Лекция 1. Основные принципы организации биосферы и экосистем
А.В. Галанин © 2012
В лекции рассматриваются следующие вопросы:
1. Два состояния атомов химических элементов – связанное и свободное.
2. Подвижные атомы и возникновение жизни.
3. Большой геологический круговорот вещества.
4. Малый биологический круговорот вещества. Биосфера и экосистемы.
5. Параметры биологического круговорота.
6. Эволюция биологического круговорота на Земле и эволюция жизни.
Лекция была прочитана 29 марта 2012 г. на "Профессорских чтениях" в институте Технологии и бизнеса в г. Находка для преподавателей школ и вузов города и района. Судя по реакции слушавших, содержащаяся в ней информация не была для них бесполезна. Думаю, она не будет бесполезна преподавателям, учащимся и студентам других сел и городов России.
Введение
Экология по своей сути является частью биосферологии, а биосферология, в свою очередь, - частью планетологии, или землеведения. Поэтому начну эту лекцию издалека - из космоса, окинув взглядом нашу планету и соседние с ней планеты Солнечной системы. Для каждой планеты здесь взяты по два ракурса: один ракурс - вид с околосолнечной, а другой - с околопланетной орбит. Здесь приводятся снимки NASA, которому я выражаю признательность.

Вид Земли и Луны из космоса. Марс Глобал Сервейор сделал этот снимок с орбиты Марса. Фото с сайта: http://nenosfirs.ucoz.ru/load/kosmos/mars/vzgljad_v_nebo_marsa_s_foto/33-1-0-62
|

Так выглядит наша живая планета с азотно-кислородной атмосферой, океанами воды, с активной биосферой и развитой жизнью. Луна - мертвый спутник Земли, практически лишенный атмосферы, жидкой воды и жизни. Фото с сайта: http://www.ellf.ru/photos/26214-ochen-krasivye-foto-zemli-iz-kosmosa-7-foto.html |

Вид части США с околоземной орбиты. Фото с сайта: http://bipbap.ru/interesting/334-potryasayuschie-fotografii-s-orbity-zemli.html
|

Живая Земля и мертвая Луна.
|
А теперь обратим внимание на нашего соседа - Марс. Его орбита немного (по космическим масштабам конечно) дальше от Солнца, чем орбита Земли. Марс меньше Земли, но тем не менее его гравитация удерживает разреженную атмосферу, на 95% состоящую из углекислого газа. На Марсе холодно, ночью даже на экваторе температура падает до - 60°С, а днем на экваторе бывает довольно тепло - до +15°С. На полюсах Марса зимой мороз бывает такой, что углекислый газ атмосферы снежинками падает на его поверхность, атмосфера зимой на полюса Марса выпадает в виде углекислого снега. Весной и летом углекислый снег испаряется, отчего на полюсах растет атмосферное давление, срабатывает парниковый эффект и становится по-марсиански совсем тепло. Зимой ветер дует от экваторов в сторону полюсов, а летом наоборот. Есть на Марсе и вода, только она здесь преимущественно в замерзшем виде. При таянии водяного льда вода быстро испаряется, так как давление очень низкое и вода здесь закипает уже при +3°С. Марсианский лед находится в грунте (криолитозоне) и в ледниках, которые на полюсах достигают огромных размеров.

Вид Марса из космоса. Шрам на его поверхности - это глубокий (до 3-4 км) тектонический разлом, шириной до 300 км и длиной до 4 тыс. км. Несмотря на холод и разреженную атмосферу, на Марсе есть круговорот воды и углекислого газа, а в толще криолитозоны, вероятно, существуют резервуары с жидкой водой и жидкими и газообразными углеводами - нефтью и природным газом абиогенного происхождения. Глубоко в толще криолитозоны Марса водяной лед может растаивать под действием эндогенного тепла планеты.
|
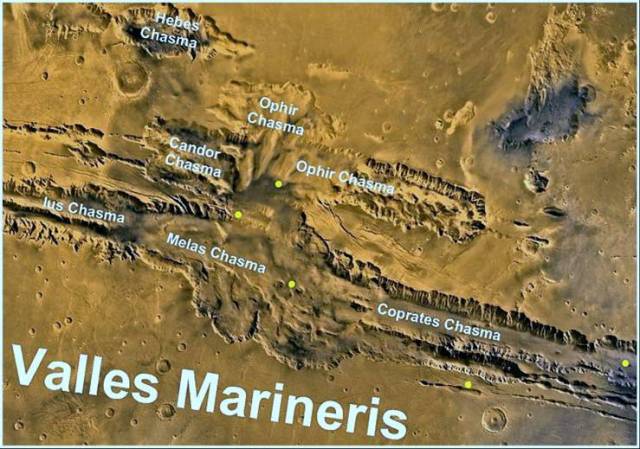
Так выглядит планета с разреженной углекислой атмосферой, с неразвитой жизнью и отсутствующей или очень малоактивной биосферой. Температура на поверхности Марса изменяется от + 15 до – 160°С.
|
Планета Венера находится к Солнцу ближе, чем Земля, по размерам она немного уступает Земле. Венера удерживает мощную атмосферу, однако состоит эта атмосфера преимущественно из углекислого газа, отчего парниковый эффект на Венере такой, что температура около поверхности достигает +400°С. Воды на Венере очень мало, кислорода тоже только следы. Зато есть озера жидкой серы. А в атмосфере плавают облака из капелек сернистой кислоты, и на поверхность выпадают кислотные дожди.

Вид Венеры из космоса. Для жизни эта планета непригодна.
|

Так выглядит мертвая планета Венера с очень мощной углекислой атмосферой, раскаленной поверхностью, непригодная для жизни. Температура на поверхности Венеры около +400 °С.
|
Ближе всех планет к Солнцу находится Меркурий. Размеры его с Луну. Атмосферы Меркурий не имеет, т.к. его гравитации не хватает для этого. Обращенная к Солнцу поверхность Меркурия раскаляется до +200°С и, возможно, даже больше. Но когда Солнце заходит, то поверхность быстро остывает до -200°С. Воды на поверхности Меркурия нет, разве что в глубине его бесчисленных кратеров есть ледники, засыпанные грунтом. Планета безжизненна.

Вид Меркурия из космоса. Так выглядит мертвая планета, с раскаленной поверхностью на стороне, обращенной к Солнцу, и очень холодной на стороне, не обращенной к Солнцу. Планета практически лишена атмосферы, здесь отсутствуют жизнь и биосфера.
|

Геологически Меркурий активен. На него не только падают метеориты, но на нем извергаются и вулканы.
|
Небольшой космический экскурс показал, что из четырех планет земной группы биосфера и активная жизнь есть только на нашей Земле. Все четыре (включая Луну) наших космических соседа мертвы. Исключением может быть только Марс, но если там и есть жизнь, то она очень примитивная в виде микроорганизмов, живущих анаэробно в толще криолитозоны, и биосфера Марса, если она есть, чрезвычайно малоактивная.
Геологический круговорот вещества на Земле
Вещество на Земле находилось в геологическом круговороте. Под действием эндогенных сил возникали и возникают горы и межгорные впадины. Горы разрушаются, вещество смывается в моря, где оседает на дно и постепенно заполняет эти впадины. Накопленные толщи осадков спресовывались под собственной тяжестью и частично оплавлялись под воздействием эндогенного тепла Земного шара. В результате на дне морей и океанов образовывались многокилометровые толщи морских осадков. При этом горы на суше под воздействием текущих вод и ветров разрушались, и на их месте образовывались равнины - пенеплен.
Проблемы геологического круговорота вещества на Земле рассматривались многими учеными (Григорьев, 1948; Виноградов, 1967; Бондарев, 1974; Вернадский, 1978; Марков и др., 1978; Галанин, 1990 и др.). Эти проблемы входят в компетенцию науки геологии.
Новые горообразовательные процессы, происходившие при очередном сжатии Земного шара и уменьшении его объема, сминали земную кору на дне океанов в складки, так возникали новые горные цепи. Впадины морей и океанов при последующих растяжениях земной коры при расширении Земного шара образовывались уже в других местах. Земная кора разламывалась на отдельные блоки, в разломы поступало расплавленное вещество из мантии Земли и раздвигало блоки земной коры. В таких местах (зоны спрединга) образовывались новые моря и океаны. И все это в геологической истории нашей планеты повторялось многократно. Так на Земле осуществлялся и осуществляется большой геологический круговорот вещества.
При этом атомы мигрировали из кристаллических решеток минералов в воду (растворялись в воде) и выносилось текучими водами в моря, размельченное вещество (песок и пыль) во взвешенном состоянии также переносились текучими водами и ветрами.
Биологический (малый) круговорот вещества возник в результате эволюции большого геологического круговорота. Он возник в соответствии с принципом Ле-Шателье как препятствие на пути рассеяния вещества в большом геологическом круговороте.
Как известно, вещество состоит из молекул, а молекулы из атомов. В простых веществах молекулы состоят из атомов одного химического элемента, в сложных – разных. Атомы и молекулы могут образовывать кристаллические решетки.
Но есть состояние атомов, когда они не связаны ни в молекулы, ни в кристаллические решетки. Это так называемые свободные, или подвижные, атомы. В подвижном несвязанном (рассеянном) состоянии могут находиться атомы всех химических элементов, но одни химические элементы переходят в подвижное состояние легче (быстрее), другие - медленнее. Свободные атомы способны к миграции (перемещению), они перемещаются не только в атмосфере и воде, но и в твердых субстанциях. Впервые на свободное (рассеянное) состояние атомов химических элементов обратил внимание В.И. Вернадский.
Благодаря способности переходить то в связное, то в рассеянное состояние, атомы в земной коре, атмосфере и гидросфере способны к геохимической миграции. Но атомы мигрируют не только в неживом веществе планеты, но и в живом (в живых организмах). Жизнь вообще - это высоко организованное движение атомов, молекул, радикалов, электронов. Следует сказать, что в основе жизни находятся подвижные атомы и ионы различных химических элементов.

Гималаи – это смятое в складки дно океана Тетис.
Тихий Оекан и цепь островов. Так выглядит наша планета с околоземной орбиты.
Эндогенные геологические процессы формируют на планете многочисленные термодинамические градиенты, связанные с неоднородностью в распределении атомов различных химических элементов. Эта неравномерность связана с неоднородностью рельефа, неоднородностью распределения горных пород, климатическими различиями. Термодинамические градиенты в литосфере, гидросфере и атмосфере Земли являются причиной геологического круговорота. Свободные подвижные атомы из мест с большей концентрацией движутся в сторону мест с меньшей концентрацией. Термодинамические градиенты на планете в результате геологического круговорота постепенно разрушаются, и структурная энтропия Земли возрастает. В геоморфологии эти процессы известны как процессы пенепленизации. Биосфера же работает против второго начала термодинамики. Живые организмы, напротив, улавливают подвижные атомы и ионы и концентрирует их в живом веществе.
Биологический круговорот вещества
Переход атомов из состояния рассеяния в упорядоченное (связанное) состояние совершается как абиогенно, так и посредством живого вещества (Вернадский, 1978). Биота (или живое вещество, по В. И. Вернадскому) тормозит возрастание в биосфере структурной энтропии и не только способствует сохранению термодинамических градиентов, но и усиливает их. Например, некоторые морские организмы способны концентрировать рассеянные в воде химические элементы в 600 000 раз (Марков и др., 1978), а растительный покров суши может почти полностью остановить твердый и эоловый сток вещества с материков в моря и океаны и резко снизить ионный сток вещества (Бондарев, 1974).
Под биосферой понимается глобальная экологическая система биогенной миграции химических элементов, способная к сохранению и поддержанию подвижного биогеохимического равновесия между литосферой, атмосферой, гидросферой и биотой. Биосфера включает в себя следующие блоки или компоненты: 1 - живые организмы (живое вещество), 2 - мертвое органическое вещество биогенного происхождения, 3 - неорганическое вещество атмосферы, 4 - неорганическое вещество гидросферы, 5 - неорганическое вещество литосферы, вовлеченное в биологический круговорот. В составе живого вещества присутствуют многие химические элементы, но преобладают углерод, водород, азот, фосфор, кальций, калий.
Биосфера возникла и функционирует как система химического и энергетического взаимодействия атмосферы, гидросферы и литосферы в постоянно циклически и ациклически изменяющихся условиях. Циклы изменения физических и химических параметров на Земле бывают суточными, годичными, одиннадцатилетними, вековыми, тысечелетними, миллионолетними вплоть до галактического года длительностью примерно в 200 миллионов лет.
Центральное место в цепи перехода химических элементов из одного состояния в другое принадлежит живому веществу - биоте. Обеспечивая биологический или малый круговорот химических элементов, биота получает из внешней среды не только вещество и энергию, но и негэнтропию, и формирует биохимическое разнообразие в широком смысле.
Сущность процесса биологической эволюции – это увеличение организованности биосферы, возрастание ее негэнтропии. В процессе эволюции происходит превращение энтропии внешней среды (энтропии большого геологического круговорота) в негэнтропию экосистем - т.е. в негэнтропию малого биологического круговорота, в конечном счете в негэнтропию популяций видов и генетических структур.
Главная причина постоянно идущих химических процессов на Земле – это возникновение неоднородности или создание градиентов концентрации веществ - неоднородности в распределении атомов разных химических элементов в разных средах и в разных местах одной среды. Диффузия атомов и молекул из мест их большей концентрации в места их меньшей концентрации, а также неоднородности полей температуры и давления, неравномерный приток солнечной энергии приводят к перемещению воздушных и водных масс – к циркуляции атмосферы и гидросферы.
За многие миллионы и миллиарды лет (в соответствии со вторым началом термодинамики) все неоднородности планеты должны были бы исчезнуть - таков закон возрастания энтропии. Однако они не исчезают, т.к. в природе происходит постоянное воспроизводство этих неоднородностей в результате землетрясений, извержения вулканов, движений блоков земной коры.
Атмосфера Земли первоначально возникла в результате дегазации недр – т.е. в результате вулканической деятельности - и состояла из аммиака, метана, сернистого газа, углекислого газа и паров воды. Переход к кислородной атмосфере произошел не сразу. Вначале свободный кислород образовывался в результате разложения молекул паров воды под действием ультрафиолета в верхних слоях атмосферы. Однако при этом кислорода образовалось очень мало. К тому же он расходовался на окисление минералов на поверхности планеты.
Главным процессом, в результате которого атмосфера Земли обогащалась кислородом, был фотосинтез сначала синезеленых водорослей или цианобактерий, а затем зеленых водорослей и наземных растений. Это коренным образом изменило геохимическую обстановку на планете – в ее биосфере, изменив геологические, физико-географические, биогеохимические и эволюционно-биологические процессы.
Первичная атмосфера Земли имела восстановительный характер. Но по мере накопления в ней кислорода она все более становилась окислительной атмосферой. А.И. Опарин и Дж. Холдейн считали, что только в бескислородной атмосфере могла зародиться жизнь, т.к. в кислородной атмосфере органические и неорганические молекулы быстро окисляются - в кислородной среде они не стойки. В докембрийской атмосфере Земли содержание кислорода было ничтожным (Виноградов, 1947).
А. А. Григорьев (1948) связал вопрос об эволюции биосферы с изменением концентраций кислорода и углекислого газа в атмосфере. Объем кислорода, выделяемого при фотосинтезе, вначале не превышал его расход на дыхание организмов и на окисление химических соединений. Но постепенно кислорода растениями стало выделяться больше, чем его расходовалось в биосфере. В итоге количество кислорода в атмосфере стало постепенно возрастать.
С достижением 2 млрд. лет назад концентрации кислорода в атмосфере 0.001 от его современного уровня стало возможным развитие аэробных организмов, отчего накопление кислорода ускорилось и фотосинтезирующая деятельность растений превратилась в основной источник кислорода. К началу фанерозоя (600 млн. лет назад) концентрация кислорода в атмосфере Земли достигла 1% от его современного содержания.
В конце силура содержание кислорода составило уже 10% от современного, и это сделало возможным выход жизни на сушу,так как в верхних слоях атмосферы возник озоновый слой из трехатомных молекул кислорода (озона), поглощающих жесткое ультрафиолетовое излучение. При этом жесткое ультрафиолетовое излучение уже не могло разрушить молекулы ДНК и РНК - носители наследственной информации.
В карбоне (меловом периоде) содержание кислорода было в 2-3 раза выше современного. Это было связано с очень интенсивной растительной жизнью - чрезвычайно интенсивным фотосинтезом. При этом растения израсходовали почти весь углекислый газ и аккумулировали углерод в толщах каменного угля. Это затормозило фотосинтез на Земле, и углекислый газ вновь стал накапливаться в результате вулканической деятельности и в результате активизации животных, грибов, бактерий, выделявших углекислый газ при дыхании. В это время быстро эволюционировала нисходящая ветвь биологического круговорота. В мезозое концентрация кислорода стала близка к современной. Рост концентрации свободного кислорода привел к бурной адаптивной радиации животной и грибной жизни.
Наконец жизнь б.м. стабилизировала газовый состав атмосферы и стала регулятором климата. Когда увеличивалось содержание углекислоты в атмосфере и гидросфере, то усиливался парниковый эффект. Климат становился теплее, отчего активнее шел фотосинтез, т.к. растения при высокой концентрации во внешней среде углекислого газа и теплой окружающей среде фотосинтезируют значительно быстрее. В результате такой активизации растения снижали концентрацию улекислого газа, и атмосфера остывала, при этом снижалась интенсивность фотосинтеза, и в атмосфере накапливался углекислый газ.
Уменьшение содержания углекислого газа в атмосфере привело в конце палеозоя к падению биопродуктивности растительности и к существенному (примерно в 2 раза) снижению общей массы живого вещества биосферы.
Биота - это особая дискретная система, состоящая из особей живых организмов, это - живое вещество по В. И. Вернадскому. Но биота – не аморфная биомасса, а высоко организованная система, способная, в свою очередь, организовывать внешнюю среду. Именно биота превратила неживую внешнюю среду на Земле в биосферу, изменив ее при этом кардинальным образом.
Важной особенностью биоты является ее способность к воспроизводству в результате размножения особей, в результате смены поколений и передачи наследственной информации от предыдущего поколения к последующему. При этом носителями наследственной информации являются особые наследственные вещества – дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК).
Экологический кризис перепроизводства мертвой органики в конце каменноугольного периода, вероятно, привел к значительному снижению в атмосфере содержания углекислого газа, к образованию и накоплению в огромных количествах торфов и каменных углей. Это привело к резкому снижению парникового эффекта атмосферы, похолоданию и мощному оледенению.
|
Движение вещества через биоту не всегда было циклическим. Биологический круговорот, каким его мы видим сейчас, на Земле возник не сразу. Например, системы океана и древнейшие экосистемы суши работали на осаждение вещества и имели незамкнутый, нециклический тип миграции химических элементов. Свободные подвижные атомы связывались в органическом веществе в скелетах водорослей и морских животных, в фитомассе отмирающих растений и преставали быть свободными. В таком состоянии биосфера суши Земли, вероятно, находилась с девонского периода до середины каменноугольного периода, а экосистема океанов и морей в таком состоянии находится и в настоящее время.
Тогда разложение мертвого органического вещества на суше не успевало за его синтезом растениями. Происходила повсеместная олиготрофизация экосистем (накопление в них мертвой органики) и от этого резко снижалась их продуктивность, так как почти все подвижные химические элементы оказались осажденными в толщах мертвой органики, покрывавшей тогда не только заболоченные равнины, но и склоны холмов и гор. Позднее это вещество в толщах Земли превратилось в каменные и бурые угли.
|
В экосистеме органические вещества синтезируются автотрофами из неорганических веществ. Затем они потребляются гетеротрофами. В результате выделения в процессе жизнедеятельности или после гибели организмов органические вещества подвергаются минерализации, т.е. превращаются в неорганические вещества. Эти неорганические вещества могут быть вновь использованы для синтеза органических веществ автотрофами.
В структуре экосистем следует различать две части:
1 - резервный фонд – вещество, которое в данное время не связанное с живыми организмами, но может ими использоваться;
2 - обменный фонд – вещество, которое в данный момент входит в состав живых организмов и передается по цепям питания.
В зависимости от расположения резервного фонда биологические круговороты можно разделить на два типа:
1 - круговороты газового типа с резервным фондом веществ в атмосфере и гидросфере (углерод, кислород);
2 - круговороты осадочного типа с резервным фондом в земной коре (фосфор, кальций, железо и др.).
Круговороты газового типа обладают большим обменным фондом, а круговороты осадочного типа меньшим, так как основная масса вещества содержащегося в земной коре находится в связанном и потому «недоступном» живым организмам виде. Круговороты осадочного типа легко нарушаются от различного рода воздействий, и при этом часть вещества выходит из биологического круговорота в круговорот геологический. Возвратиться опять в биологический круговорот она может, но уже в результате геологических процессов - орогенезов, вулканической деятельности. Извлечь нужные живым организмам вещества из земной коры из связанного состояния живым организмам гораздо сложнее, чем из атмосферы. Хотя такие возможности у растений есть: это выделение корнями во внешнюю среду кислот, которые переводят атомы и ионы из кристаллических решеток в свободное состояние - в почвенные растворы. Минералы могут растворять лишайники эпилиты, поселяющиеся на камнях и скалах. Но растут эти лишайники чрезвычайно медленно.
Интенсивность биологического круговорота зависит от температуры окружающей среды и количества в ней воды, т.е. от степени подвижности свободных атомов и ионов, от их количества во внешней среде. Так, например, биологический круговорот интенсивнее протекает во влажных тропических лесах, а медленнее - в северных бореальных лесах и в тундре. Кроме того, в бореальных лесах и в тундре биологические процессы протекают только в теплое время года, а во влажных тропических лесах они идут круглый год.
Растительный покров - это блок биотического покрова Земли, накапливающий в биосфере световую энергию в форме энергии химических связей продуктов фитосинтеза углеводов; потребляющий из атмосферы и гидросферы углекислый газ; обогащающий атмосферу и гидросферу кислородом. Посредством биологического круговорота в биологический цикл превращений из внешней среды вовлекаются элементы минерального питания и вода. Для того чтобы получить из почвенного раствора необходимые элементы минерального питания, растения всасывают корнями большое количество воды, поднимают ее по сосудам к листьям и испаряют воду, забирая растворенные в ней вещества.
Фотосинтез - это процесс образования органического вещества из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилла у растений, и бактериохлорофилла и бактериородопсина у синезеленых водорослей или цианобактерий). Фотосинтез является совокупностью процессов поглощения, превращения и использования энергии квантов света в различных реакциях, в том числе превращения углекислого газа в органическое вещество.
В основе фотосинтеза лежат окислительно-восстановительные реакции, в которых электроны переносятся от донора (молекул воды H2O) к акцептору (CO2) с образованием восстановленных соединений (углеводов) и выделением кислорода. Интегрально процесс фотосинтеза описывается следующей химической реакцией:
СО2 + Н2О + свет > углевод + О2
Рассмотрим круговороты наиболее значимых для живых организмов веществ. К малому биогеохимическому круговороту биогенных элементов относятся круговороты углерода, азота, фосфора, серы и др. http://xreferat.ru/10/1977-1-rol-zhivyh-organizmov-v-biologicheskom-krugovorote.html
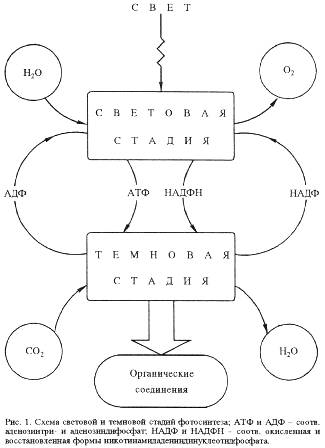
- В действительности фотосинтез - очень сложный и многоступенчатый процесс, состоящий из световой и темновой стадий. Световая стадия состоит из разложения воды на кислород и водород, при этом из молекулы воды квантами света выбивается и активизируется электрон, который приводит к переходу АДФ в АТФ (аденозиндифосфорной кислоты в аденозинтрифосфорную кислоту) и НАДФ в НАДФН. В темновой фазе за счет энергии молекул АТФ и НАДФН молекулы углекислого газа активизируются, и из них происходит синтез полимеров - органических соединений. При этом атомы водорода, захваченные в световой стадии из молекул воды, соединяются с атомами кислорода из углекислого газа и образуют воду. Схема с сайта:
- http://www.ximuk.ru/encyklopedia/2/4899.html
|
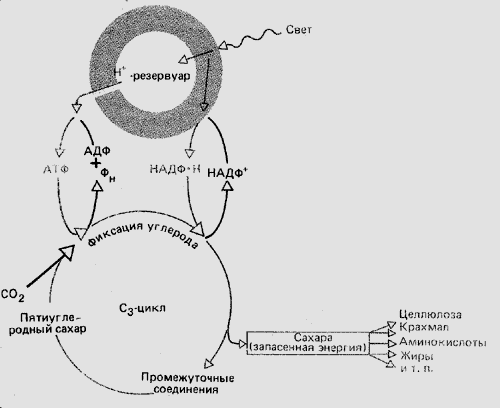
- На этой схеме также показаны две стадии фотосинтеза: световая, на которой из молекул воды выбиваются протоны и электроны водорода, и темновая, на которой происходит образование органического вещества из углекислого газа. Схема с сайта: http://bio.1september.ru/article.php?ID=200100302
|
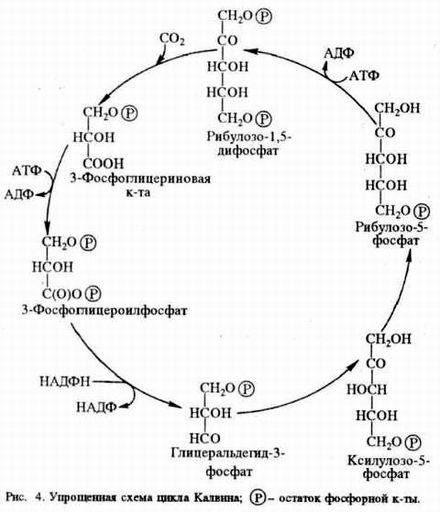
Схема темновой фазы фотосинтеза (цикл Кальвина). Схема из химической энциклопедии "Фотосинтез" по адресу: http://www.edudic.ru/hie/1638
|
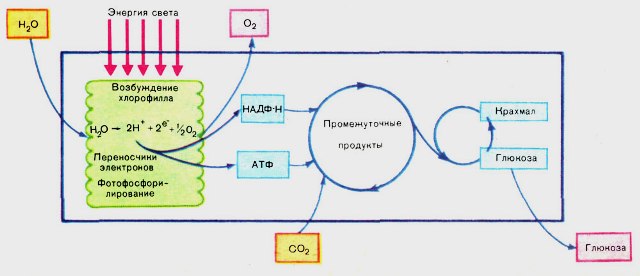
- Еще одна схема фотосинтеза, показывающая более детально темновую фазу фотосинтеза. Схема с сайта: http://www.ejonok.ru/nature/Биология/Фотосинтез.
Энергия света возбуждает молекулу хлорофилла, которая взаимодействует с молекулой воды и разрушает ее. При этом в системе появляются 2 свободных "энергичных" электрона, 2 протона (2 положительно заряженных ядра водорода) и один атом кислорода. Свободные атомы кислорода объединяются по два и образуют молекулы кислорода. А протоны переносят энергию в молекулы АТФ и НАДФН.
Следует сказать, что растения не только фотосинтезируют, но и дышат, поглощая при этом кислород и выделяя углекислый газ. При дыхании растения (как животные и грибы) извлекают запасенную ими на свету энергию квантов света и расходуют ее для построения своего тела. Дыхание растений происходит ночью. Поэтому не следует держать растения в спальнях. Однако фотосинтез у растений во много раз превосходит дыхание, поэтому в итоге растения больше выделяют кислорода при фотосинтезе, чем поглощают его при дыхании.
|
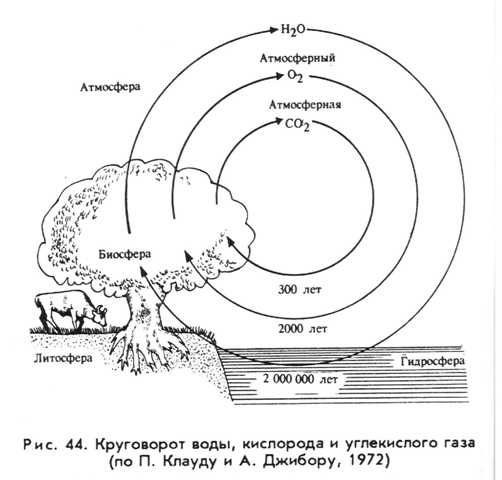
Схема взята с сайта по адресу: http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
|
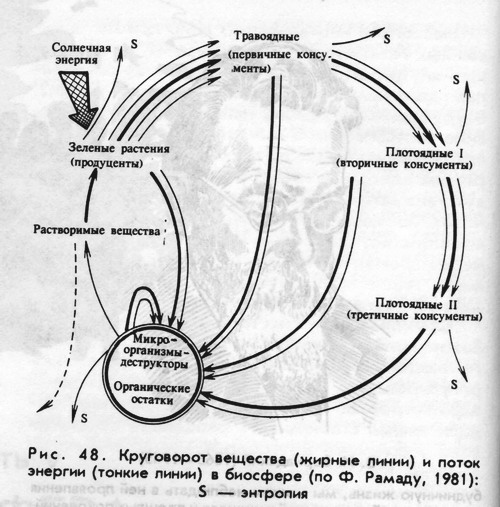
Схема взята с сайта по адресу: http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
|
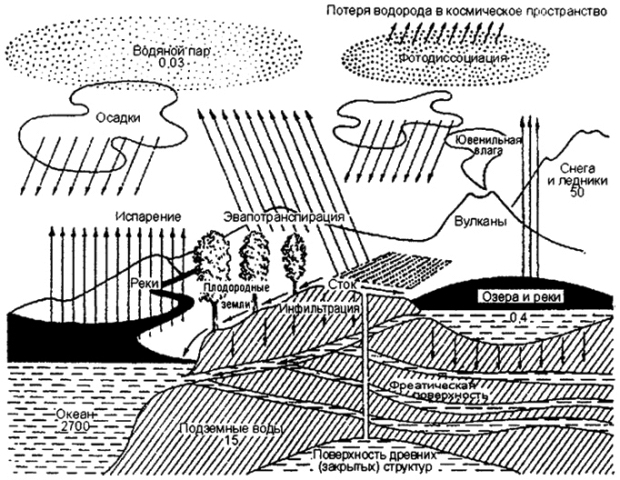
Структура круговорота воды на Земле. Схема взята с сайта по адресу: http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
|
Вода на Земле находится в трех агрегатных состояниях: жидком, газообразном, твердом и кристаллическом. Кроме того, вода входит в состав тел живых организмов, порой ее содержание там достигает 95% от общего веса тела и даже иногда болше. Состояние воды в живых организмах можно рассматривать как еще одно ее агрегатное состояние - живая вода. Испаряясь жидкая вода становится паром и поступает в атмосферу, где пар переносится на большие расстояния. Охлаждаясь, водяной пар становится туманом, и парообразная вода снова переходит в жидкое состояние, а если воздух очень холодный, то капельки воды становятся снежинками - переходят в твердое кристаллическое состояние.
Дожди выпадают на поверхность океанов, морей и поверхность суши, просачиваются в почву, собираются в грунтовые воды, ручьи и реки и стекают в моря. При этом в воду поступают подвижные атомы из горных пород и из почвы и выносятся ручьями и реками в моря и океаны. Снега выпадают на поверхность, растаивают и превращаются в жидкую воду. Но если температура остается недостаточно высокой для их таяния, то снега превращаются в фирновые снежники и потом в ледники - глетчеры.
Водяной пар в верхних слоях атмосферы разлагается на кислород и водород, водород теряется в космическом пространстве, т.к. гравитация Земли не может удержать легкие атомы водорода. Восполнение этой потери происходит в результате поступления на поверхность ювенильных вод из мантии Земли. |
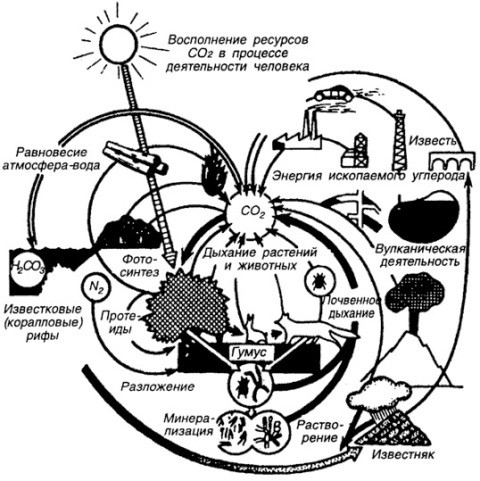
Структура круговорота углерода в биосфере Земли. Схема взята с сайта по адресу: http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
|
Углерод, пожалуй, главный химический элемент жизни на Земле. Благодаря своей валентности, равной 4, атомы углерода способны образовывать длинные цепочки молекул полимеров. Эти полимеры бывают линейные и циклические. В состав полимеров, в основе которых находятся цепочки из углеродных атомов, могут входить атомы водорода, кислорода, серы, азота.
Но живые растения при фотосинтезе используют углеод из углекислого газа, содержание которого в атмосфере Земли очень мало (по сравнению с соседними планетами - Марсом, Венерой). Только благодаря переходу на циклическое использование углерода жизнь на суше Земли в каменноугольный период не прекратилась, израсходовав почти весь углекислый газ атмосферы.
Животные, растения грибы, бактерии в результате дыхания, а также вулканы во время извержения, заводы, ТЭЦ и автомобили выделяют в атмосферу углекислый газ. Но количество его остается примерно на одном уровне, так как углекислый газ поглощают растения при фотосинтезе. Поглощает углекислый газ атмосферы и океан: когда содержание его в атмосфере начинает расти, то он переходит в растворенное состояние. В океане излишки углекислого газа взаимодействуют с водой, при этом образуется угольная кислота. Кроме того, углекислый газ, растворенный в воде океанов и морей, расходуется в процессе фотосинтеза водорослями.
В результате баланс углекислого газа в атмосфере и в океане поддерживается на низком, но приемлемом для жизни уровне как в атмосфере, та и в океанах и морях - вот уже не менее 200 миллионов лет. |
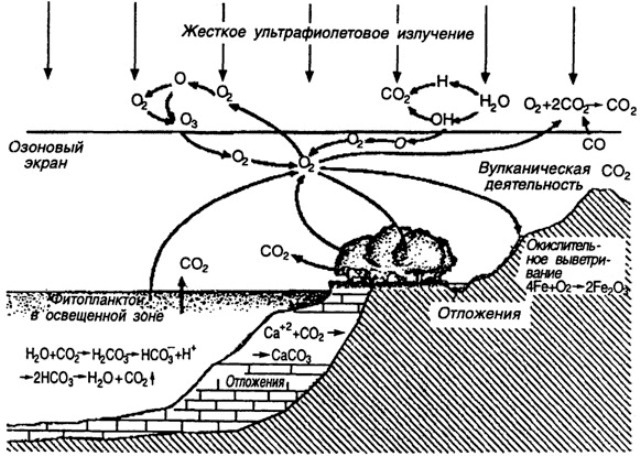
Структура круговорота кислорода в биосфере Земли
(по П. Клауду, А. Джибору, 1972). Схема взята с сайта по адресу:
http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
|
Первоначальная атмосфера Земли была практически бескислородная. Кислород - это результат работы биосферы на протяжении многих сотен миллионов лет, в значительной степени это - продукт фотосинтеза (вначале - фотосинтеза цианобактерий, затем водорослей, мохообразных,а затем высших споровых растений и, наконец, голосеменных и покрытосеменных растений).
Но кислород растениями не только выделяется - фотосинтез идет на свету, а в темноте растения дышат, потребляя кислород и выделяя углекислый газ. Просто процессы фотосинтеза у растений превалируют над процессами у них дыхания. Кислород при дыхании потребляют животные, грибы, бактерии. Запас кислорода в атмосфере такой большой, что небольшие изменения (колебания) в его содержании практически не оказывают влияния на биоту, в отличие от колебаний в содержании углекислого газа.
Углекислый газ на Земле - фактор в минимуме, а кислород - тоже фактор, но до минимума ему пока далеко. Тем не менее, если прекратить или резко сократить фотосинтез на планете (на суше и в океане), но продолжать интенсивно сжигать каменный уголь, нефть и газ, то содержание кислорода в атмосфере сократится в течение 100 лет настолько, что жителям Земли придется одеть скафандры и постоянно таскать с собой балоны с кислородом. Большое содержание в атмосфере планеты кислорода - важный признак наличия у нее активной биосферы и интенсивного фотосинтеза. |
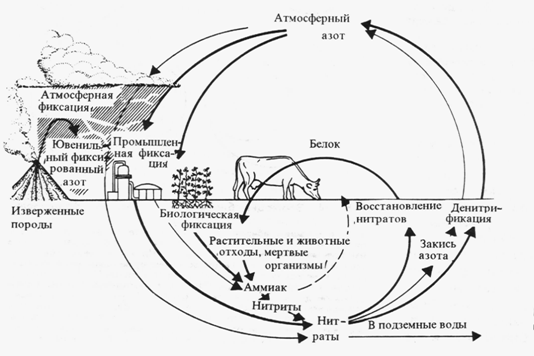
Структура круговорота азота в биосфере. Схема взята с сайта по адресу: http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
|
Главный источник азота для биоты – это молекулярный азот атмосферы. Но растения, за редким исключением, не могут извлекать его из атмосферы. Переход азота в доступные живым организмам соединения может осуществляться разными путями. Так, электрические разряды при грозах синтезируют из азота и кислорода воздуха окись азота, которая с дождевыми водами попадает в почву и образует селитру или азотную кислоту. Имеет место и фотохимическая фиксация атмосферного азота. Более важной формой усвоения азота является деятельность азот-фиксирующих микроорганизмов, которые, поглощая атмосферный азот, синтезируют сложные протеиды. Отмирая, эти микроорганизмы обогащают почву органическим азотом, который быстро минерализируется и становится доступен растениям. Таким путем в почву от этих организмов ежегодно поступает около 25 кг азота на 1 га.
Наиболее эффективная фиксация азота осуществляется бактериями, симбиотически связанными с растениями из сем. Бобовых. Образуемый ими органический азот попадает в почву, а также включается в наземные органы растений-хозяев. Таким путем в наземных и подземных органах бобовых растений на 1 га накапливается за год 150-400 кг азота.
В водной среде и на очень влажной почве непосредственную фиксацию атмосферного азота осуществляют цианобактерии. Во всех этих случаях в в почве образуются нитраты. Эти соединения через корни и проводящие пути доставляются в листья, где используются для синтеза протеинов. |
Сапрофаги, разлагающие органическое вещество, превращают органические азотсодержащие вещества в неорганические, часто в аммиак, который затем может войти в цикл нитрификации. Происходит постоянное возвращение азота в атмосферу бактерями денитрификаторами, которые разлагают нитраты с образованием молекулярного азота. Благодаря их деятельности ежегодно с 1 га почвы в атмосферу поступает до 50-60 кг азота. Азот может аккумулироваться в глубоководных осадках океана, и так выходить из биологического круговорота.
Высокое содержание азота в атмосфере Земли резко отличает ее от планет Солнечной системы. Такого количества азота на Венере и на Марсе нет. Ни растения, ни животные, ни грибы не выделяют в процессе своей жизнедеятельности свободный азот в таких количествах, в каких он содержится в атмосфере (около 70%). Так откуда же он взялся и когда накопился в таких количествах? Ответить на этот вопрос пока можно только предположительно. По всей вероятности, до того, как возникла современная биосфера Земли, в основе которой лежит фотосинтез и использование солнечной энергии, приходящей в виде светового излучения, на нашей планете была другая жизнь и другая биосфера, энергетика которой была основана на извлечении химической энергии из нитратов. Органическое вещество в той биосфере создавали организмы хемосинтетики.
Такая жизнь и сегодня на Земле есть, только она скрыта в океане на большой глубине, куда не проникает солнечный свет. Эту жизнь в 60-х годах ХХ века обнаружили вокруг подводных вулканов - так называемых черных курильщиков. По цепям питания органическое вещество, созданное микроорганизмами хемосинтетиками, передается животным, в том числе и многоклеточным, которые за его счет строят свои тела и получают необходимую энергию. Суть моей гипотезы сводится к тому, что до современной фотосинтетической биосферы на нашей планете была биосфера хемосинтетическая, и современный азот нашей атмосферы - это ее производное.
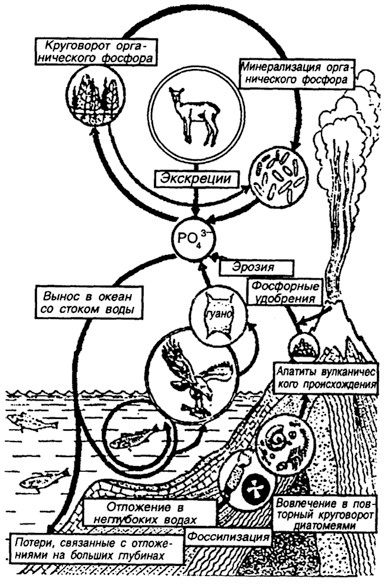
Структура круговорота фосфора в биосфере Земли (по П. Дювиньо, М. Тангу, 1973). Схема взята с сайта по адресу:
http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
|
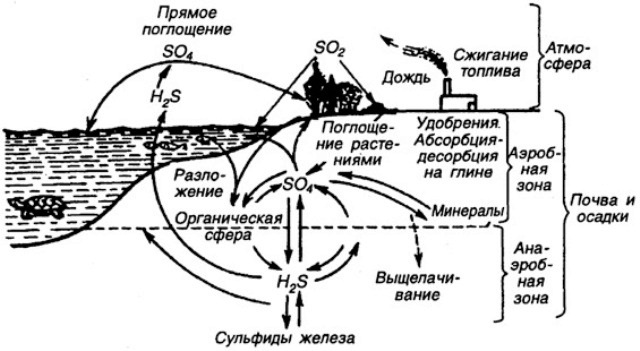
Структура круговорота серы в биосфере Земли. Схема взята с сайта по адресу: http://bibl.tikva.ru/base/B1688/B1688Part13-182.php
Фосфор и сера - также необходимые для живых организмов химические элементы минерального питания растений. Как фосфор, так и серу растения получают из почвенных растворов, которые всасывают корнями. Запас подвижного фосфора и подвижной серы в почве бывает ограниченным, поэтому для стимулирования продукционного процесса у растений серу и фосфор приходится вносить в виде минеральных или органических удобрений. А в локальных экосистемах существует их самый настоящий биологический круговорот. Но круговорот этот не полностью замкнут даже в естественных экосистемах, не говоря уж о посевах монокультур. Потеря серы экосистемой называется выщелачиванием почвы.
|
Эволюция биологического круговорота и эволюция биосферы
В мезозое началась, а в кайнозое продолжалась существенная перестройка биосферы суши, - переход ее в принципиально новое биогеохимическое состояние. Поскольку растительный покров суши превратился в своеобразную ловушку для углерода и элементов минерального (почвенного) питания, их содержание в доступной растениям форме во внешней среде снизилось. Продуктивность растительного покрова в связи с этим понизилась. Стратегия развития биосферы суши в мезозое и кайнозое состояла в том, чтобы обеспечить цикличность биогеохимических превращений элементов в биологическом круговороте, максимально уравновесив этот круговорот с термодинамическим потенциалом местообитаний. А это было возможно только за счет развития нисходящей ветви биологического круговорота - гнилостных бактерий, грибов, животных питающихся мертвой органикой.
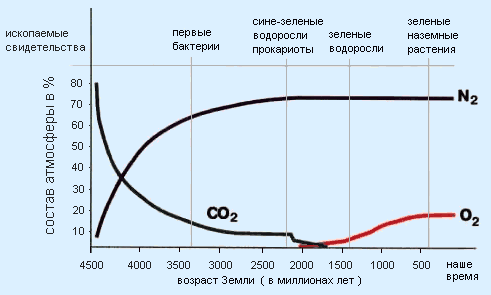
- Изменение содержания азота, углекислого газа и кислорода в атмосфере Земли за 4,5 млрд. лет. Схема с сайта:
- http://znaniya-sila.narod.ru/solarsis/zemlya/earth_05.htm
Из этой схемы следует, что в течение промежутка времени с 4500 млн. лет назад по 2200 млн. лет назад в биосфере Земли источником органического вещества и энергии был хемосинтез, атмосфера в то время практически не содержала кислорода. Фотосинтез возник около 2200 млн. лет назад, а примерно 1000 млн. лет назад стала возможна жизнь на суше. Разумеется, эти временные границы весьма приблизительны.
|
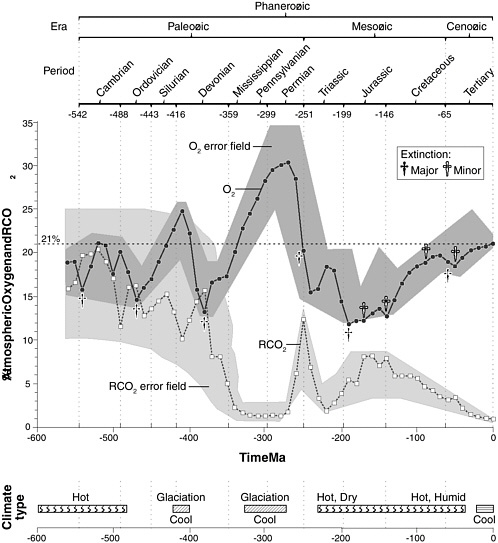
Изменение содержания в атмосфере кислорода и углекислого газа в фанерозое за 600 млн. лет. Схема с сайта: http://forum.ixbt.com/post.cgi?id=print:54:52103&page=5
По всей вероятности, содержание углекислого газа в атмосфере Земли на этой схеме завышено, особенно в первой половине фанерозоя. При столь высоком содержании углекислого газа оледенения были бы невозможны из-за парникового эффекта.
|
Параметры биологического круговорота
В качестве основного принципа организации и эволюции биосферы суши необходимо рассматривать постоянную тенденцию к увеличению объема и к ускорению биологического круговорота химических элементов в биосфере. Интенсификация биогеохимических процессов достигается самыми различными путями: дискретностью жизни, увеличением разнообразия форм живых организмов и их сообществ, совершенствованием фотосинтезирующего аппарата и совершенствованием физиологии организмов гетеротрофов.
Увеличение разнообразия форм жизни обеспечивало включение в биогеохимический процесс самых разных местообитаний, а дискретность жизни на уровне особей обеспечивала ее быстрое ''растекание'' по поверхности Земли и возможность сочетания в одних биоценозах организмов с самыми разными свойствами.

Влажный тропический лес на берегах Амазонки. Фото с сайта: http://gumarchek.ru/?eef=pictures-of-plates-of-the-rainforest
|
Растения эпифиты (укореняющиеся на стволах и ветвях деревьев) обеспечивают быстрое включение вещества в новый цикл круговорота. Высвобождающиеся при разрушении коры и древесины деревьев атомы, минуя их поступления в почву, включаются эпифитами в новый цикл. Эпифитов больше там, где влажнее климат, где больше выпадает осадков и где больше вероятность выноса химических элементов из почвы за пределы конкретной экосистемы (Галанин, 1990).
На севере эпифитами являются мхи, печеночники и лишайники, а на юге к ним добавляются высшие сосудистые растения - травянистые орхидеи и даже древесные (кустарники).
Возникновение в результате эволюции растений лиан можно объяснить тем, что сообщество, состоящее только из крупных деревьев, не может включить в биологический круговорот все подвижные химические элементы, имеющиеся в почве. Однако этих, остающихся невключенными в биологический круговорот, элементов недостаточно для роста деревьев. Кустарники и травы, имея малый габитус, могли бы реализовать этот остаточный эдафический потенциал, но их развитие под пологом деревьев лимитировано малыми ресурсами световой энергии. За миллионы лет эволюции в таких экосистемах сформировался новый тип жизненной форы растений - древовидные лианы.
|
Лианами, имеющими тонкие стволы, значительная ассимилирующая поверхность листьев выносится на уровень древесного полога, но достигается это минимальными вещественными затратами на формирование мощного ствола. А ведь вещество, сосредоточенное в стволах деревьев, в ежегодном биологическом круговороте не участвует. Стволы деревьев функционируют десятки и сотни лет, и только после смерти дерева падают на землю и разлагаются сапрофитами.

Лиана в тропическом лесу.
|
В древесине стволов деревьев и кустарников сосредоточена большая масса химических элементов, «выключенных» из ежегодного обращения в биологическом круговороте. Но при наличии в сообществе лиан биогеохимическая работа экосистемой производится при меньших «вещественных затратах», т.е. более оптимально.
По мере эволюции экосистем в них, вероятно, при увеличении интенсивности продукционного процесса происходило снижение запаса биомассы на единицу биогеохимической работы, поэтому отношение скорости накопления фитомассы к запасу биомассы экосистемы является одной из самых важных характеристик биологического круговорота вещества. Это отношение можно назвать коэффициентом оборачиваемости вещества в биологическом круговороте в конкретной экосистеме.
Принцип интенсификации биологического круговорота наиболее ярко проявляется в экологической роли травоядных животных. Растительная масса, накопленная автотрофами, идет в пищу млекопитающим, которые своими экскрементами и трупами удобряют почву и тем самым ускоряют продукционный процесс у растений и способствуют повторному использованию атомов вещества в течение одного вегетационного периода.
Исследуя автотрофный цикл биологического круговорота, Р.И.Злотин и К.С.Ходашова (1973) пришли к выводу, что основная роль животных в экосистеме заключается не в рассеивании энергии, накопленной продуцентами, а в участии в процессах биологического круговорота, прежде всего во влиянии на его автотрофный цикл.
При участии животных консументов интенсификация круговорота происходит за счет ускорения биогеохимических процессов, а не за счет увеличения количества вещества в потоке круговорота. |
Опубликованные материалы (Информация с сайта: http://mlekopitajuschie.ru/travojadnye-mlekopitajushchie-v-aridnyh-ekosistemah-vnetropicheskoi-evrazii/ocenka-roli-travojadnyh-zhivotnyh-v-biologicheskom-krugovorote.html) позволяют в общих чертах представить размеры потребления и переработки разными млекопитающими травянистой растительности. В мочевине, например, содержится гораздо больше азота — важного биогена, чем в кале. Полевка выделяет с калом 26% поглощенного азота, а с мочой 74%. Азот мочевины почти не требует редуцирующей деятельности микроорганизмов и находится в готовом или почти готовом виде для усваивания растениями. Концентрация мочевины в моче больших песчанок в естественных условиях зимой равна в среднем 33,5 мг в 1 мл мочи, с колебаниями от 16,5 до 72,7 мг/мл (Соколов, Скурат, 1962). Большая песчанка летом за сутки съедает в среднем 79 г сырой массы корма и выделяет 6-12 г сырого помета (Лобачев, Хамдамова, 1972), Суточное потребление большой песчанкой воздушно-сухого корма, по данным разных авторов (Гладкина, Мокеева, 1958; Абатуров, 1979), колеблется от 7,6 до 26 г. А. И. Щеглова (1976) указывает, что этот вид за сутки выделяет с мочой 63% общего количества поступающей с пищей воды. Следовательно суточный диурез большой песчанки в весеннелетнее время лежит в пределах примерно 30 мл. По материалам Т. А. Середневой и A. Незговорова (1977а, б), взрослый байбак за сутки съедает 723 г. сырой зеленой массы, годовалый сурок — 430 г, сеголеток — 299 г. Следовательно, взрослый байбак выделяет за сутки около 600 г., годовалый — более 350 г. и сеголеток — около 250 г. жидкости за сутки. Сибирский лемминг за сутки съедает 92 г. сырой массы растений (Токмакова, 1975, 1982), следовательно, количество выводимой им жидкости, преимущественно мочи, составляет около 80 г., что составляет около 200% живой массы зверька. Овца в загоне съедала 4,2 кг сырой растительной массы в сутки, а на пастбище — 8 кг (Слоним, 1962). Ппри содержании в клетке малого суслика потребление корма составляет 12,5 г/сутки сухого веса против 30 г/сутки, поедаемых в естественных условиях. При изучении потребности в пище малого суслика в клетке Б. Д. Абатуров и Т. А. Середнева (1973) получили суточное потребление одной особью, равное 37 г воздушно-сухого веса. Годовалые сайгаки при содержании в клетке в мае и июне потребляли от 1,6 до 3,9 кг сырой растительной массы травянистых растений (Абатуров и др.,1973). В июне - августе лоси съедали 30-40 кг сырой растительной массы, в октябре - декабре —15-20, в апреле — 6-12 кг в день. (Кнорре, Кнорре, 1959).
Обилие и доминирование растительноядных млекопитающих (копытных и грызунов) в аридных экосистемах подчеркивается всеми авторами, специально занимавшимися их животным миром. К степям и пустыням, видимо, приурочено возникновение животноводства, которое весьма долгий срок было здесь основной формой использования природных ресурсов. Все авторы, занимавшиеся влиянием копытных на фитоценозы аридных экосистем, подчеркивают, что умеренный выпас есть одно из необходимых условий существования здесь нормально развитого растительного покрова (Высоцкий, 1901; Пачосский, 1917 и др.). Полезное влияние зеленоядных млекопитающих на растительность не ограничивается уничтожением мертвых остатков. Одно из приспособлений травянистых растений к постоянному воздействию на них животных — это возникновение отавности. Отавностью называется способность растений восстанавливать более или менее энергично и полно массу надземных частей, отчуждаемых в результате поедания животными или при скашивании (Вощинин, 1935).
Повторное отрастание растений в течение вегетационного сезона после скусывания их кормящимися животными может дать значительное увеличение суммарного урожая растительной массы с единицы площади. На степных пастбищах, когда при выпасе поедается не более 50% доступного корма, и в дальнейшем (в следующем году) не наблюдается снижения урожайности растений. Наоборот, урожай неоднократно поедаемой скотом за вегетационный сезон воздушно-сухой массы, по сравнению с извлекаемой с луга единожды за сезон при сенокошении, увеличивается в 2-5 раз (Евсеев, 1954). Пасущиеся животные поедают и разбивают копытами отмершие части растений, чем также ускоряют биологический круговорот. Однако при чрезмерном выпасе животных изменяются режим влажности и степень освещенности поверхности почвы, что затрудняет семенное возобновление, препятствует нормальному развитию плотнокустовых злаков и угнетает фотосинтез. При отсутствии же выпаса вес мертвых растительных остатков часто в несколько раз превышает размеры годового прироста растительной массы. На старом заповедном участке дерновинно-злаковой степи в Аскании-Нова мертвые остатки составляли в среднем 84% веса воздушно-сухой надземной массы растений с колебаниями по отдельным участкам от 68 до 94% (Шалыт, 1950).

Стадо коров и степь – это экосистема с автономным круговоротом вещества. Фото с сайта: http://fotki.yandex.ru/users/tanysmirnowa/view/622616/
|
Но не только крупные млекопитающие могут активно участвовать в биологическом круговороте вещества. Так, полевки при численности 100 особей на 1 га. отчуждают около 4% чистой первичной продукции лугового растительного покрова (Давыдов, 1978). При этом с экскрементами в среднем за 1 сутки на 1 га от них поступает около 30 г азота, 2 г фосфора и 40 г калия. Это способствует неоднократной реассимиляции элементов в течение одного вегетационного сезона.
Не только травоядные млекопитающие играют большую роль в ускорении биологического круговорота вещества на Земле. Насекомые в этом ускорении играют порой не меньшую, а иногда и большую роль. Здесь следует отметить листогрызущих гусениц самых разных видов насекомых, чья роль в отчуждении фитомассы очень велика, а также роль грибов паразитов, часто поселяющихся на листьях живых растений. Достаточно вспомнить нашествия на сибирские леса непарного шелкопряда, начисто выедающего хвою лиственницы, листья березы, или полчища саранчи, за сутки съедающие всю зеленую фитомассу на своем пути. |

- Гусеницы листоеды выгрызли значительную часть листовых пластинок. Но надо помнить, что их трупы удобрили почву и стимулировали рост растений. Фото с сайта:
- http://forum.tvoysad.ru/vrediteli-i-bolezni-rasteniy-f40/abrikos-zabolel-pomogite-t6297.html
|

- Жирная гусеница за свою короткую жизнь съедает немало листьев и тем самым принимает самое активное участие в биологическом круговороте. Фото с сайта:
- http://www.mayfordpond.org.uk/gallery.htm
Не зря говорят, что Земля - это планета растений и насекомых. Именно с ними мы встречаемся на каждом шагу и в большом количестве.
|

- Паразитический вид грибов вызывает поражение пасленовых, называемое фитофторой. Фитофтора отчуждает живую фитомассу и возвращает вещество, потребленное растением во внешнюю среду, откуда оно снова в тот же вегетационный период вовлекается в новый цикл биологических превращений. Фото с сайта:
- http://www.vsedlyasada.ru/ovoschi/tomaty/bolezni-tomata-i-borba-s-nimi/
|

- Болезнь фитофтора поражает листья, стебли и зеленые плоды. На этом фото видно, что таким образом возвращается в новый цикл биологического круговорота примерно половина вещества, накопленного за лето растениями. Фото с сайта:
- http://tlttimes.tltsu.ru/blog/animals/979.html
|

- Так выглядит нашествие саранчи. Фото с сайта:
- http://vasily-sergeev.livejournal.com/2394283.html
|

- Вот это зеленое существо и есть гигантская саранча. Фото с сайта:
- http://lenta.mk.ua/article/835596.html
|

Нашествие саранчи в Астраханской области. Фото с сайта: http://yablor.ru/blogs/nashestvie-saranchi-astrahan/1768143
|
Нашествия саранчи - страшное бедствие растениевода. Размножившиеся полчища этих прожорливых насекомых обрушиваются на поля, огороды и сады, съедая зеленые листья и оставляя крестьян без урожая. За 1-2 часа саранча съедает огромное количество фитомассы и, умерев с голода, в конце концов становится удобрением, успев при этом отложить яйца - т.е. оставить потомство.
Саранча может летать, поэтому быстро передвигается с одного поля на другое.
Причина массовых нашествий саранчи в том, что созданные человеком аграрные экосистемы с экологической точки зрения несовершенны. В них разрушены механизмы естественной регуляции численности организмов. Если в естественных равновесных биоценозах численность каждого вида контролируется другим видом, для которого особи этого вида служат пищей, то в сообществах и экосистемах неравновесных возможны вспышки численности одного какого-то вида такие, что он может уничтожить данное сообщество и экосистему. В естественных, хорошо сформированных экосистемах существуют обратные связи, которые обеспечивают регулирование численности одних видов другими. Если в равновесной системе увеличивается численность саранчи, то сразу же начинает расти численность ее врага, который питается саранчой. Но когда сокращается численность саранчи, то снижается и численность ее врага, т.к. поймать редких особей саранчи ему становится не так-то просто.
Массовые нашествия саранчи говорят только об одном - экологическом несовершенстве создаваемых искусственных экосистем. |
Известны нашествия еще одного массового вредителя, на сей раз вредителя лесов - непарного сибирского шелкопряда. Хвою лиственницы, мелколиственных и широколиственных древесных пород поедают прожорливые гусеницы симпатичной бабочки.

- Гусеницы непраного шелкопряда поедают хвою лиственницы, отчего полог в таких лесах становится сильно разреженный, что благоприятствует росту трав. А вот деревья при этом резко снижают свою продуктивность. Фото с сайта:
- http://knifeclub.com.ua/forum/viewtopic.php?f=73&t=20937
|

- Бабочки розового непарного шелкопряда (Lymantria mathura). Осенью бабочки откладывают яйца, которые зимуют, и весной из них выводятся прожорливые гусеницы, которые по стволам, сучьям и веточкам ползут в кроны деревьев. Фото с сайта: http://macroid.ru/showphoto.php?photo=23941&size=big&cat=63699
-
|

- Лиственичный лес, пораженный непарным шелкопрядом. Хвоя на лиственницах гусеницами съедена полностью. Словно пожар прошел по этому лесу. Фото с сайта:
- http://www.ursn10.ru/info/consultation/article40.html
|

- Яйцекладки непарного шелкопряда на березе неплохо укрыты от холода. Фото с сайта:
- http://botgard.uran.ru/node/41
|
Непарный шелкопряд в природе существует также как фактор ускорения биологического круговорота и регулирования видового состава древостоев. Когда сообщество деревьев становится биогеохимически не оптимальным, а вид лесообразователь оказывается далеко не в оптимуме в изменившихся условиях, то для быстрой перестройки экосистемы требуется быстрая эллиминация этого вида. Эту роль в природе и выполняют массовые насекомые. Удалив хвою и листья деревьев, они тем самым стимулируют рост трав под пологом леса, и такая лесная экосистема начинает работать как экосистема луга или степи. Так же как в случае с нашествиями саранчи, нашествия непарного шелкопряда часто случаются в лесах вблизи полей, они поражают небольшие массивы леса, но редко случаются в девственных лесах, так как там сохранились виды животных регуляторов численности шелкопряда, которые активно поедают гусениц шелкопряда.
Эдафический потенциал дождевого тропического леса очень велик, объем вещества, находящегося в этих экосистемах в биологическом круговороте, огромный и скорость оборота этого вещества, по сравнению с другими экосистемами, очень большая. Здесь в биологический круговорот вовлечено очень много атомов углерода, кислорода, водорода, азота, серы, кальция, калия, натрия, меди, железа и других химических элементов. Многоярусная растительность в тропическом лесу улавливает много солнечной энергии. По многочисленным цепям питания вещество и энергия распределяются в синузии животных и грибов, через ценопопуляции грибов, животных и микроорганизмов вещество снова возвращается во внешнюю среду - в почву и атмосферу, а из них включается в новый цикл биологического круговорота, не покидая локальную экосистему, не уходя за ее пределы.

- Восточный берег Мадагаскара порос тропическим дождевым лесом. Под полог проникает мало света, отчего напочвенный покров здесь не развит. Фото с сайта: http://www.otdohnuli.ru/stories/1376
|

- Тропические леса Амазонки насыщены разнообразными видами растений и животных. Фото с сайта:
- http://katyaburg.ru/razdely/krasota-prirody/tropicheskie-lesa-amazonki
|

- Тропический дождевой лес. Отметим сплошное покрытие стволов эпифитами. Фото с сайта:
- http://lori.ru/1466154
|

- Многоярусный тропический дождевой лес в бассейне Амазонки. Фото с сайта:
- http://youroute.ru/places/16310
|
В локальных экосистемах тропического леса биологический круговорот замкнут для многих химических элементов. Эти элементы удерживаются в круговороте благодаря сложной биоценотической системе, работающей слаженно и надежно. Но когда тропический лес вырубают, древесину вывозят, порубочные остатки сгнивают и вещество, образующее при их гниении, быстро вымывается дождевыми водами. Прежний тропический лес на этом месте больше не возобновляется, и это связано с резким снижением эдафического потенциала местообитания. Почвы в тропических дождевых лесах не плодородны, резерв вещества в них находится не в почве, а в наземной фитомассе. Девственный тропический лес буквально не выпускает химические элементы из круговорота: в его экосистемевсе атомы из одного цикла тут же поступают в новый цикл, и потому не успевают вымываться ни в грунтовые воды, ни в реки. Циклы вещества здесь почти замкнуты, но стоит разрушить такое сообщество, как все вещество, ранее участвовавшее в круговороте, вымывается за пределы экосистемы.

Лиственичное редколесье в Колымском нагорье в Магаданской области. Фото А.А. Галанина.
|
Но большой запас наземной фитомассы в экосистеме далеко не всегда свидетельствует о большой скорости биологического круговорота и о высокой скорости накопления этой биомассы. Например, в Магаданской области в 30-40 годы лесозаготовители изъяли большой запас древесины в лиственничных лесах, считая, что это весьма продуктивные экосистемы (раз в лесах такие большие деревья росли).
Но лесозаготовители просчитались. Через 40 лет интенсивных рубок почти все лиственничные леса в Магаданской области были вырублены, но возобновилось за это время древесины едва 10% от вырубленного ее количества. Биологический круговорот в северных лесах чрезвычайно медленный, хотя объем вещества, находящегося в этом круговороте, здесь может быть довольно большой. Вообще в лесоведении есть понятие расчетная лесосека, которая должна быть равна такому количеству вырубаемой за год древесины, которое в этих лесах прирастает за вегетационный период. В Магаданской области расчетная лесосека в свое время была рассчитана неправильно или намеренно завышена.
Все хорошо знают, что чем медленнее течение реки, тем больше и глубже ее плеса, а на перекатах, где течение быстрое, там очень мелко. Моделируя биологический круговорот, можно сравнить его с рекой, только течет эта река множеством проток, и течение в каждой из них разное: то очень быстрое, то очень медленное. |
Биологическое разнообразие как фактор оптимизации экосистем
Разнообразие биотического покрова – основное условие его оптимального функционирования в биосфере. Главными механизмами разнообразия несомненно являются: дискретность жизни на уровне особей, способность особей к размножению и передача наследственной информации от предыдущего поколения следующему, устойчивость наследственного вещества (ДНК и РНК) к воздействию разных факторов в процессе онтогенеза особей, возможность появления и накопления в естественных популяциях растений мутаций в результате воздействия на их ДНК и РНК жестких излучений, резких химических и температурных воздействий.
В стабильных условиях особи-мутанты имеют мало шансов выжить (в стабильных экосистемах действует так называемый стабилизирующий естественный отбор), но когда условия существования резко изменятся, то шансов выжить и оставить потомство у мутантов становится больше, особенно у тех, у которых мутации не вызвали дисбаланс биогеохомических процессов в организме.
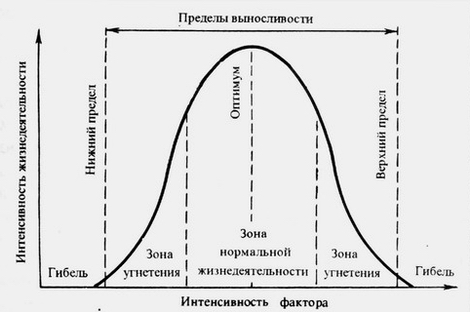
Диаграмма экологической амплитуды вида по фактору Х (ось абсцисс). За пределами зон угнетения находятся пределы толерантности вида.
|
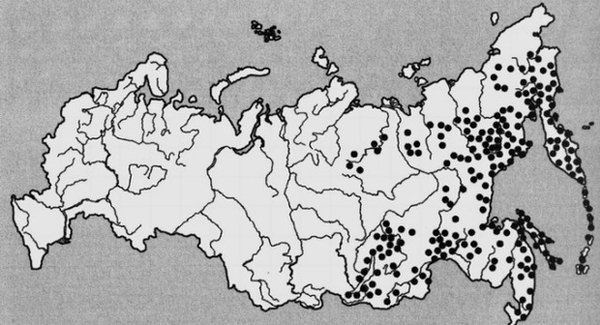
Ареал распространения кедрового стланника на территории России.
|
Диплоидность и разделение полов – это тоже механизм, обеспечивающий сохранение биоразнообразия и способствующий его формированию. При половом размножении в популяциях диплоидных организмов в аллелях в рецессивных генах могут накапливаться мутации, которые в фенотипе не проявляются и стабилизирующим отбором не отбрасываются. Генофонд популяции диплоидных организмов в рецессивных генах хранит резерв нового генетического материала на случай резкого изменения условий существования популяции. В этих случаях мутировавшие гены из рецессивных могут стать доминантными.
В популяциях гаплоидных организмов такое накапливание мутировавших генов невозможно, все мутации из популяции отметает стабилизирующий отбор, поэтому скорость видообразования у гаплоидов (мохообразные) на 2-3 порядка ниже, чем у диплоидов. У гаплоидов весь генетический материал контролируется стабилизирующим отбором, а у диплоидов есть некоторый резерв мутировавших генов.
Важным механизмом биоразнообразия является способность живых существ к миграциям. Когда изменяются условия существования вида в пределах какой-то части его географического ареала, когда условия оказываются за пределами толерантности вида, то вид в данной местности вымирает. Однако он сохраняется в другой части своего географического ареала - там, где условия остались в пределах его толерантности. Кроме того, вид при этом расселяется в новые районы, условия в которых стали пригодны для существования особей данного вида.
Благодаря способности к миграции в новые районы виды получают шанс пережить резкую смену условий, уйдя из прежних районов и мест обитания в новые, ставшие для них пригодными. Так происходит обогащение флор и фаун новыми видами, так происходят изменения сообществ живых организмов и экосистем.
Миграции растений обеспечиваются специальными приспособлениями плодов и семян к переносу ветром, водными потоками и животными. Все эти механизмы распространения семян и плодов выработались в процессе длительной эволюции, а в случае переноса семян и плодов животными - в результате сопряженной эволюции растений и животных. Парашютики семян одуванчика и других сложноцветных, и не только сложноцветных, прицепки, с помощью которых семена и плоды других растений цепляются за шерсть млекопитающих - все эти свойства у растений возникли как приспособления к распространению семян и плодов.
Далеко не все виды могут разделяться на дочерние или трансформироваться в новые виды. Очень часто при резких климатических изменениях или иных изменениях экологических условий виды вымирают, так как не могут найти себе подходящих местообитаний в перестроившейся биосфере. Такие виды не могут перестраиваться на прежнем месте, но они не могут в силу тех или иных причин и откочевать на новые места. Сначала они становятся редкими, а затем и вовсе вымирают. В геологической истории Земли известны эпохи, когда вымирание видов носило просто катастрофический характер. В эти эпохи происходили мощные перестройки всей биосферы. Так в свое время в конце мезозойской эры в меловом периоде вымерли многие виды пресмыкающихся, например, динозавры, и многие виды голосеменных растений, а еще раньше - в начале мезозоя - вымерли или почти все виды гигантских папоротников. Всего 5-10 тыс. лет назад на Земле вымерли последние мамонты, около 200 лет назад из биосферы Земли исчезла Стеллерова корова - вид морских млекопитающих, обитавший в морях северной Пацифики, на острове Мадагаскар исчезла гигантская нелетающая птица, ставшая прообразом птицы рух в арабских сказках про Синбада-морехода. А многие виды исчезли с лица Земли и вовсе незнаемые людьми, т.к. ученые не успели их обнаружить в природе и дать им даже названия.
Процессы вымирания одних видов в биосфере компенсируются появлением новых видов в результате процессов видообразования. Если баланс этих двух процессов резко нарушится в пользу вымирания, то Землю, скорее всего, ждет судьба Венеры. Атмосфера Земли вернется к прежнему состоянию, когда в ней было велико содержание углекислого газа, метана и паров воды, при этом резко снизится содержание кислорода. Температура на поверхности и в приземном слое атмосферы тогда повысится примерно до +200°С, закипят и испарятся океаны и моря. Жизнь на белковой основе в таких условиях, разумеется, станет просто невозможна. Человечество обязано взять на себя ответственность не только за будущее своих детей и внуков, но и за будущее самой биосферы, и это будущее во многом будет зависеть от того, насколько процесс вымирания видов в биосфере Земли отстает от процесса образования новых видов.
Процессы видообразвания протекают очень медленно. Так, например, для разделения родительского вида сосудистых растений на два дочерних вида, при наличии между двумя популяциями барьера, не позволяющего особям этих двух популяций скрещиваться друг с другом, природе потребуется как минимум 500 тыс. лет, а чаще около 1 миллиона лет. Отдельные виды в биосфере могут сохраняться 10 и более миллионов лет, практически не изменяясь за это время.
Разнообразие видов и разнообразие жизненных форм или экобиоморф - это далеко не одно и то же. Продемонстрирую это на таком примере. На лугу видов, родов и семейств растений может обитать в 2-3 раза больше, чем в темнохвойном лесу. Однако в пересчете на экобиоморфы и типы синузий окажется, что биоразнообразие темнохвойного леса как экосистемы значительно выше, чем биоразнообразие луга как экосистемы. На лугу мы имеем 2-3 класса экобиоморф (травы однолетние, травы многолетние длиннокорневищные, травы многолетние дерновинные), а в темнохвойном лесу 8-11 классов (деревья хвойные первого яруса, деревья широколиственные второго яруса, кустарники, лианы, травы длиннокорневищные, травы дерновинные, мхи зеленые, лишайники кустистые эпигейные, лишайники эпифиты кустистые, лишайники эпифиты накипные). На лугу видов много, но все они относятся к небольшому числу классов экобиоморф. В лесу же видов может быть меньше, но разнообразие классов биоморф выше. Если сходные морфологически ценопопуляции видов выполняют сходную биогеохимическую работу - сходно участвуют в биологическом круговороте, то разные жизненные формы и разные синузии выполняют разную биогеохимическую работу - по-разному участвуют в биологическом круговороте.
Биоразнообразие организмов в биосфере не исчерпывается разнообразием таксонов и разнообразием экобиоморф живых организмов. Например, мы можем попасть в район, который целиком занят одной локальной элементарной экосистемой - верховым болотом или сырым ольховым лесом в устье большой реки. В другом районе на такой же по площади территории мы встретим не менее 10-15 типов локальных элементарных экосистем. Экосистемы хвойно-широколиственного леса на дне долин рек закономерно сменяются здесь экосистемами кедрово-дубовых разнотарвно-кустарниковых лесов на южных пологих склонах гор, лиственично-дубовыми разнотравными лесами на северных пологих склонах гор, елово-пихтовыми лесами в верхней части северных крутых склонов гор и экосистемами остепненных лугов и куртинной растительности на крутых южных склонах гор. Нетрудно понять, что такое внутриландшафтное разнообразие экосистем определяется не только разнообразием слагающих их видов и экобиоморф, но и разнообразием экологического фона ландшафта, связанного в первую очередь с разнообразием форм рельефа, разнообразием почв и подстилающих их горных пород.
Таким образом, фактором устойчивости локальной экосистемы и фактором ее биогеохимической оптимальности является не только разнообразие видов, обитающих в этой локальной экосистеме, но и разнообразие видов в соседних экосистемах, из которых возможен занос диазачатков (семян и спор). Сказанное относится не только к растениям, ведущим прикрепленный образ жизни, но в еще большей степени к животным, могущим перемещаться из одной локальной экосистемы в другую. Многие особи животных, не принадлежа конкретно ни к одной из локальных экосистем (биогеоценозов), тем не менее играют важную экологическую роль, участвуя в обеспечении биологического круговорота сразу в нескольких экосистемах. Мало того, они могут в одной локальной экосистеме отчуждать биомассу (кормиться в ней), а в другой выбрасывать экскременты, стимулируя таким образом рост и развитие растений в этой второй локальной экосистеме за счет вещества из первой. Порой такой перенос вещества и энергии из одних экосистем в другие может быть чрезвычайно мощным. Этот поток связывает между собой совершенно разные экосистемы. Например, проходные лососевые рыбы осуществляют поток вещества из морей в верховья рек, при этом вещество, вынесенное текучими водами из экосистем суши, возвращается назад в эти экосистемы. Выметав икру, рыбы гибнут и растаскиваются животными далеко за пределы речных пойм, пойманные животными, они поедаются ими, а экскременты оказываются выброшенными почти на водоразделах, становясь удобрением в лесах и на лугах.
Общий генофонд растительного покрова ландшафтного района (его флора) локальными экосистемами этого района используется наиболее полно благодаря давлению биоразнообразия. При этом локальные экосистемы за счет этого генетического фонда в видовом отношении становятся более богатыми. При их формировании и перестройках экологический подбор подходящих компонентов осуществляется из большего количества претендентов, диазачатки (семена, споры) которых попали в данное местообитание. Таким образом, вероятность формирования экологически оптимального растительного сообщества при этом увеличивается, увеличивается и вероятность формирования биогеохимически оптимальных локальных экосистем.
ЛИТЕРАТУРА
Бондарев Л. Г. Вечное движение. Планетарное перемещение вещества и человек. М.: Мысль, 1974. 158 с.
Вернадский В.И. Живое вещество. М.: Наука, 1978. 358 с.
Виноградов А. П. Введение в геохимию океана. М.: Наука, 1967. 216 с.
Галанин А. В. Принципы организации растительного покрова // Вестн. ДВО АН СССР, 1990. № 2. С. 121-139.
Григорьев А. А Основы теории физико-географического процесса // Тр. 2-го Всесоюз. геогр. съезда. М.: Изд-во АН СССР, 1948. С. 11-12.
Колесников С.И. Экология. – Ростов на Дону: «Феникс», 2003.
Марков К. К., Добродеев О. П., Симонов Ю. Г., Суетова И. А. Введение в физическую географию. М.: Высш. шк., 1978. 191 с.
Николайкин Н.И. Экология.: Учеб. для вузов/ Николайкин Н.Н., Николайкина Н.Е., Мелехина О.П. – 2-е изд., перераб. и доп.- М.: Дрофа, 2003.
Петров К.М. Общая экология: Взаимодействие общества и природы: Учебн. пособие. 2-е изд.- СПб.; Химия, 1998.
Хотунцев Ю.Л. Экология и экологическая безопасность: Учеб. пособие для студ. высш. пед. учеб. заведений. – М.: Издательский центр «Академия», 2002.
Шилов И.А. Экология: Учеб. для биол. и мед. спец. вузов И.А. Шилов.-4-е изд., испр.- М.: Высшая школа, 2003.
Читать конец: Лекция 2. Экологические кризисы и их причины


