
|
Дифференциация деревьев разных видов по мощности по мере их роста и в зависимости от влияния друг на друга
А.В. Галанин © 2013
С этой проблемой сталкиваются все лесоведы и лесоводы. Пришлось с ней столкнуться и мне. Две мои аспирантки изучали дифференциацию деревьев в естественных популяциях в лесах заповедников Дальнего Востока и Забайкалья. Для этого они закладывали постоянные пробные площади и измеряли на них толщину всех деревьев с периодичностью раз в 5 лет. Оказалось, что все деревья одного вида на пробной площади по мере их роста четко разделяются (дифференцируются) на несколько дискретных групп по толщине ствола.
|
Вот эти группы в результате статистической обработки измерений и установили мои аспирантки. Кроме того, они установили, что дискретные группы деревьев сохраняются по прошествии нескольких лет, они весьма устойчивы, хотя отдельные особи по мере роста могут как прибавлять в скорости роста, так и замедлять ее. Особи по мере роста могут переходить из одной группы толщины в другую, но при этом размерная структура остается дискретной – группы не только не исчезают (не сливаются друг с другом), но напротив, их дискретность может увеличиваться.
Сделав вывод о существовании механизмов самоорганизации в естественных древостоях, в результате которых в сообществах деревьев увеличивается разнообразие функциональных элементов и оптимизируется биогеохимическая работа экосистемы, аспирантки вышли на защиту своих диссертаций. Защиты прошли успешно, и ряды геоботаников пополнились двумя кандидатами биологических наук. Но при защите у одного из оппонентов была претензия к главному выводу. Суть вывода – в том, что причиной дифференциации являются конкурентные отношения особей друг с другом, в результате которых деревья как бы "удаляются" друг от друга на расстояния, на которых угнетают друг друга в меньшей степени. Разумеется, особи не бродят по площади, но те из них, которые оказываются на "запрещенных" конкуренцией расстояниях от особей лидеров, просто погибают, в результате чего древостой изреживается. Оппонент заявил, что дискретные группы особей образуются из-за того, что в них входят особи разного возраста, а появление всходов под пологом леса происходит неравномерно, а порциями – в некоторые годы активно, а в другие нет.
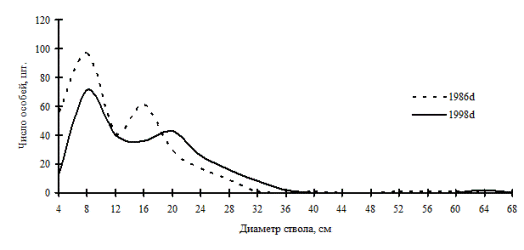
Частотные распределения особей дуба монгольского в дубняке лещиновом папоротниково-разнотравном на территории Сихотэ-Алинского биосферного заповедника. По оси ординат – диаметр стволов в см, по оси абсцисс – число особей на пп. (По Н.А. Василенко, 2008). Диаграмма с сайта: ttp://geobotany.narod.ru/vasilenko_2.htm |
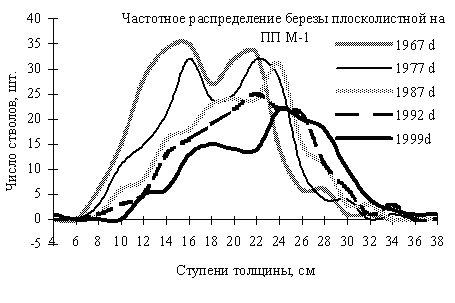
Частотные распределения особей березы плосколистной в разные годы, по 2-х сантиметровым ступеням толщины в Сихотэ-Алинском заповеднике. По оси ординат – диаметр стволов в см, по оси абсцисс – число особей на пп. (По Н.А. Василенко, 2008). Диаграмма с сайта: http://geobotany.narod.ru/nachod2.htm |
Итак, интрига здесь в том, является ли дифференциация деревьев по толщине следствием их разного возраста, или роль возраста как фактора дифференциации деревьев по размеру в лесу – не главная. К сожалению, измерить возраст каждого дерева на пробных площадях в заповедниках мы не могли в силу ряда причин. Определение же возраста у 10% особей наших выборок и выявленная слабая корреляция возраста и диаметра стволов на пробных площадях оппонента не убедили.
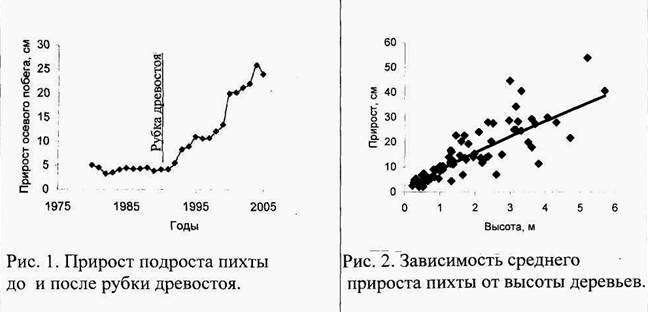
Графики с сайта: http://science-bsea.bgita.ru/2… |

Фото с сайта: http://www.newsufa.ru/Image/cache/Image_ _getty_rf_photo_of_tree_rings_300xauto.jpg |
В геоботанической и лесоводственной научной литературе опубликовано немало фактов, доказывающих правоту моих аспиранток. Снижение конкурентного пресса в лесу после рубок ухода вызывает резкое увеличение прироста осевого побега у оставшихся деревьев, что показано на рис. 1. Косвенно этот график доказывает то, что возраст деревьев в лесу – не главный фактор дифференциации по размерам и эдафической мощности. На рис. 2 видно, что у более высоких особей пихты прирост значительно больше. Чем выше в ценопопуляции становятся особи лидеры, те меньше шансов у отставших от них в росте соседей догнать лидеров. На фото поперечного разреза ствола ели (вверху справа) отчетливо видно, как интенсивно особь прирастала в первые 9–10 лет своей жизни, когда конкуренция между елочками в посадке еще отсутствовала, так как их фитогенные поля еще не смыкались друг с другом, и елочки еще не конкурировали за ресурсы внешней среды. Однако затем, когда елочки достигли такой величины, что их фитогенные поля сомкнулись, началась конкуренция за ресурсы, и прирост в толщину резко снизился.
Приведенные факты однозначно доказывают, что в ценопопуляциях деревьев по мере их роста усиливается конкуренция за ресурсы и происходит дифференциация по размеру и ценотической мощности деревьев. В этом случае возраст особи слабо связан с ее высотой, толщиной и ценотической мощностью.

|
В 2004 г. в своем саду возле дома во Владивостоке я посадил несколько особей разных видов хвойных деревьев: лиственницу даурскую (Larix dahurica), пихту белокорую (Abies nephrolepis), тис остроконечный (Taxus cuspidata) и пихту цельнолистную (Abies heterophylla). Все саженцы были выращены из семян и в момент посадки были примерно одного размера и одного возраста – 4–5 лет. На фотографии слева видно, как малы были эти деревца. Тогда я надеялся, что к моему выходу на пенсию деревца подрастут, и в углу сада будет прекрасный фрагмент хвойного леса.
Итак, стартовые условия у всех особей были примерно равные. Но уже в первый год (2005) прирост в высоту у особей пихты цельнолистной был больше, чем у особей пихты белокорой. Оказалось, что генотип пихты белокорой меньше соответствует климату Южного Приморья, чем генотип пихты цельнолистной. Хотя уже в Среднем Сихотэ-Алине генотип пихты белокорой больше подходит климатическим условиям, пихта цельнолистная там отсутствует. А пихта белокорая отсутствует в Южном Приморье, хотя на значительной высоте на склонах вулкана Чанбайшань в Корее и в Китае пихта белокорая вместе с елью корейской образует верхний лесной пояс. |

|
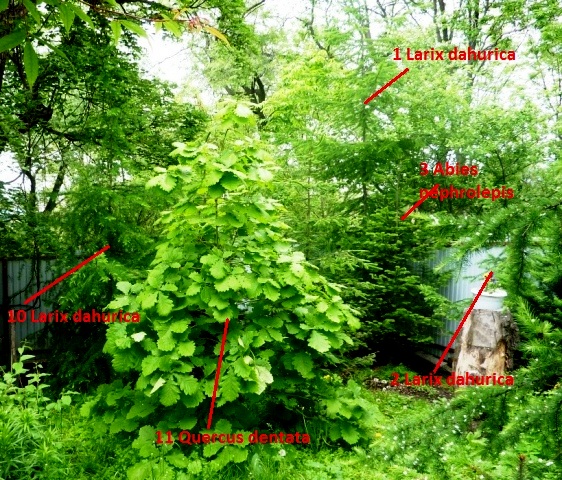
В 2007 г. деревца заметно подросли, особенно особи лиственницы под номерами 1 и 10, а вот лиственницы под номерами 2 и 3 заметно отстали от них в росте. Особи 2 и 3 были посажены рядом с двумя особями тиса остроконечного, высаженными одновременно с лиственницами и пихтами.
Таким образом, подтвердилось сильное фитонцидное угнетение тисом остроконечным лиственницы даурской. Эти виды – из разных историко-генетических комплексов и совершенно не приспособлены к жизни в одном сообществе. Тис когда-то в третичном периоде образовывал древостои и доминировал в лесах юга Дальнего Востока. Но с тех пор климат здесь сильно изменился и тис превратился в изгоя, сохранившись как реликт кое-где под пологом лесов из кедра корейского. Его фитонцидные свойства, даже ядовитость коры и хвои для соседей, способствуют тому, что вид этот пережил многих своих третичных видов "коллег".
В 2006 г. в компанию к пихтам цельнолистным я посадил крошечную особь дуба зубчатого Quercus dentata, которую привез из Хасанского района из окрестностей п. Зарубино. В окрестностях Владивостока и севернее этот вид не встречается. |

|
В 2010 г. различия в росте лиственницы номер 1 и номер 2 стали просто разительными. Тис, который из-за пня ильма не виден, резко затормозил рост лиственницы номер 2.
Пихты жестколистные и пихты белокорые пока не конкурируют между собой и растут быстро, но значительно медленнее, чем лиственница номер 1.
Прекрасно растет и дуб зубчатый. Надо сказать, что дуб зубчатый, в отличие от дуба монгольского, – вечнозеленое растение, не имеющее генетически обусловленной способности осенью сбрасывать листья. Они у него просто погибают от мороза, но мертвые остаются на веточках до весны и опадают в конце весны – начале следующего лета. Дуб зубчатый образует древостои на самом юге Приморского края и в Корее. Он прекрасно приспособлен к перенесению лесных и степных пожаров. |
|
В 2012 г. различия между особями лиственницы номер 1 и номер 2 стали разительными. Не зная историю их посадки, трудно поверить, что они одного возраста и одного вида. Лиственница номер 10 того же возраста, что и лиственницы номер 1 и номер 2, и посажена она была одновременно с ними. Эта лиственница немного уступает по размерам лиственнице номер 1, хотя рядом с ней растет мощный конкурент за свет, воду и минеральные элементы - мощная старая особь липы маньчжурской. Но негативное влияние тиса на лиственницу номер 2 значительно сильнее влияния липы на лиственницу номер 10.
Дуб зубчатый заметно вырос и успешно конкурирует как со старой липой, так и с молодыми пихтами цельнолистными. |

|
Так выглядит уголок нашего сада в 2013 г. Когда-то совсем недавно в 2002 г. здесь был замусоренный в течение десятилетий участок – помойка.
Лиственница даурская справа на переднем плане – тоже ровесница всех других лиственниц, посаженных мной в 2004 г. Посмотрите, как сильно она отличается от своих сестер, что посажены рядом с тисом остроконечным. А ведь они ровесницы и посажены здесь одновременно. |

|
В 2013 г. дуб зубчатый радовал нас своими огромными кожистыми листьями, развивающимся крепким стволом и большими приростами верхушки и боковых ветвей.
Тис остроконечный совсем забил лиственницы с номерами 2 и 3, а лиственницы с номерам 1 и 10 в росте опередили всех своих ровестников соседей. |

|
В 2013 г. особь лиственницы номер 1 достигла в высоту 7 м, в то время как высота особи лиственницы номер 2 едва превышает 2 м. У нее изогнутые веточки, мелкая хвоя, и верхушка по мере роста отклоняется от тиса. Пихта белокорая номер 3 отстала в росте от пихты цельнолистной номер 7.
Высота дуба зубчатого превысила 3 м. Он прекрасно растет, захватывая все большее пространство и становится активным конкурентом особей пихты цельнолистной.
Уголок хвойно-широколиственного леса в нашем саду сформировался, в нем прекрасно пахнет хвоей, в него слетаются птицы во время весеннего и осеннего пролетов, а некоторые вьют гнезда на липе и ильмах и выводят птенцов. А по утрам радуют нас своими песнями.
Старый ильм, который спилили прежние хозяева, мы заменили новыми молодыми деревьями сразу 5 видов.
В 2013 г. в этом лесном уголке любят прятаться соседские коты и кошки, а наша собака лайка ревниво прогоняет их. Но нашего кота, напротив, лайка защищает от чужих котов, что коту, кстати, не всегда нравится. Но вообще наш кот очень хорошо относится к своей собаке, иногда играет с ней в догонялки. А вот чужих собак боится и забирается от них либо на карниз дома под крышу, либо на липу, либо на ильмы. |

|
За 8 лет на пустыре в результате посадки очень маленьких саженцев нескольких видов деревьев сформировался оригинальный молодой лес, точнее, фрагмент леса. За этот короткий срок произошла дифференциация деревцов, так как они вступили друг с другом в конкурентные отношения за свет, воду и элементы минерального питания.
На этом примере можно убедиться, что возраст дерева и его мощность связаны весьма слабо, поэтому определять возраст деревьев по толщине ствола на пробных площадях очень опасно: можно прийти к совершенно неверным выводам.
Знать конкурентные и аллелопатические взаимодействия разных видов деревьев при создании посадок просто необходимо.
В условиях города Владивостока вблизи автодорог (на расстоянии 10 м) можно выращивать все виды деревьев, использованные в настоящем эксперименте. Следует помнить, что тис остроконечный обладает очень сильным фитонцидным влиянием на лиственницу даурскую. Дуб зубчатый вполне может активно использоваться в городских посадках в г. Владивостоке.
Считаю, что проведенный мной эксперимент с посадкой саженцев деревьев на пустыре в центре города позволяет сделать такие выводы. |
|

