УДК 581.5+581.524(576.16)
ЭКОЛОГО-ЦЕНОТИЧЕСКИЕ ЭЛЕМЕНТЫ КОНКРЕТНОЙ ФЛОРЫ
(их выделение и анализ)
А. В. Галанин © 2010
A.V. G A L A N I N. ECOLOGO-COENOBIOTIC ELEMENTS OF CONCRET FLORA
(THEIR DISTINGUISHING AND ANALYSIS)
© OCR - А.В. Галанин, 2010. Воспроизводится по тексту: Галанин А.В. ЭКОЛОГО-ЦЕНОТИЧЕСКИЕ ЭЛЕМЕНТЫ КОНКРЕТНОЙ ФЛОРЫ (их выделение и анализ) // БОТАНИЧЕСКИЙ ЖУРНАЛ 1973, ТОМ 58, №11 . С.1608-1918.
Эта работа - часть моей кандидатской диссертации. В то время "эколого-ценотические элементы флоры" не выделялись при флористических исследованиях, такого понятия просто не существовало. Флора понималась ботаниками как список видов некоей территории, причем этой территорией мог быть весь СССР, Европа, остров Врангеля, территория любой области или административного района. А вот всякая оценка обилия вида на этой территории, его распределения по типам экотопов (местообитаний) были прерогативой геоботаники и к флористике отношения не имели. Впервые поломать такие представления решил мой научный руководитель Борис Александрович Юрцев при изучении флоры хребта Сунтар-Хаята на стыке Хабаровского края, Магаданской области и Якутии. Его монография "Флора Сунтар-Хаята" вышла в 1968 г. Эколого-ценотические группы видов во флоре Сунтар-Хаята Б.А. Юрцев выделял визуально (интуитивно) на основании наблюдения поведения вида в ландшафте, деление было весьма приблизителиным. Доказательства того, что такие группы видов существуют в действительности, что они не есть фантазия исследователя Борис Александрович не сделал. Это он поручил мне своему аспиранту, и сделать это я должен был на примере конкретных флор на Чукотке за три года аспирантуры. И я это сделал. Доказательством тому служит защита кандидатской диссертации в 1974 г. на диссертационном совете в Ботаническом институте в Ленинграде.
Статья, которую сейчас предлагаю как страничку в интернет, была моей первой статьей. Тогда не было компьютеров, и даже калькуляторов. Все расчеты я делал с помощью логарифмической линейки. Работа была адова, таким образом мне пришлось рассчитать квадратную матрицу межвидовой сопряженности (корреляции по альтернативным признакам) для 250 видов. Таблица была склеена из листов миллиметровой бумаги и вывешена на стену в комнате в частном доме в Паргалове, которую я тогда снимал. Она была от пола до потолка и длиной во всю стену. Считал я ее 5 месяцев. При защите диссертации мой оппонент В.И. Василевич сказал, что матрицы такого размера на тогдашних ЭВМ рассчитать было невозможно, что я выполнил работу ЭВМ следующего поколения. Да, сегодня такую матрицу самый заурядный ПК рассчитает за 2-3 минуты. Но это было 40 лет назад.
Как сейчас помню как я отстаивал некоторые термины в редакции Ботанического журнала. Редактор хотел заменить их более привычными и понятными для него терминами. Особенно ему не нравился термин "граф" да еще эколого-ценотической структуры флоры. "Всех графов мы выгнали сразу после революции" - говорил он мне - " а ты тут их опять выкопал, да еще в наш журнал тащишь". Только после того, как я сказал, что в таком случае я заберу из журнала свою статью, и показал ему монографию "Теория графов", редактор пропустил в Ботанический журнал термин "граф". Итак, статья: БОТАНИЧЕСКИЙ ЖУРНАЛ 1973, ТОМ 58, №11 . С.1608-1918.
Выявление эколого-ценотических элементов производилось путём расчета межвидовых сопряжений по альтернативным признакам. В одну эколого-ценотическую группу объединены виды, имеющие приблизително равную встречаемость, экологические амплитуды которых в значительной степени совпадают. Результаты работы показали наличие высокой экологической индивидуальности видов. Между географическим составом всей флоры и составом отдельных эколого-ценотических элементов существует некоторое несоответствие .
Настоящее исследование было проведено летом 1970 г. в одном из центральных районов Чукотского полуострова (среднее течение р. Эргувеем). Район Эргувеем расположен в пределах Охотско-Чукотского вулканогенного пояса (фото 1, карта 1). Сопочные массивы высотой 400 – 800 м сочетаются здесь с обширные пространствами межгорных дегрессий, в пределах которых наблюдаются всхолмления со щебнистыми бугорковатыми вершинами и интенсивно заболоченные, часто заполненные водой понижения (фото 2, 3). Склоны сопок террасированы, многие террасы перекрыты суглинком и заболочены. Плоские вершины сопок и нагорные террасы несут черты криогенной сортировки обломочного материала и имеют нанорельеф в виде системы каменных колец, опоясывающих бугорки.
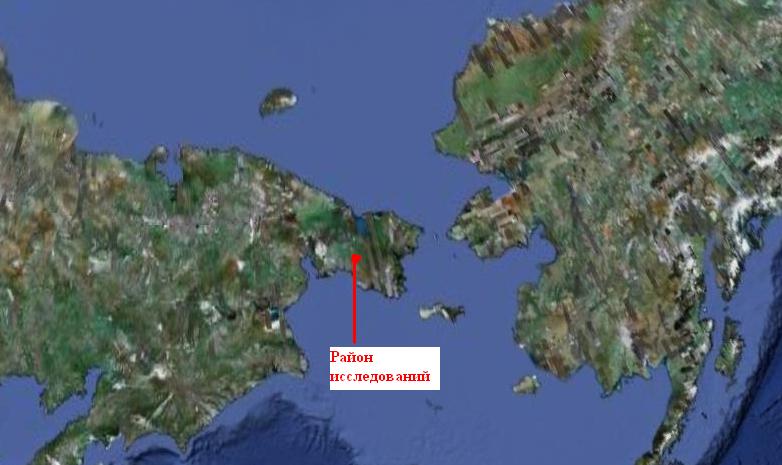
Фото 1. Положение района исследований (среднее течение р. Эргувеем) на космическом снимке.
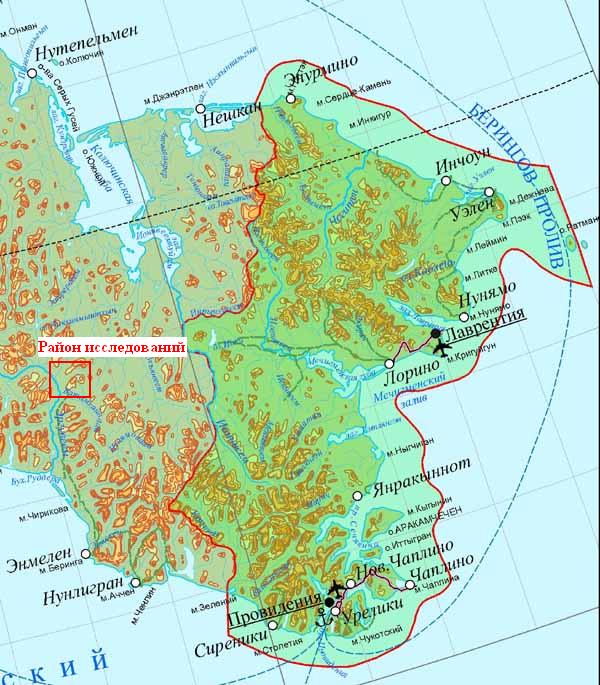
Карта 1. Положение района исследований на карте Чукотского полуострова. Зеленым цветом выделена территория современного парка "Берингия"
Склоны сопок резко переходят в пологие шлейфы и конуса выноса, где поверхностный слой суглинка содержит включения грубого обломочного материала. Шлейфы и конуса выноса имеют грядовый и бугорковатый криогенный микрорельеф. Криогенные процессы наблюдаются и на склонах сопок средней крутизны ( полосы, медальоны и т. д.), на крутых же склонах преобладают осыпи, в результате чего под такими склонами образуются россыпи незадернованного обломочного материала. Для района характерна густая речная сеть. В долинах более крупных рек (Эргувеем, Ватамкайваам, Пепенвеем) хорошо выражены пойма и первые надпойменные террасы, в пределах которых можно выделить следующие элементы: галечные и песчаныпляжи, береговые валы, заболоченные участки первой надпойменной террасы, берега старичных озер, заиленные берега временных проток.

Фото 2. Рельеф района низкогорный со следами былого оледенения. Фото А.А. Галанина.
|

Фото 3 . В то время в бассейне р. Эргувеем выпасались стада оленей, насчитывающие тысячи голов. Фото А.А. Галанина.
|
Климат района арктический, умеренно океанический (Клюкин, 1960).Средняя температура января приблизительно –25 , июля - +7 , причем зимой возможны оттепели, а летом — заморозки и выпадение снега. Осадков за год выпадает около 700 мм, на холодное время приходится 120 — 130 мм. Лето короткое, снег стаивает к середине июня и выпадает во второй половине сентября.
Благодаря расчлененному рельефу и сильным ветрам происходит перераспределение снега, и в депрессиях, оврагах, поймах рек и местах перегибов склонов образуются огромные снежники, которые едва успевают полностью стаять к концу лета. На многих мелких речках и ручьях зимой образуются наледи площадью в несколько десятков гектар.
Летняя облачность и большое количество жидких осадков способствуют переувлажнению и заболачиванию. Вода часто сочится по шлейфам и широким ложбинам стока. Район расположен в зоне сплошного распространения вечной мерзлоты (Клюкин, 1960). Мерзлота, особенно в условиях влажного климата, является основным фактором формирования микро- и нанорельефа и обусловливает высокую подвижность субстрата.
Флора района Эргувеем насчитывает 297 видов сосудистых растений, из них арктических 59, аркто-альпийских 156, гипоарктических и гипоаркто-монтанных 61, бореальных 19, аркто-бореальных 2. Долготный географический спектр флоры следующий: циркумполярных видов 104, широко распространенных в Евразии, не идущих в Америку 26, узко распространенных в Евразии (к востоку от р. Колымы), не идущих в Америку 16, евразиатско-западноамериканских 48, собственно восточносибирско- и чукотско- западноамериканских (в широком смысле, берингийских) 35, сибирско- и восточносибирско-американских 18, восточночукотско- американских 1, видов узкораспространенных, ареал которых занимает Чукотский полуостров, п-ов Сьюард, острова Берингова и Чукотского морей (узкоберингийских) 16. Приведенные цифры характеризуют флору как арктическую, берингийского склада.
Наиболее представительными по числу видов семействами являются: злаки (32 вида), осоковые (31), сложноцветковые (28), гвоздичные (23), ивовые (19), розовоцветные (18), камнеломковые (17), норичниковые (11), ситниковые (11), бобовые (9), верескоцветные (9).
Целью работы было выявление эколого-ценотических элементов конкретной флоры данного района. В связи с этим мы занимались разработкой и опробованием метода межвидовых сопряженностей, впервые примененного к растительному покрову целого ландшафтного района.
В понимании конкретной флоры мы следуем А. И. Толмачеву (1931, 1932). Согласно принятой методике, площадью выявления конкретной флоры следует считать территорию порядка 100 км2, в пределах которой с некоторой повторностью выявляется весь набор местообитаний (фаций), присущих данному ландшафту. В пределах этой территории на сходных местообитаниях обнаруживаются сходные наборы видов.
Эколого-ценотический элемент конкретной флоры объединяет виды со сходным распределением в растительном покрове данного района. Совокупность эколого-ценотических элементов отражает флористическую структуру растительного покрова на ландшафтном, или топологическом, уровне организации. На этом уровне распределение видов определяется эдафическими, микро- и мезоклиматическими факторами, в отличии от более высоких географических уровней, на которых ведущая роль принадлежит факторам макроклиматическим и историческим.
Характер эколого-ценотического распределения каждого вида может быть установлен путем определения сходства-различия между распределениями данного и остальных видов конкретной флоры. При использовании этого способа следует учитывать два важных момента: сходство-различие видов по общей встречаемости в растительном покрове и сходство-различие по совместной встречаемости.
Распределение видов в растительном покрове носит в значительной степени статистический, вероятностный характер. В связи с этим Л. Г. Раменский (1925) писал: «Глубину, ясность и определенность обстановки физического эксперимента приходится заменять сравнительной обработкой экстенсивного, поверхностного, но зато широко собранного материала массовых наблюдений». Кроме массовости материала, при статистическом исследовании объекта необходимо соблюдение принципа репрезентативности, согласно которому в выборке должны быть представлены все варианты генеральной совокупности с сохранением количественных отношений между ними, существующих в генеральной совокупности (Василевич, 1969). При описании растительности проблема репрезентативности встает особенно остро, в связи с тем, что выборка состоит не из дискретных единиц, а из более или менее однородных участков, «вырезаемых» из растительного покрова.
Случайный набор образцов, дающий наиболее репрезентативную выборку, к растительности большой территории практически неприменим. Это связано с тем, что за пределами исследованной территории соотношение площадей элементов ландшафта существенно меняется, и поэтому случайная выборка репрезентативна только относительно этой территории. Кроме того, заложение площадок методом случайных чисел на большой территории связано с огромными затратами времени и труда, которые, в силу вышеотмеченных обстоятельств, ни в какой мере не оправданы.
Нами был использован комбинированный способ: сочетание пропорционального отбора ключевых участков с систематическим отбором описаний в пределах этих участков. Три ключевых участка (фото 4) представляли собой квадраты по 9 кв. км каждый, расположение с учетом охвата всех элементов ландшафта в пределах района, площадь которого составляла приблизительно 150 кв. км. Расстояния между центрами конкретных описаний составляли 300 м. Описывались однородные на глаз участки растительного покрова, площадью не менее 20 кв. м. В этом случае, когда однородность прослеживалась на значительной площади, детально обследовался участок в 100 кв. м. Всего было составлено 450 геоботанических описаний.
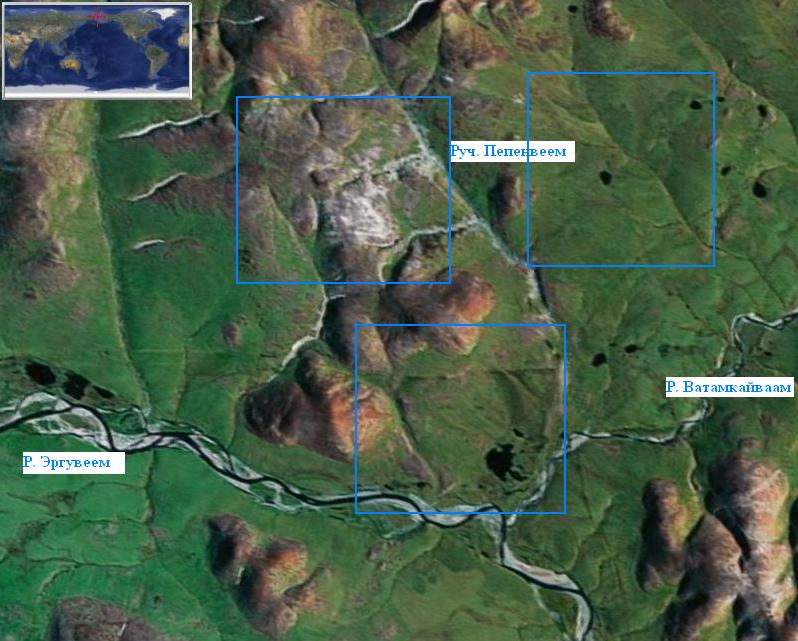
Фото 4 . Космический снимок района среднего течения р. Эргувеем. Показаны три квадрата, в пределах которых составлялись геоботанические описания.
При обработке использовались только данные встречаемости видов, показатели обилия не учитывались. В процессе обработки применялись перфокарты типа К-5. Путем прямого кодирования на перфокарты, каждая из которых несет информацию об одном виде, наносились данные о его встречаемости в описаниях, соответствующих номерам перфорации. Путем подсчета вырезов определялась общая встречаемость ( na ), равная отношению числа описаний с присутствием данного вида к общему числу описаний в выборе, а путем подсчета вырезов, совпадающих при наложении двух перфокарт, определялась совместная (попарная) встречаемость ( n a /в), равная отношению числа описаний, в которых встречаются оба вида вместе, к общему числу описаний в выборке.
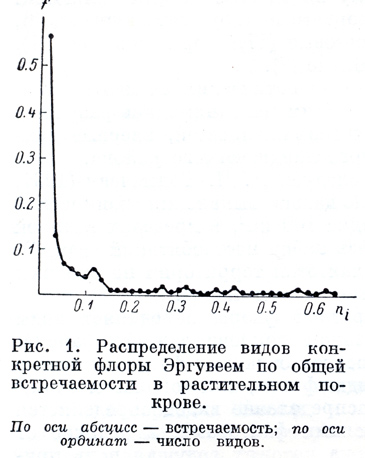 |
На основании полученных данных по общей встречаемости видов построена кривая распределения (рис.1.) Она показывает соотношение между числом видов и их встречаемостью в растительном покрове. Около 60% всех видов флоры встречаются менее чем в 2% описаний. Число же видов с высокой встречаемостью очень мало. Сильная растянутость кривой по оси X говорит о широком варьировании встречаемостей видов (до 64%). Благодаря наличию на кривой распределения деформаций деление видов на классы встречаемости становятся более естественным, так как границы между классами выбраны при тех значениях встречаемостей, на которые приходится меньшее число видов. При известной условности границ это позволяет сократить число граничных видов в классах до минимума. Всего выделено 5 классов по встречаемости: 1) 0.7 > n > 0.3; 2) 0.3 > n > 0.1; 3) 0.1> n > 0.5; 4) 0.05 > n > 0.015; 0.015 > n > 0.000.
Матрицы совместной встречаемости составлялись отдельно для каждого класса, за исключением пятого. Кроме того, совместная встречаемость определялась для видов соседних классов. Как нам представляется, подобный послойный метод анализа позволяет существенно уменьшить искажения, возникающие при расчете сопряженности между видами с разной встречаемостью, так как в этом случае в пределах одного класса диапазон изменения встречаемостей сравниваемых между собой видов по сравнению со всей флорой значительно снижен. Такой подход позволяет проанализировать всю флору целиком, не отбрасывая виды с высокой встречаемостью. Расчет же сопряженности между видами соседних классов позволяет увязать структуры, полученные в пределах отдельных классов в единую систему. Группы, выделенные таким путем, объединяют виды, сходные по двум параметрам — общей и совместной встречаемости.
Статистическая оценка распределения встречаемости двух видов основана на расчете степени отклонения их совместной встречаемости ( в случае независимости распределений) от действительно наблюдаемой совместной встречаемости. В качестве меры статистического сходства распределений видов в геоботанике используется несколько коэффициентов сопряженности (Cole, 1949; Грейг-Смит, 1967; Василевич, 1969, и др.), которые основаны на оценке распределения описаний по клеткам четырехпольной таблицы. |
В нашем случае абсолютная величина сопряженности не играет большой роли, и оценка эколого-ценотического сходства видов может быть произведена по трехбалльной шкале: 1) достоверная положительная сопряженность, 2) достоверная сопряженность отсутствует, 3) достоверная отрицательная сопряженность. При оценке достоверности сопряженности был использован метод Пирсона хи-квадрат (Василевич, 1969). Применение метода послойного анализа позволяет сократить число вычислений величин достоверности в пределах каждого класса встречаемости до нескольких, т. е. рассчитывать пороговое значение отклонения наблюдаемой встречаемости от (теоретической) ожидаемой, которое достоверно при данном объеме выборки. Определив наименьшее значение отклонения, достоверное для данного класса, мы отбираем пары видов (А и В), отклонение встречаемостей у которых выше порогового значения. Степень сопряженности расчитывалась по формуле:

Организация матриц производилась на основе построения графов. Результаты представлены на рис. 3, 4. Кружки с соответствующим номером обозначают виды. В зависимости от степени сопряженности они соединены линиями разной толщины.
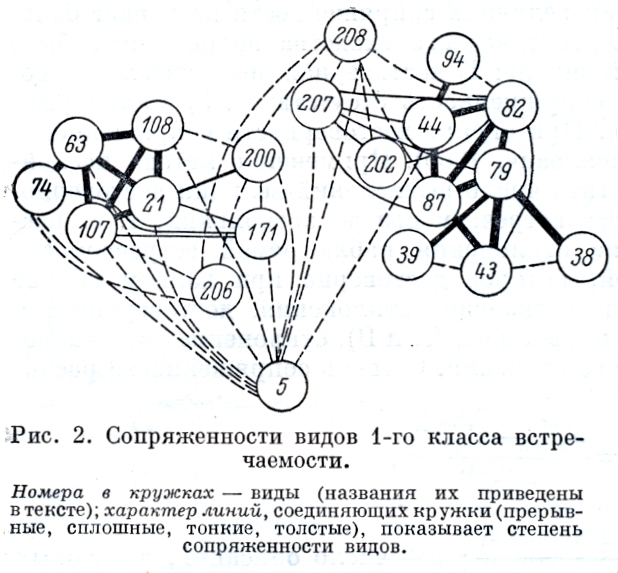 |
На рис. 2 представлены виды 1-го класса встречаемости. Среди них выделяются 2 группы положительно сопряженных видов и группа видов, слабо сопряженных как между собой, так и с видами первых двух групп. Первая группа состоит из Poa malacantha (21), Luzula confusa (63), Salix phlebophylla (74), Minuartia arctica (107), M. Macrocarpa (108), видов, которые имеют широкую экологическую амплитуду и распространены в щебнистых пятнистых и куртинных тундрах на склонах сопок и нагорных террас. Вторая группа более многочисленна, в нее входят Eriophorum vaginatum (38), E. angustifolium (39), Carex lugens (44), Salix fuscescens (79), Betula exilis (87), Salix pulchra (82), Carex stans (43), которые также имеют широкую экологическую амплитуду, но встречаются в осоково-пушицевых, пушицевых кочкарных и разнотравно-осоково-кустарничковых тундрах надпойменных террас, равнин, шлейфов и обширных нагорных террас. Группа слабосопряженных видов состоит из Hierochloe alpina (5), Diapensia obovata (206), Vaccinium vitis - idaea ssp . minus (208), которые имеют еще более широкую экологическую амплитуду. Hierochloe alpina встречается как в сухих щебнистых и щебнисто-суглинистых тундрах, так и в составе полигонально-валиковых болот на торфянистых микроповышениях. Diapensia obovata , встречаясь на сухих нагорных террасах и пологих склонах сопок, нередко заходит на сырые суглинисто-щебнистые участки шлейфов, изредка встречается на сухих торфяных буграх. Vaccinium vitis - idaea ssp. minus по широте экологической амплитуды не уступает Hierochloe alpina , но в отличие от нее на сильно обдуваемых участках не встречается. |
К группе положительно сопряженных видов горных местообитаний тесно примыкают Dryas punctata (171) и Cassiope tetragona (200).Дриада встречается в сухих щебнистых тундрах склонов и вершин сопок, нагорных террас, часто выходит на шлейфы, старые галечники надпойменных террас. Кассиопея приурочена к местам, где расположены снежники, но в данном районе встречается очень часто благодаря их широкому распространению. Это суглинистые щебнистые пятнистые тундры на шлейфах и внутренних частях обширных нагорных террас. Polygonum viviparum (94) и Vaccinium uliginosum ssp . microphylla (207) примыкают к группе видов равнинных местообитаний, но встречаются и в сухих щебнистых (голубика) и сырых щебнисто-суглинистых тундрах (горец) нагорных террас, шлейфов и пологих склонов.
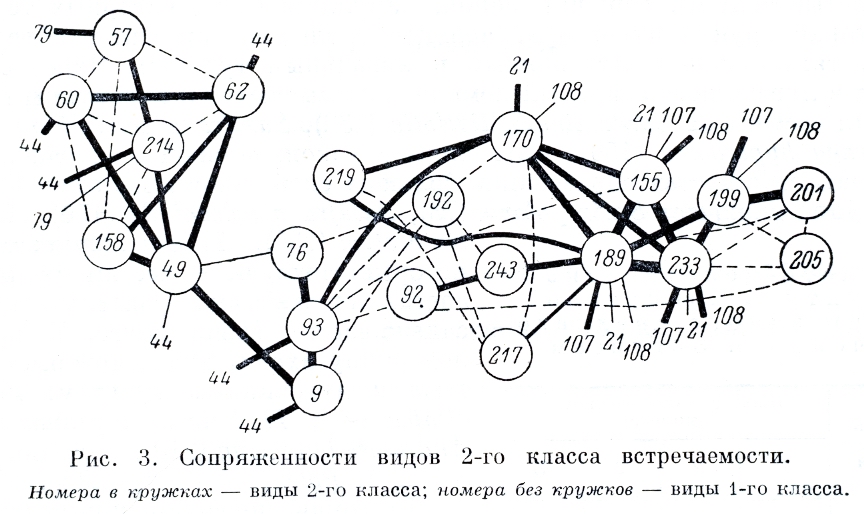 |
На рис. 3 изображены виды 2-го класса встречаемости. Степень дискретности образуемой ими структуры ниже, чем в 1-м классе, хотя закономерная ориентация видов выражена неплохо. В целом структура напоминает шнур с утолщениями на концах и перетяжкой в средней части. С одного конца группируются виды равнинных, с другого — горных местообитаний. В центре — непрочное скопление видов, характерных для сырых суглинистых и суглинисто-щебнистых тундр разных вариантов и оттенков. Анализ сопряженности между видами 1-го и 2-го классов позволяет равнинные виды обоих классов объединить в один комплекс. В подобный комплекс объединяются и виды горных местообитаний. Таким образом, во 2-м классе встречаемости происходит нарищивание групп видов, выявленных в 1-м классе. Кроме того, на этом уровне может быть выделено сочетание видов, промежуточных по |
своей экологии между краевыми группами. Это Carex misandra (49), Arctagrostis latifolia (9), Polygonum ellipticum (93), Salix reticulata (76), Novosieversia glacialis (170), Pedicularis willdenovii (219), Hedysarum obscurum s. l. (192), Polygonum tripterocarpum (92), Antennaria friesiana (243), Pedicularis capitata (217). Перечисленные виды не образуют замкнутой системы, что соответствует и характеру их экологии. У Carex misandra экологический оптимум — в сырых пятнистых суглинистых и суглинисто-щебнистых тундрах, однако она встречается и в осоково-пушициевых болотах вместе с C. rotundata и C. lugens, Acrtagrostis latifolia встречается в таких же группировках, но сильнее тяготеет к участкам с хорошим дренажем. Polygonum ellipticum часто входит в состав разнотравно-кустарничково-моховых шлейфов, конусов выноса и нагорных террас на хорошо увлажненных, но не заболоченных участках. Salix reticulata встречается в верхней части шлейфов склонов, конусов выноса, на внутренних и средних частях нагорных террас, в местах с интенсивным подтоком оснований. Novosieversia glacialis входит в состав более сухих щебнисто-суглинистых тундр нагорных террас и пологих склонов сопок. Pedicularis willdenovii встречается в сырых разнотравно-осочковых тундрах шлейфов и нагорных террас. Hedysarum obscurum обитает на сухих галечниках пойм, на надпойменных террасах, сухих участках шлейфов и нагорных террас, реже на пологих склонах солок. Polygonum tripterocaprum в данном районе служит неплохим индикатором участков с хорошим дренажем и селится на шлейфах склонов и нагорных террас. Antennaria friesiana является видом горным, но нередко заходит в пятнистые щебнисто-суглинистые тундры шлейфов и конусов выноса. Pedicularis capitata растет в сухих щебнистых тундрах шлейфов и нагорных террас.
Для сухих щебнистых тундр склонов сопок и нагорных террас характерны Oxytropis tschuktschorum и Artemisia furcata, которые изредка встречаются и в суглинисто-щебнистых тундрах с достаточным увлажнением. Arctous alpina и Empetrum nigrum приурочены к сухим щебнистым участкам нагорных террас и пологих склонов сопок в их нижней части, там, где скапливается достаточное количество снега, но где он стаивает не очень поздно и «нивальности» не наблюдается.
Виды равнинных местообитаний (Carex algida, Juncus castaneus, J. biglumis, Saxifraga foliolosa, Pedicularis sudetica ssp. albolabiata встречаются в сырых разнотравно-осоковых-моховых тундрах шлейфов, обширных нагорных террас, в местах с интенсивным минеральным подтоком, часто вдоль ручейков. Это весьма распространенные виды, которые нередко заходят и в обычные осоково-пушицевые тундры.
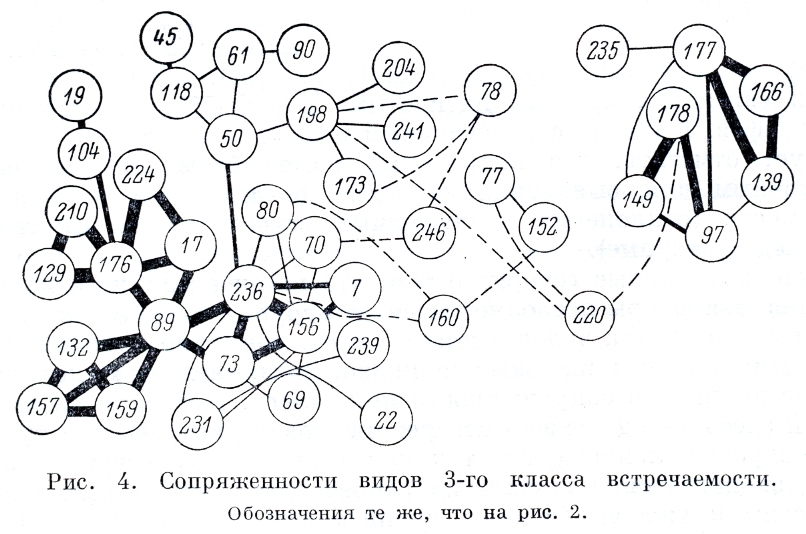 |
На рис. 4 изображены виды 3-го класса встречаемости. Здесь выделяются 2 группы. Первая группа Silene stenophylla (97), Draba stenopetala (139), Ermania parryoides (149), Saxifraga eschscholtzii (166), Potentilla uniflora (177), P. elegans (178). Это горные виды с относительно узкой экологической амплитудой, обитатели сухих щебнистых малоснежных тундр (крутые щебнистые склоны, вершины сопок, наружные части нагорных террас). Вторая группа имеет оригинальное и сложное строение, объединяя виды, тяготеющие к луговинным и нивальным тундрам. Через Oxyria digina в луговинно-нивальный комплекс связываются 3 подгруппы видов. Первая состоит из Cardamine bellidifolia (132), Saxifraga porsildiana (157), Saxifraga hyperborea (159) — это виды с узконивальной экологией. Они встречаются в местах образования больших снежников и наледей. Вторая подгруппа включает Salix polaris (73), Saxifraga nelsoniana (156), Artemisia arctica (236), Trisetum spicatum (7), которые тяготеют к местам скопления снега, но избегают участков, где он залеживается очень долго. В третью подгруппу входят виды, которые тяготеют к луговинным тундрам: Festuca altaica (17), Thalictrum alpinum (129), Androsace bungeana (210), Gentiana glauca (224), Potentilla emarginata (176). |
Ко второй подгруппе примыкает несколько видов: Luzula nivalis (69), Lloydia serotina (70), Salix chamissonis (80), Valeriana capitata (231), Nardosmia frigida (239), которые тяготеют к хорошо укрываемым снегом местам, но встречаются и в других тундрах.
В очень нечеткое образование связаны Carex atrofusca (50), Beckwithia chamissonis (118), Juncus albescens (61), Carex holostoma (45), Rumex arcticus (90), Rhododendron parvifolium (189), Loiseleuria procumbens (204), Saussurea angustifolium (241), Dryas octopetala (173), которые по характеру экологии связаны с сырыми пятнистыми суглинисто-щебнистыми тундрами шлейфов, в местах с интенсивным подтоком оснований.
Следует отметить, что полученный нами объем выборки оказался недостаточным для выявления сходства — различия видов 4-го класса встречаемости (отклонения наблюдаемых встречаемостей от теоретических — недостоверные).
Полученные данные говорят о том, что метод послойного корреляционного анализа флоры позволяет выделить группы сходных по характеру экологического ареала видов в классах с относительно высокой встречаемостью. Для анализа же видов с низкой встречаемостью получить достоверные данные по сопряженности не удалось. Несмотря на это, опробованный здесь метод может быть рекомендован для работы в тундровых и лесотундровых ландшафтах, так как редкие виды могут быть разделены на экологические группы на основе визуальных наблюдений или же отнесены к уже выделенным группам.
На основании построенных графов составлена развернутая схема эколого-ценотической структуры конкретной флоры района Эргувеем. Всего статистическим способом выделено четыре эколого-ценотических комплекса: 1) виды горных местообитаний — Г; 2) виды, занимающие промежуточное положение в корреляционной структуре между 1-м и 3-м комплексами, — Т; 3) виды равнинных местообитаний — Р; 4) виды нивально-луговинных местообитаний — Н. На основе визуальных наблюдений в 4-м и 5-м классах встречаемости могут быть выделены еще два комплекса видов пойменных местообитаний и прибрежно-водных видов. Комплексы Г, Р и Т имеют представителей во всех трех классах встречаемости, комплекс Н только в 3-м классе (см. таблицу).
Содержание клеток таблицы раскрыто в списках видов эколого-ценотических групп и комплексов. В этих списках кратко охарактеризованы особенности долготного и широтного географического распространения видов.
ВИДОВОЙ СОСТАВ ЭКОЛОГО-ЦЕНОТИЧЕСКИХ ГРУПП И КОМПЛЕКСОВ КОНКРЕТНОЙ ФЛОРЫ ЭРГУВЕЕМ.
Г. ГОРНЫЕ ВИДЫ
1-й класс встречаемости.
• Poa malacantha Kom., чукотско-западноамериканский аркто-альпийский.
• Luzula confusa Lindl., циркумполярный аркто-альпийский.
• Salix phlebophylla Anderss., чукотско-западноамериканский аркто-альпийский.
• Minuartia arctica (Stev. ex Ser.) Aschers. et Graebn., сибирско-западноамериканский аркто-альпийский.
• M . macrocarpa (Pursch) Ostenf., сибирско-западноамериканский арктоальпийский.
• Dryas punctata Jus., евразиатско-западноамериканский аркто-альпийский.
2-й класс встречаемости.
• Oxytropis tschuktschorum Jurtz., охотско-чукотско-западноамериканский и аркто-альпийский.
• Antennaria friesiana (Trautv.) Ekman, собственно восточносибирско-американский арктический.
• Novosieversia glacialis ( Adam) F. Bolle, сибирско-западноамериканский и аркто-альпийский.
• Saxifraga firma Litw. ex A. Los., собственно восточносибирско-западноамериканский аркто-альпийский.
• Artemisia furcata M. B., восточносибирский аркто-альпийский.
• Rhododendron kamtschaticum Pall., охотско-восточночукотско-западноамериканский гипоаркто-монтанный.
• Arctous alpina (L.) Nied., циркумполярный гипоаркто-монтанный.
• Empetrum nigrum L. s. l., циркумполярный гипоаркто-монтанный.
3-й класс встречаемости.
• Silene stenophylla Ledeb., собственно восточносибирский аркто-альпийский.
• Draba stenopetala Trautv., охотско-восточночукотско-западноамериканский аркто-альпийский.
• Ermania parryoides Cham. ex Botsch., собственно восточносибирско-западноамериканский аркто-альпийский.
• Saxifraga eschscholzii Sternb., чукотско-западноамериканский аркто-альпийский.
• Potentilla uniflora Ledeb., восточносибирско-западноамериканский аркто-альпийский.
• P . elegans Cham. et Schlecht., собственно восточносибирско-западноамериканский аркто-альпийский.
• Artemisia borealis Pall., циркумполярный гипоаркто-монтанный.
Р. РАВНИННЫЕ ВИДЫ.
1-й класс встречаемости.
• Eriophorum vaginatum L., собственно восточносибирско-американский арктический вид.
• E . angustifolium Honck., циркумполярный аркто-бореальный.
• Carex stans Drej., циркумполярный арктический.
• C . lugens H. T. Holm., собственно восточносибирско-американский гипоарктический.
• Salix fuscescens Anderss., восточносибирско-западноамериканский гипоарктический.
• Betula exilis Sukacz., восточносибирско-западноамериканский гипоарктический.
• Salix pulchra Cham., сибирско-западноамериканский гипоарктический.
• Polygonum viviparum L., циркумполярный аркто-альпийский.
• Ledum decumbens (Ait.) Small, сибирско-американский гипоаркто-монтанный.
2-й класс встречаемости.
• Carex algida Turcz. ex V. Krecz., сибирско-американский гипо-арктический.
• C . misandra R. Br., циркумполярный аркто-альпийский.
• Juncus castaneus Smith, циркумполярный аркто-альпийский.
• J. Biglumis L., циркумполярный аркто-альпийский.
• Saxifraga foliolosa R. Br., циркумполярный аркто-альпийский.
• Pedicularis sudetica Willd. ssp. albolabiata Hult., восточносибирско-американский аркто-альпийский.
3-й класс встречаемости.
• Carex holostoma Drej., почти циркумполярный аркто-альпийский.
• C. atrofusca Schkuhr, циркумполярный спорадический аркто-альпийский.
• Juncus albescens (Lange) Fernald, восточносибирско-американский аркто-альпийский.
Т. ВИДЫ, ЗАНИМАЮЩИЕ ПРОМЕЖУТОЧНОЕ ПОЛОЖЕНИЕ
В КОРРЕЛЯЦИОННОЙ СИСТЕМЕ.
1-й класс встречаемости.
• Cassiope tetragona (L.) D. Don, циркумполярный аркто-альпийский.
• Diapensia obovata (F. Schmidt ) Nakai, восточносибирско-западноамериканский аркто-альпийский.
• Hierochloe alpina ( Liljebl.) Roem. et Schult., циркумполярный аркто-альпийский.
• Vaccinium vitis-idaea L. ssp. Minus (Lodd.) Hult., циркумполярный гипоарктический .
2-й класс встречаемости.
• Arctagrostis latifolia (R. Br.) Griseb., циркумполярный аркто-альпийский.
• Polygonum bistorta L. ssp. ellipticum (Willd.) Petrovsky, собственно восточносибирско-западноамериканский аркто-альпийский.
• Salix reticulata L., почти циркумполярный аркто-альпийский.
• Pedicularis wildenovii Vved., евразиатско-западноамериканский аркто-альпийский.
• Polygonum tripterocarpum A. Gray, ex Rothrock , восточносибирский гипоарктический.
• Hedysarum obscurum L. s. l., евразиатско-западноамериканский аркто-альпийский.
• Pedicularis capitata Adams., восточносибирско-американский аркто-альпийский.
3-й класс встречаемости.
• Rohdiola atropurpurea (Turcz.) Trautv. Et Mey., охотско-чукотско-западноамериканский аркто-альпийский.
• Salix chamissonis Anderss., охотско-восточночукотско-западноамериканский аркто-альпийский.
• Lloydia serotina (L.) Reichenb., сибирско-западноамериканский аркто-альпийский.
• Nardosmia frigida (L.) Hook., евразиатско-западноамериканский гипоарктический.
• Luzula nivalis Laest., циркумполярный аркто-альпийский.
• Beckwithia chamissonis (Schlecht.) Toim., чукотско- западноамериканский арктический.
• Rhododendron parvifolium Adams, собственно восточносибирской гипоаркто-монтанный.
• Rumex arcticus Trautv., сибирско-западноамериканский арктический.
• Loiseleuria procumbens (L.) Desv., европейско-американско-собственно-восточносибирский аркто-альпийский.
Saussurea angustifolia (Willd.) DC., чукотско-американский гипо-арктический.
11. Dryas octopetala L., евразиатско-западноамериканский аркто-альпийский.
12. Salix sphenophylla A. Skv., восточносибирско-западноамериканский аркто-альпийский
13. Saxifragagrandipetala (Engler et Irmsch.) A. Los., чукотско-западноамериканский арктический.
Н. НИВАЛЬНЫЕ ВИДЫ 3-ГО КЛАССА ВСТРЕЧАЕМОСТИ.
1. Oxyria digyna (L.) Hill, циркумполярный аркто-альпийский.
2. Salix polaris Wahlenb., евразиатско-западноамериканский аркто-альпийский.
• Cardamine bellidifolia L., циркумполярный аркто-альпийский.
• Saxifraga porsildiana (Calder et Savile) Jurts. et Petrovsky, чукотско-западноамериканский аркто-альпийский.
• Saxifraga hyperborea R. Br., циркумполярный аркто-альпийский.
• Festuca altaica Trin., восточносибирско-западноамериканский гипоаркто-монтанный.
7. Potentilla emarginata Pursch (non Desf.), циркумполярный аркто-альпийский.
8. Artemisia arctica Less., собственно восточносибирско-западноамериканский аркто-альпийский.
9. Thalictrum alpinum L., почти циркумполярный аркто-альпийский.
• Androsace bungeana Schischk. et Bobr., сибирско-западноамериканский аркто-альпийский.
• Stellaria ciliatosepala Trautv., циркумполярный аркто-альпийский.
• Gentiana glauca Pall., собственно восточносибирско-западноамериканский аркто-альпийский.
• Saxifraga nelsoniana D. Don, сибирско-западноамериканский аркто-альпийский.
• Trisetum specatum (L.) Richt., циркумполярный аркто-альпийский.
• Valeriana capitata Pall., евразиатско- западноамериканский аркто-альпийский.
• Poa paucispicula Scribn. Et Merr., восточносибирско - западноамериканский аркто-альпийский.
Анализ и сравнение этих списков показывает, что связь между экологическим и ареалогическим сходством видов очень незначительная. В одну эколого-ценотическую группу или комплекс могут входить виды с весьма разными ареалами, от узкоберингийских до циркумполярных. Однако среди равнинных видов процент циркумполярных и почти циркумполярных выше, чем среди горных. В порядке увеличения процента видов с широким географическим распространением в долготном направлении эколого-ценотические комплексы можно расположить в следующем порядке: Г— Т— Н— Р.
Интересно, что виды, не заходящие в Америку, имеются только в комплексах Г и Т. Это позволяет говорить о том, что равнинные пространства осушенного шельфа в известной степени являлись барьером для распространения горных видов, равнинные же виды мигрировали более свободно (Юрцев, 1973).
В отношении широтного распространения видов наиболее однородными являются комплексы Н и Г, среднее положение занимает комплекс Т и наиболее однороден комплекс Р. На наш взгляд, это связано с тем, что экологический режим в горных местообитаниях сильнее связан с общеклиматическим режимом района по сравнению с равнинными местообитаниями. Именно благодаря некоторой «независимости» от изменений общего климата в сообществах равнин совместно произрастают арктические, гипоарктические, гипоаркто-монтанные и аркто-альпийские виды. Растительный же покров горных местообитаний более чувтвителен к общеклиматическим изменениям, и изменения растительности здесь происходят раньше, чем на равнинах.
Разделение видов на классы в зависимости от их общей встречаемости соответствует задачам флористических исследований, так как при неблагоприятных изменениях климата виды не меняют общего характера своей экологии, а снижают активность в смысле Б. А. Юрцева (1968); поэтому они не исчезают из флоры, а в незначительных количествах сохраняются в отдельных местообитаниях, иногда весьма различных. Таким образом, в первоначальных условиях относясь к одной экологической группе, в новых условиях эти виды не образуют сопряженных систем. Снижение активности видов в ландшафте часто приводит к спорадичности их распространения в пределах данного района. Однако спорадичность может быть вызвана и спорадичностью распространения отдельных эдафических факторов, с которыми связаны некоторые виды флоры. Так, спорадичными могут быть растения-кальцефиты в районе, где богатые кальцием субстраты встречаются спорадически. В районе, Эргувеем к таким видам можно отнести Potentilla bifora Willd., Artemisia globularia Bess., Kobresia simpliciuscula ( Wahl.) Maak. Спорадичность же таких видов, как Festuca altacia Trin., Spiraea stevenii Rydb., Dodecatheon frigidum Cham. et Schlecht., Carex rupestris Bell. ex All., Sanguisorba officinalis L. s. L., с какими-либо определенными факторами среды не связана и вызвана снижением активности этих некогда широко распространенных видов. Вопрос о причинах спорадичности распространения видов заслуживает самого пристального внимания флористов и геоботаников.
ЛИТЕРАТУРА
Арктическая флора СССР. (1962— 1971). 1—6.—Василевич В. И. (1969). Статистические методы в геоботанике. — (Грейг—Смит П.1967). Количественная экология растений. — Клюкин Н. К. (1960). Прикладной климатологический справочник Северо-Востока СССР. — Раменский Л. Г. (1925). Основные закономерности растительного покрова и их изучение. Вестн. опытн. дела.— Толмачев А. И. (1931). К методике сравнительно-флористических исследований Бот. ж., 16,1.— Толмачев А. И. (1932). Флора центральной части Восточного Таймыра. Тр. Полярн. комисс., 8, ч. 1.— Юрцев Б. А. (1968). Флора Сунтар-Хаята. Проблемы истории высокогорных ландшафтов.— Юрцев Б. А. (1973). Ботанико-географическая зональность и флористическое районирование Чукотской тундры. Бот . ж ., 58,7.— Cole L. (1949). The measurement of interspecific association. Ecologu, 30.
Калининградский государственный Университет (Получено 3 V 1973).
ПОСЛЕСЛОВИЕ
Прошло 40 лет, как я написал эту статью и 37 лет, как она вышла в Ботаническом журнале. Для жизни человека это много. Для истории - один миг. Для науки?... Смотря что считать наукой. В той науке, которой занимаюсь я (ботаническая география, флористика), принципиально нового ничего не появилось, хотя лавина публикаций мощная. Но никаких прорывов в теории и методологии нет. Сегодня практически в каждой флористической и ботанико-географической статье, а тем более монографии, используется понятие "эколого-ценотический элемент флоры", никто уже и не ссылается на авторов этого понятия. Это же очевидно, что флора состоит из эколого-ценотических элементов, что она имеет эколого-ценотическую структуру. Если и ссылаются, то на работы иностранных авторов последнего десятилетия. А ведь они были не первыми. Сегодня считается шиком процитировать иностранного автора, выказав тем самым свое преклонение перед всем иностранным и пренебрежение всем отечественным. За такой подход в институтах дают надбавки к жалованию, при конкурсе заявок на гранты это тоже учитывается в первую очередь. А ведь это неправильно. По большому счету, это разрушает нашу страну, нашу национальную культуру. А может, я не прав?

