1 - голарктический ареал и 1 вид с азиатским ареалом.
Горные виды, составляющие 2.5%, распределены между бореальным (2 вида, 1 %) и неморальным (3 вида, 1.5%) географическими элементами. Такой низкий процент горных видов подчеркивает разницу в лихенобиоте дубняков юга Приморского края и средней и северной его частей, которую в большей степени формирует Сихотэ-Алинское нагорье.
Неморальный океанический элемент составляет небольшой процент видов - 6 видов (3 %). Распространение этих видов связано с океаническими и субокеаническими регионами, и характерно для лихенобиоты наших приморских дубняков.
Анализ типов ареалов (табл. 9) показал преобладание лишайников с мультирегиональным ареалом - 81 вид (40% от всех видов). Евразиатско-американский тип ареала включает 43 вида (21.4%), восточноазиатский тип ареала - 32 вида (15.9%), голарктический тип ареала - 18 видов (8.9%), азиатско-американский - 8 видов (4%), евразиатский - 6 видов (3%), азиатский - 4 вида (2%), восточноазиатско-американский - 1 вид (0.5%), видов с неясным типом ареала - 8 (4%).
Анализ географических элементов показывает, что преобладают виды с неморальным распространением - 140 вида (69.6%), бореальным - 37 (18.5%), и мультизональным - 14 (7%), в том числе на дубе монгольском: неморальных - 131 вид (65.3%), бореальных - 32 (16.4%), мультизональных - 13 (6.5%); на дубе зубчатом: неморальных - 62 вида (30.8%), бореальных - 11 (5.5%), мультизональных - 7 (3.5%).
Анализ типов ареала показывает, что преобладают виды с мультирегиональным распространением - 81 вид (40%), евразиатско-американским - 43 (21.4%), восточноазиатским - 32 (15.9%), в том числе на дубе монгольском: мультирегиональных - 81 вид (40%), евразиатско-американских - 42 (20.9%), восточноазиатских - 32 (15.9%); на дубе зубчатом: мультирегиональных - 37 видов (18.4%), евразиатско-американских - 16 (7.9%), восточноазиатских - 15 (7.5%).
Таблица 9.
Соотношение видов по типам ареалов
Тип ареала |
Число видов Процент видов |
| |
|
Мультирегиональный |
81 |
40 |
Евроазиатско-американский |
43 |
21.4 |
Восточноазиатский |
32 |
15.9 |
Голарктический |
18 |
8.9 |
Азиатско-американский |
8 |
4 |
Евроазиатский |
6 |
3 |
Азиатский |
4 |
2 |
Восточноазиатско-американский |
1 |
0.5 |
Неясный тип ареала |
8 |
4 |
Таким образом, лихенобиота дубовых лесов юга Приморского края является неморальной. Преобладание мультирегиональных и евразиатско-американских видов говорит о древности ее формирования. Кроме того, наличие довольно большого количества видов с евразиатско-американским типом ареала в лихенобиоте дубовых лесов юга Приморья подтверждает наличие когда-то в далеком прошлом связи с Северной Америкой через берингийский мост суши (Галанин, Беликович, Галанина, 1998).
Количество восточноазиатских видов эпифитных лишайников в сравнении с сосудистыми растениями (Галанин и др., 2004), говорит о том, что процессы видообразования у лишайников протекают медленнее раза в три-четыре - возможно это связано с их медленным ростом.
Следует отметить важную особенность таксономического состава сообществ эпифитных лишайников на дубах на самом юге Хасанского района, где в массе еще встречается дуб зубчатый. Там не было найдено ни одного экземпляра из родов Usnea и Evernia , хотя севернее, для заповедника «Кедровая падь», который также находится в Хасанском районе, в литературе приводятся виды из родов Usnea и Evernia (Гурулева, Княжева, 1972, Княжева, 1973). Однако и в этих работах виды из этих родов приводятся в большей степени для хвойных пород деревьев. Встречаемость видов из родов Usnea и Evernia повышается при движении на север и восток к Лазовскому району: в окрестностях пос. Киевка в Лазовском районе вид Evernia mesomorpha был найден четыре раза, Usnea rubicunda - один раз, U . diplotypus - три раза. Все находки этих видов были на дубе монгольском. В Сихотэ-Алинском заповеднике на дубе монгольском эти роды представлены обильно и являются там обычными (Скирина, 1995).
Довольно распространенные и обильные в Сихотэ-Алинском заповеднике виды Parmelia saxatilis и P. fertilis на юге Хасанского района встречаются редко и необильно. Наиболее часто эти виды начинают встречаться по мере продвижения на северо-восток, их обилие при этом также увеличиваются . Уже на севере Хасанского района в окрестностях п. Кравцовка эти изменения становятся заметны. В целом, отмечено малое количество видов пармелиоидных лишайников в Хасанском районе и постепенное их увеличение на севере района в окрестностях п. Кравцовка и далее на восток к Лазовскому заповеднику .
В автореферате своей докторской диссертации «Распространение лишайников-эпифитов на территории России в естественных условиях и при антропогенном воздействии» А. В. Пчелкин (2002) применил к эпифитным лишайникам систему флористического районирования Земли, разработанную для сосудистых растений. В соответствии с ней, все дубняки на юге Приморья относятся к Восточноазиатской области Маньчжурской провинции. Материалы исследования этого автора были представлены данными из Сихотэ-Алинского заповедника, в котором для дуба монгольского приведены доминирующие виды со встречаемостью, выраженной в процентах: Parmelia aurulenta ( B = 57.7), P . saxatilis ( B = 32.0), Caloplaca gordejevi ( B = 42.3), Buellia disciformis ( B = 37.1), Parmelia fertilis ( B = 14.4).
На самом юге Хасанского района, где в массе встречается дуб зубчатый, картина с доминирующими видами в дубняках складывается совершенно иная. Здесь гораздо реже встречаются пармелиевые лишайники вообще, зато среди доминирующих лишайников выступают виды из семейства Physciaceae . Среди доминантов выделенных А. В. Пчелкиным (2002), сохраняют свое положение только Myelochroa aurulenta ( Parmelia aurulenta ) - 52% встречаемости и Caloplaca gordejevi – 62%. На юге Хасанского района встречаемость видов Parmelia saxatilis - 1% и P . fertilis - 11%. Вид Buellia disciformis здесь вообще встречается крайне редко (2%) на дубе монгольском и не встречен совсем на дубе зубчатом, но становится часто встречающимся видом в дубовых лесах южного Сихотэ-Алиня (Скирина, 1995).
Встречаемость видов-доминантов, указанных А. В. Пчелкиным (2002) для Сихотэ-Алинского заповедника, увеличивается на севере Хасанского района в окрестностях п. Кравцовка, где с дубом произрастает пихта черная, и при движении на восток Приморского края, где влияние Сихотэ-Алинской флоры становится более ощутимым, эти виды чаще встречаются, появляясь и на дубе зубчатом, в Лазовском районе.
Подобная картина распределения и изменения встречаемости видов эпифитных лишайников на дубах на юге Приморского края повторяется с другими видами. Например, вид Flavoparmelia caperata на юге Хасанского района не является доминантом и встречается изредка (6 %), причем в месте произрастания бывает обильным, а далее на север и восток в южной части Приморского края становится доминирующим (до 86 %) в дубняках из дуба монгольского. Так же ведут себя виды Collema furfuraceum (от 2% до 31%), Lecanora argentata (от 1% до 9%), L . с hlarotera (от 17% до 28%), Caloplaca flavorubescens (от 1% до 29%), Ochrolechia parella (от 12% до 57%), Parmotrema chinense (от 21% до 49%), Parmelia saxatilis (от 1% до 21%).
Таким образом, в результате вышеописанного распределения эпифитных лишайников на дубах на юге Приморского края выявляется некий градиент при переходе от Сихотэ-Алиня к Черным горам, протянувшимся вдоль побережья моря в Хасанском районе. Возможно, это граница между двумя лихенобиотами: «корейской» и «сихотэ-алинской». Все вышесказанное позволяет предположить, что самый юг Хасанского района в лихенологическом районировании может быть отнесен к иной провинции, нежели Сихотэ-Алинь с его южными отрогами.
Заключение
На юге Приморского края флора лишайников-эпифитов, встречающихся на дубе монгольском и дубе зубчатом, насчитывает 201 вид, относящихся к 61 роду и 26 семействам. Из них 197 видов (60 родов и 25 семейств) встречается на дубе монгольском и 82 вида (33 рода и 13 семейств) - на дубе зубчатом, соответственно. Общими для дуба монгольского и дуба зубчатого являются 78 видов.
Впервые изучены эпифитные лишайники на дубе зубчатом, для которых характерны следующие особенности по сравнению с лишайниками на дубе монгольском: меньшее таксономическое разнообразие, меньшее разнообразие лихеносинузий, меньшая встречаемость накипных и кустистых жизненных форм, смена лидирующих таксонов, преобладание неморальных и восточноазиатских элементов.
Построено пять графов, отражающих распределение видов на дубе монгольском и дубе зубчатом в нескольких местообитаниях Хасанского и Лазовского районов. В каждом из них выделены группы площадок на основании характерных сочетаний видов. Сравнительный анализ этих комбинаций в отношении доминирующих и редких видов позволил выявить существенные различия, обусловленные характером местообитаний и географическим положением.
По составу доминирующих лишайников, образующих “ядра” характерных видов, на обоих видах дуба - на юге Хасанского района и в южных отрогах Сихотэ-Алинских гор Лазовского района - выявлены две обособленные лихенобиоты, условно названные “корейская” и “сихотэ-алинская”.
На юге Хасанского района “ядра” характерных видов на дубе зубчатом и дубе монгольском имеют много общего (виды, их встречаемость, проективное покрытие), что говорит о большей давности связей этих двух пород в исследованных местообитаниях. Лихенобиота на дубе зубчатом здесь более видоспецифична по сравнению с таковой в Лазовском районе.
В Лазовском районе “ядра“ характерных видов на дубе зубчатом и дубе монгольском существенно отличаются. Видовой состав лишайников на дубе зубчатом здесь обогащается за счет видов, более характерных для дуба монгольского.
Таксономический и географический анализ видового состава лишайников на дубе монгольском и дубе зубчатом подтвердил наличие границы между двумя лихенобиотами - "корейской" и "сихотэ-алинской". Данное исследование может быть использовано при уточнении лихенологического районирования юга Дальнего Востока.

Фото 3. Лес из дуба зубчатого, Лазовский р-н (фото Н.А. Василенко).
|

Фото 4. Лес из дуба монгольского, Лазовский р-он (фото И.А. Галаниной). |
Виды, составляющие ядро характерных видов дуба монгольского.

Фото 5. Lecanora paсhyheila (фото И.А. Галаниной).
|

Фото 6. Myelochroa aurulenta (фото Stephen Sharnoff, 2001).
|

Фото 7. Heterodermia hypoleuca (фото Stephen Sharnoff, 2001).
|
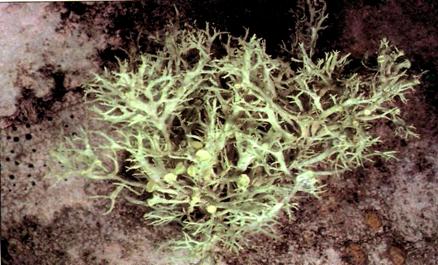
Фото 8. Ramalina dilaceratа(фото Stephen Sharnoff, 2001).
|

Фото 9. Tephromela atra (фото Stephen Sharnoff, 2001).
|

Фото 10. Lecidella elaeochroma (фото Stephen Sharnoff, 2001).
|
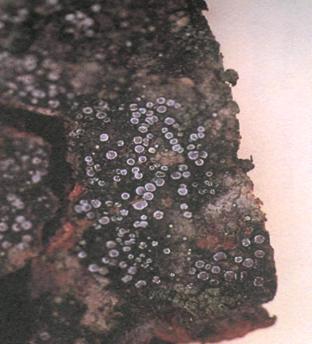
Фото 11. Schismatomma pericleum (фото Stephen Sharnoff, 2001).
|

Фото 12. Phaeophyscia hispidula (фото Stephen Sharnoff, 2001).
|
Виды, составляющие ядро характерных видов лишайников дуба зубчатого

Фото 13. Phaeophysca rubropulchra (фото Stephen Sharnoff, 2001).
|

Фото 14. Physconia detersa (фото Stephen Sharnoff, 2001).
|

Фото 15. Phaeophysca hirtuosa (фото И.А. Галаниной).
|

Фото 2. Лес из дуба зубчатого, Хасанский р-н (фото И.А. Галаниной).
|
Общие виды для обоих дубов, входящие в состав ядер характерных видов

Фото 16. Caloplaca gordejevi (фото И.А. Галаниной).
|

Фото 16. Caloplaca gordejevi (фото И.А. Галаниной).
|

Фото 17. Myelochroa entotheiochroa (фото И.А. Галаниной).
|

Фото 18. Lecanora allophana(фото Stephen Sharnoff, 2001).
|

Фото 1. Лес из дуба монгольского, Хасанский р-н (фото И.А. Галаниной).
|
|



