УДК 634.0.18
Эколого-ценотическая структура и динамика прирусловых кедровых
и кедрово-лиственичных лесов в верхнем лесном поясе
Сохондинского биосферного заповедника
А.В. Галанин, Л.М. Долгалева
 |
© OCR А.В. Галанин, Л.М. Долгалева. Публикуется по: А.В. Галанин, Л.М. Долгалева. Эколого-ценотическая структура и динамика прирусловых кедровых
и кедрово-лиственичных лесов в верхнем лесном поясе
Сохондинского биосферного заповедника // Экосистемы Сибири и Дальнего Востока. Находка: ИТИБ, БСИ ДВО РАН, 2010. С. 21-44.
Флора и растительность окрестностей горы Сохондо (ныне она принадлежит территории Сохондинского заповедника) исследовались на протяжении более 200-летнего периода со времен первых флористических находок Н.П. Соколова (1793), работавшего в составе экспедиции П.С. Палласа (1772 и 1773). Позднее Г.И. Радде изучал районы Сарды и хребта Хамар-Дабан (территория современной Бурятии). Исследования Г.И. Радде, осуществленные в рамках экспедиции, были изданы на немецком языке - «Reisen im Suden von Ost-Sibirien in den Jahren 1855-1859». Работы Черкасова П.А. (1867, 1884) также расширили флористический список Даурии и пополнили знания о разнообразии экосистем. Список работ современных исследователей включает в большей степени флористические списки и очерки растительности заповедника (Дылис и др, 1965; Виппер, 1968; Сишшвинский, 1968; Худяков, 1976; Коротков, 1978; Дулепова, 1979, 1983; Дулепова, Уманская, 1983; Касич, 1983; Васильченко, 1983, 1984, 1987, 1988; Васильченко, Галанин, 1983; Галанин, Проскурина, 1983; Васильченко, Шевырева, 1988; Дулепова, Уманская, 2000; Галанин, Беликович, 2002а; Беликович, Галанин, 2005; и др.). Постоянные мониторинговые наблюдения, начало которым положено в Сохондинском заповеднике (СГБЗ) в 80-х гг. прошлого столетия, сделали возможным изучение структуры и ее динамики на основе многолетних наблюдений, определение изменения в строении ценозов, анализ состояния лесной растительности на ключевых участках (Галанин, 1989, 1991; Галанин, Беликович, 1988, 20026, 2002в; Беликович, Галанин, 2000, Долгалева, 2009 а, б). |
Введение
Решение проблем мониторинга и сохранения биоразнообразия лесной растительности должно основываться на теоретических представлениях о ее составе и структуре. Последние, в свою очередь, подвержены сильнейшим естественным и антропогенным флуктуациям, которые должны оцениваться по натурным описаниям наиболее динамичных лесных сообществ. Наиболее чувствительны к изменению экологических факторов леса, испытывающие влияние флювиальных процессов. В бореальных (таежных) лесах в таком качестве обычно рассматривают как пойменные, прирусловые, приручьевые леса (Лесная энциклопедия, 1986). Однако анализ литературы, посвященной лесам, связанным с флювиальными процессами Западной Сибири (Таран, 1990, 1993, 1995, 1998, 2000), показали, что, по сути, исследования ограничиваются пойменными сообществами и не затрагивают хвойные леса рек высоких порядков, видовое и структурное разнообразие которых существенно различно. Различными могут быть и сукцессионные процессы, происходящие на верхних отметках размещения лесов в горах.
В условиях ксерофитизации климата прирусловая и пойменная растительность оказывается весьма чувствительной к происходящим изменениям, она часто первая реагирует на климатические и антропогенные флуктуации, что позволяет считать эти экосистемы удобными объектами для изучения динамики эколого-ценотической структуры. В связи с этим в работе были поставлены задачи: описать и типизировать сообщества прирусловых хвойных лесов верхнего лесного пояса модельных участков Сохондинского заповедника; оценить их видовое и структурное разнообразие; определить их динамику в ходе экологических смен и составить предварительные прогнозы изменения структуры в ходе сукцессии.
Исследования в Сохондинском государственном биосферном заповеднике планировались как длительный мониторинг растительного покрова. Для этого в заповеднике заложена система постоянных пробных площадей (Галанин, Беликович, 2004). С периодом в 5-7 лет проведено переописание пробных площадей по стандартной методике с перечетом древостоя, оценкой обилия видов, картированием растительности. Работа выполнялась в рамках программы ДВО РАН «Комплексные исследования в бассейне р. Амур на период 2004-2007 гг.». Фактический материал включает 125 геоботанических описаний, данных 10 постоянных пробных площадей (ППП) прирусловых лесов, заложенных по стандартной методике.
Заповедник площадью 210988 га в соответствии с новым геоботаническим районированием, предложенным группой авторов (Галанин и др., 2008) расположен в Ононской и Хентейской провинциях в пределах Даурской ботанико-географической подобласти Восточно-Азиатской области. Принадлежность в большей степени Амурскому бассейну (реки Букукун, Агуца, Агуцакан, Кыра, Ингода), и в меньшей - Байкальскому (р. Буреча) определила ряд особенностей растительности, отчасти идентичной с Восточной и Южной Сибирью, а также имеющей сходные черты с Маньчжурией и Сихотэ-Алинем. Климатические циклы Центральной Азии также усиливали степные черты растительности в аридные эпохи, способствовали ускорению сукцсссионных смен, влияли на границы высотных поясов. Размещение на юге Восточной Сибири ООПТ с огромным горным массивом на стыке степной и лесной области, геоморфологические и климатические особенности, наличие высокогорных хвойных лесов на южной границе провинции уникальны для мониторинга лесных экосистем.
Структура фитоценозов трактуется некоторыми авторами как в широком аспекте (Мазинг, 1969), так и в узком (Миркин, 1978), когда под структурой понимается горизонтальное и вертикальное расчленение растительного покрова. Наиболее часто структура древостоев рассматривается как статистическое распределение особей древесных видов по диаметру. В основу методики и методологии наших исследований легли многие работы, начатые в европейской части России и в северных районах (Тарашкевич, 1923, 1924; Тюрин, 1927, 1971; Ипатов, 1968, 1969, 1971 и др.) и продолженные в сибирских районах (Фалалеев, Данилин, 1962; Дыренков, 1984, Аушев, 2004).
Сложность анализа прирусловых лесов Даурии состоит в высокой скорости динамических процессов, определяемых как климатическими и гидрологическими, так и пирогенными факторами естественного и антропогенного характера. Леса верхнего лесного пояса, испытывающие действие флювиальных процессов, представляют сложный комплекс, где сочетаются теневыносливые и светолюбивые виды, гигрофильные и ксерофильные синузии, криогенные и некриогенные участки. В этих условиях любые изменения гидрологического или температурного режима четко отражались на структуре экосистем. На структуру экосистем в зависимости от сезона и оказывают влияние как криогенные, так и стоковые (флювиальные) процессы различного характера: поверхностные, подрусловые, постоянные и временные водотоки.
Прирусловые таежные (бореальные) леса имеют следующие особенности: наличие устойчивого потока поколений в ценопопуляциях ели, кедра, лиственницы; высокое видовое богатство и видовую насыщенность; высокое синузиальное разнообразие; высокое структурное разнообразие (наличие высокой степени дифференциации древостоя по толщине, способствующей формированию структур «плотной упаковки»).
Эколого-ценотическая структура является важнейшим параметром мониторинга экосистем, поскольку под влиянием внешних (абиотических и антропогенных) и внутренних факторов любой ценоз претерпевает изменения. Эти изменения отражаются в первую очередь в структурной организации древостоя, который является важнейшим фактором устойчивости экосистемы. Принцип структурной дифференциации, рассмотренный рядом авторов (Работнов, 1992; Галанин и др., 2000; Вайс, 2006 и др.) общепризнан исследователями.
Характер флювиальных процессов и их влияние на формирование растительности
По характеру и результатам деятельности флювиальных процессов можно выделить три вида поверхностного стока вод: сток временных русловых потоков; плоскостной безрусловой склоновый сток; сток постоянных водотоков (Вернадский, 1933; Белый и др., 1977). Среди временных русловых потоков выделяются временные потоки оврагов и временные горные потоки. Для Сохондо характерны временные горные потоки. Их верховья расположены в верхней части горных склонов и представлены системой сходящихся многочисленных русел, рытвин и промоин, образующих в совокупности водосборный бассейн. Часто реки и ручьи протекают под обломочным материалом, представляя сеть подземных водотоков, растекающихся под каменной россыпью. Ниже по склону вода в ручье движется в едином русле - канале стока.

фото 1. Река Букукун в верховьях. На берег реки выходит сторона пп-17.Сложный пойменный кустарниково-разнотравно-зеленомошный елово-кедрово-лиственичный лес. |
В периоды сильных дождей и интенсивного таяния снега временные горные потоки движутся с большой скоростью и захватывают обломочный материал, который усиливает эрозионную деятельность. При выходе на предгорную равнину скорость движения уменьшается, горные потоки ветвятся на многочисленные рукава, в результате чего весь принесенный обломочный материал откладывается и образуется конус выноса временного горного потока, где местами наблюдаются дифференциация принесенного материала и зональность его распространения. В вертикальном разрезе отложений конусов выноса местами имеет место переслаивание мелко- и крупнообломочного несортированного, слабо окатанного материала.
Плоскостной безрусловый сток возникает в периоды выпадения дождей и таяния снега, когда вода стекает по склонам в виде сплошной тонкой пленки или густой сети отдельных струек. Они захватывают, главным образом, мелкоземистый материал, слагающий склоны и переносят его вниз. У подошвы течение воды замедляется, и переносимый материал откладывается как непосредственно у подножья, так и в прилегающей части склона (делювий). Для горных рек наиболее характерны непротяженные делювиальные отложения в связи с широким развитием гравитационных процессов на склонах. Наибольшая мощность делювия (до 0,5 м) наблюдается у основания склона, а вверх по склону она постепенно уменьшается. Продолжающийся процесс плоскостного смыва и образование делювия постепенно приводят к выполаживанию склонов.
Мощные водные потоки рек, расчленяющие горные склоны, производят значительную эрозионную, переносную и аккумулятивную деятельность. Это наиболее динамичные системы, преобразующие рельеф. Интенсивность работы рек определяется их живой силой, т. е. кинетической энергией, которая зависит от скорости течения. На интенсивности процессов в речных долинах сказывается турбулентный характер течения, перемешивающий всю массу воды от дна до ее поверхности. Наибольшие скорости наблюдаются в приповерхностной части потока на стрежне, меньше у берегов и в придонной части, где поток испытывает трение о породы, слагающие русло. |
В зависимости от характера и интенсивности питания изменяются режим рек, количество и уровень воды, а также скорость ее течения. В соответствии с изменением уровня воды в реке говорят о высоком уровне, соответствующем половодью, и низком меженном уровне, или межени, наступающей после спада половодья. Помимо этого, в реках наблюдаются периодические паводки, соответствующие кратковременному повышению уровня воды от затяжных дождей.
В Даурии для рек высокогорий характерно так называемое подрусловое течение, обусловленное глыбовым строением русел и сезонным питанием рек, что способствовало поверхностному расположению корневой системы деревьев и кустарников и особенности структуры таких экосистем. На растительные сообщества, кроме руслового и подруслового течения, воздействуют криогенные процессы, влияющие на мощность водного потока, длительность вегетационного периода прирусловых лесов, а в целом - на тип и структуру лесного сообщества.
Районы и объекты исследования
Климат СГБЗ резко континентальный, с господствующим сибирским антициклоном. Весна теплая, сухая, ветреная, с возвратными холодами, которые могут продлиться до июня. Летний период отличает влияние тихоокеанских муссонов, приносящих осадки с июля по август, когда выпадает до 80% годовой суммы осадков. Среднегодовая температура - 2,9° С. Продолжительность вегетационного периода около 90 дней, а у подножий 130-140. Снежный покров зимой достигает максимум 45-53 см, но почва промерзает на большую глубину. На территории распространена многолетняя мерзлота островного типа.
Исследования проведены в Сохондинском государственном биосферном заповеднике, в котором леса занимают 68% территории, часть лесов отнесена нами к прирусловым лесам. Изучались возрастной состав и дифференциация особей деревьев по размеру (обхват ствола на высоте 130 см), выявлялись видовой состав, ярусность, синузиальное сложение прирусловых лесов. Выбор спектра дефиниций и теоретических оснований классификации лесных экосистем и их динамики определился основополагающими лесоводческими работами В.Н. Сукачева (1964, 1975), трудами по генетической классификации лесной растительности Б.П. Колесникова (1956, 1958) и др.

Лиственично-кедровый кустарничково-разнотравный зеленомошный лес в урочище Верхний Букукун, ПП-5.
|

Лиственнично-кедровый баданово-брусничный с багульником лес в урочище Верхний Букукун, ПП-6.
|
Для исследования были выбраны наименее нарушенные участки таежных массивов реки Букукун в районе Букукунского озера, Букукунского Рудника, р. Сохондо. Характеристика и координаты ППП сведены в таблицу 1. Все обследованные участки расположены в зоне южной тайги. Основные характеристики климата, рельефа и растительности этой территории опубликованы (Васильченко, Галанин, 1983; Русских, 1983; Растительный..., 2002). Определение видового состава проводилось по сводке «Флора Сибири» (1999).
При выборе основных участков, на которых закладывались постоянные пробные площади, учитывалась репрезентативность этих участков определенным типам леса. Все площади располагались вблизи водных артерий, вдоль их притоков и в долинах небольших ручьев, включая ключи сезонного наполнения. Нами рассматривалась динамика кедровых и кедрово- лиственничных лесов верхнего лесного пояса Сохондинского биосферного заповедника. Для этого в заповеднике в 1982-1984 гг. была заложена, маркирована и описана система постоянных пробных площадей, работы с которой были возобновлены в 2000 г. и продолжаются в настоящее время (Беликович, Галанин, 2002; Беликович, Галанин, 2002; Галанин, Беликович, Галанина, 2002; Галанин, Беликович, 2004). Исследованный участок расположен в юго-западной части заповедника в бассейне р. Букукун на отрезке от села Букукун до озера Букукунского. Его протяженность составляет около 50 км и имеет сложное геологическое и геоморфологическое строение. Ширина поймы варьируется от 2-3 до 500 м.
На прирусловых отмелях в условиях длительного затопления и отложения руслового аллювия происходит формирование прибрежных ивняков. Для видового состава наиболее характерны Equisetum arvense, Inula britanica, Spiraea salicifolia. Прирусловые валы заняты ивняками и тополевыми лесами, доходящими до верхнего течения р. Сохондинки, до высот 1100—1200 м. н.у.м. На наносных прирусловых террасах обширные заросли состоят из Salix rorida и S . schwerinii . Для них типичны высокая сомкнутость крон и плотность древостоя, а напочвенный покров оказывается разреженным, с проективным покрытием не более 5-7%. В процессе сукцессии ивняки полностью замещаются тополевыми или березово-лиственничными лесами. На высоких редко затапливаемых наносных островах, образованных петлями и протоками р. Букукун в нижнем лесном поясе встречаются сосново- лиственнпчные (Pinus sylvestris и Larix gmelinii) разнотравно-злаковые леса. В них слабо развит подлесок из Padus avium, Swida alba, Rosa acicularis. Травяной ярус разнотравно-злаковый. В верхнем лесном поясе преобладают кедровые и кедрово-лиственничные леса, занимающие протяженные участки вдоль русел рек и ручьев.
Редкость и уникальность малонарушенных участков бореальных лесов для юго-восточной части Даурии обусловили выбор закладки ППП в экосистемах, типичных для прошлого столетия в этом регионе. Однако вследствие бесконтрольных рубок, климатических и антропогенных сукцессий, природных пожаров кедровые и кедрово-лиственничные леса стали редкостью. Сохранившиеся или восстановленные участки кедровников в заповеднике являются хорошими модельными объектами для изучения динамических процессов. Для этого мы проанализировали как старовозрастные леса, так и восстанавливающиеся на гарях под действием стоковых процессов. Выявление общих структурных черт таких лесов при понимании их природных различий мы понимаем как одну из задач настоящей работы.
Результаты и обсуждение
Временные водотоки
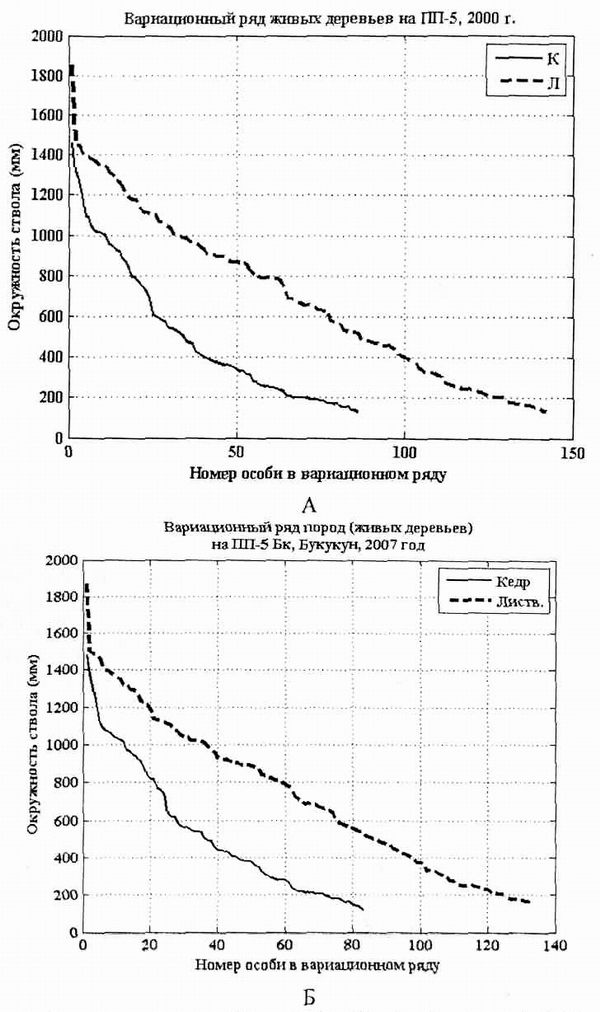
Рис. 1. Вариационные ряды Pinus sibirica (кедр) и Larix gmelinii (лиственница) в кедрово-лиственничном лесу в 200 г. (А) и в 2007 г. (Б) на пп-5, Верхний Букукун, ключ Загадочный. |
Ниже приводится характеристика изменений в прирусловых кедровых и кедрово-лиственничных лесах на постоянных пробных площадях, происшедших в результате смены климатических и гидрологических условий в верховьях р. Букукун.
ПП-5 заложена в 1983 г. А.В. Галаниным и А.В. Беликович. Размер ПП 50x50 м, она маркирована угловыми столбами с разбивкой на 25 квадратов размером 10x10 м, перечет древостоя выполнен в 2000 и 2007 гг., и видов сосудистых растений по квадратам. ПП заложена на склоне западной экспозиции, уклон 8 гр., в прирусловом (в приручьевом - в терминологии лесоустройства) сообществе, на ручье Загадочном - временном водотоке. Лес сформировался на гранитно-гнейсовых курумах. В описании 2000 г. лес характеризовался как кедрово-лиственничный с составом древостоя 7Л4К, III бонитета. Сомкнутость полога 60%, максимальная высота 18 м, средняя 14-16 м. Возобновление шло лиственницей, в кустарниковом ярусе доминировали жимолость, шиповник, багульник, брусника, в травяном покрове преобладали бадан, грушанка, седьмичник, вейник, княженика. Ревизия этой площади выполнена в 2007 г. Л.М. Долгалевой. В результате статистической обработки получены графики ценотичесой структуры древостоя в 2000 и 2007 гг. (рис. 1, А, Б). Вариационные ряды представляют собой убывающую последовательность окружностей стволов, скомпонованных по видам доминантам древостоя. Чем больше на пробной площади особей определенной породы дерева, тем длиннее вариационный ряд. Таким образом, оказывается, что в пионерном ценозе вариационный ряд длинный, в коренном короткий. Форма кривой (вогнутая, выпуклая, прямая), описывающей вариационный ряд показывает характер структуры ценопопуляции. Вогнутый вариационный ряд показывает интенсивное возобновление, сопровождающееся высокой степенью конкуренции. Выпуклая кривая вариации свидетельствует о постепенной деградации ценопопуляции, сопровождающейся низким возобновлением и высокой долей старых особей.
Вариационный ряд характеризовался доминированием лиственницы и замедленным пополнением основного древостоя из подроста. В ряду кедра количество особей с длиной окружности ствола 600-1000 мм меньше, чем в ценопопуляции лиственницы (рис. 1).
Повторное переописание показало, что в 2007 г. структура популяции кедра изменяется мало. Кривая вариационного ряда популяции лиственницы стала почти прямой за счет сокращения числа молодых особей и выпадения перестойных. Это обусловлено, по нашему мнению, сменой гидрологического режима ключа с надруслового, способствующего развитию оптимальных для лиственницы сырых условий произрастания на подрусловое, к которым в лучшей степени адаптирована популяция кедра, активизировавшая прирост.
В процессе развития ценоза в ходе дифференциации постепенно сформировались две дискретные группы особей кедров по толщине (рис. 2, А). Эти группы сохранились и в 2007 г. Процесс образования таких структурных элементов в ценопопуляции кедра связан с каким-то ритмическим процессом в возобновлении этого вида в данном фитоценозе, а также с выходом части особей кедра из нижнего яруса древостоя в верхний. |
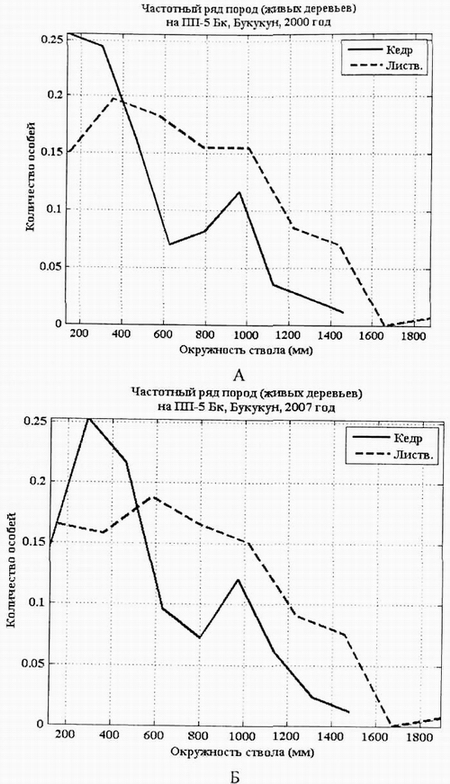
Рис. 2. Кривые распределения кедра сибирского и лиственницы даурской по диаметру в кедрово-лиственничном лесу на пп-5 в 2000 г. (А) и в 2007 г. (Б). |
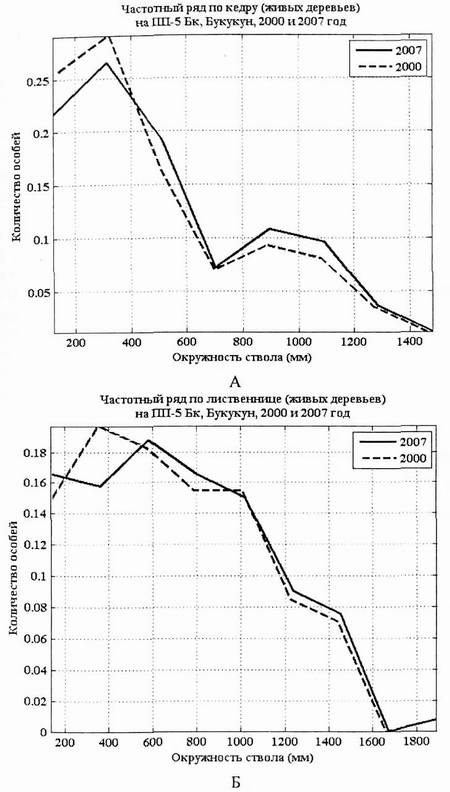
Рис. 3. Кривые распределения Pinus sibirica (А) и Larix gmelinii (Б) по диаметру в кедрово-лиственничном лесу на пп-5 в 2000 и 2007 гг.
|
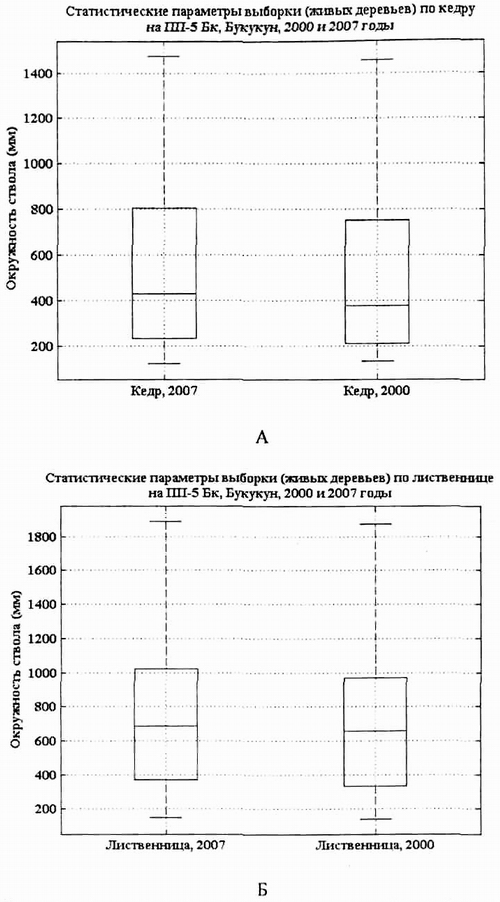
Рис. 4. Статистические параметры выборки живых Pinus sibirica (А) и Larix gmelinii (Б) на пп-5 в 2000 г. и в 2007 г. |
Ценопопуляция лиственницы по толщине дифференцирована слабее, чем ценопопуляция кедра. Правда, модальное значение у лиственницы за 7 лет сдвинулось вправо: с 390 мм в 2000г. до 590 мм в 2007 г. Такая структура характерна для ценопопуляций кедра и лиственницы в состоянии динамического равновесия экосистемы.
За 7 лет количество тонких особей кедра сократилось, но не за счет их гибели, а за счет перехода в группу с большим диаметром стволов. Семилетний период, очевидно, недостаточен для заметной перестройки структуры древостоя в этом типе леса (рис. 3, А, Б).
Наиболее четко прослеживается динамика процессов в схемах статистических выборок, называемых «ящиком с усами», где основные значения, 50% выборки графически ограничены прямоугольником, данные, выходящие за пределы выборки - точками или значками +, распределенная совокупность определяется отрезками (Глытина, Долгалева, 2009). Так, статистические параметры выборки на ПП-5 (рис. 4, А, Б) показывают увеличение прироста кедра, что подтверждается как повышением уровня медианы (средних значений толщин), так и увеличением длины «ящика».
Это происходит при выравнивании толщин особей, т.е. увеличении общего числа толстых групп особей в популяции. Заметна и асимметрия «ящиков» кедра (медиана находится ниже центральной области), что свидетельствует о нормальном пополнении тонких особей и активном приросте средних особей, выходящих в верхний ярус. Доминирующие кедры вариативны по толщине в диапазоне 800-1500 мм по обхвату и не выпадают из общей выборки, что характерно для стабильных популяций.
«Ящики» лиственницы за 7 лет практически не претерпевают изменений что свидетельствует как об усилении конкурентных взаимоотношений с кедром, так и об изменении экологических условий. Это подтверждается и натурными наблюдениями. На ПП-5 в 2007 г. произошла ксерофитизция экотопа. Временный водоток высох, период его наполнения и поверхностного стока, по наблюдениям сотрудников заповедника, стал короче вдвое. Изменение гидрологических условий отразилось и на травяном покрове: в сообществе появились Chamaenerion angustifolium, Vicia cracca, Artemisia sp., Achillea acuminata, Hedysarum alpinum, Astragalus membranaceus, Trifolium lupinaster, Vicia unijuga, характерные для умеренно сухих биотопов, а виды Bistorta vivipara, Ribes fragrans, Luzula rufescens, Aconilum cf. delphinipholium, Pedicularis labradorica, отмеченные на ПП в 2000 г. А.В. Галаниным, А.В. Беликович, выпали из травяного покрова фитоценоза в 2007 г. |
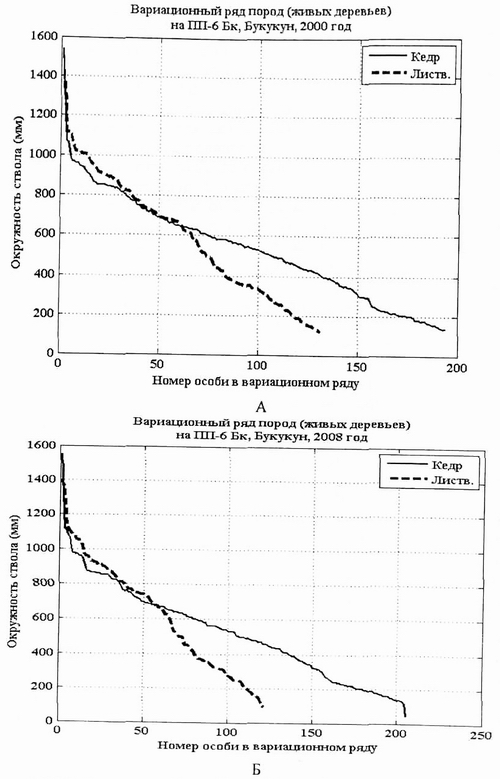
Рис. 5. Вариационный ряд Pinus sibirica (кедр) и Larix gmelinii (лиственница) в лиственнично-кедровом баданово-брусничном с багульником лесу на пп-6 в 2000 г. (А) и в 2008 г. (Б). |
ПП-6 заложена в 1983 г. А.В. Галаниным и А.В. Беликович. Листвененнично- Кедровый баданово-брусничный с багульником лес. Площадь расположена в окрестностях стационара Верхний Букукун в 100 м от зимовья под скалами на склоне западной экспозиции. Часть нагорной террасы в верхней части левого склона в реку Букукун, на территории бывшего рудника Верхний Букукун. Площадь расположена чуть ниже большого шахтного отвала, из которого вытекает водоток. Деятельность на руднике была прекращена в начале 60-х гг., однако и в настоящее время из шахты вытекает ручей, вода в котором обогащена минеральными элементами оловянных руд. Площадь расположена на участке вдоль ручья, который здесь уходит под камни. Площадь представляет собой местообитание с плоскостным подрусловым склоновым стоком. Лесной полог двухярусный. Формула древостоя 5К5Л. Бонитет древостоя III . Древесный полог вполне сформирован. Высота 1 яруса 18-20 м, 2 ярус - 14-15 м. Сомкнутость 0,7-0,8. В возобновлении преобладает кедр, лиственницы значительно меньше. Напочвенный покров неоднороден, имеются мохово-кустарничковые, бадановые и бруснично-зеленомошные парцеллы. Кустарников очень мало, сколь либо выраженного яруса они не образуют. Выходов камней на поверхность очень мало. В пределах пробной площади выделяются две фации: фация 1 - дно лога, фация 2 - склон лога (борт юго-западной экспозиции).
На площади обильны следующие виды травянистых растений: Berrgenia crassifolia, Maianthemum bifoiium, Goodyera repens, Carex pallida, Carex cf . tenuifolia, кустарники и кустарнички: Ledum palustre, Rhodococcum vitis-idaea, Lonicera pallasii, Spiraea flexuosa и др. Структура ревостоя такого леса отражена в графических схемах вариационных рядов кедра и лиственницы. Кривые вариационных рядов за 7 лет изменись очень мало (рис. 5, А, Б). Это позволяет считать данный фитоценоз коренным.
Особи кедра и лиственницы, заняв свои экологические ниши, практически не испытывают дефицита ресурсов. Склоновые потоки, равномерно распределяясь по поверхности, питают влагой как молодые, так и перестойные особи. Делювиальные процессы способствуют накоплению гумуса в нижней части склона, подпитываемого также ручьем, стекающим из рудника. Такие условия идеальны для лиственницы, которая преобладает в нижней части склона. Умеренно увлажненные, дренированные склоны, заняты преимущественно особями медленно растущего кедра, для которого экологически выгоднее как дренаж, так и лучшее освещение при склоновом размещении.
За 8 лет более четкой стала дифференциация особей лиственница на две группы по толщине ствола. Такой процесс характерен для относительно стабильных фитоценозов. Статистические параметры выборки деревьев кедра и лиственницы (рис. 7) в этом фитоценозе, рассмотренные в их динамике, подтверждают такой вывод. |
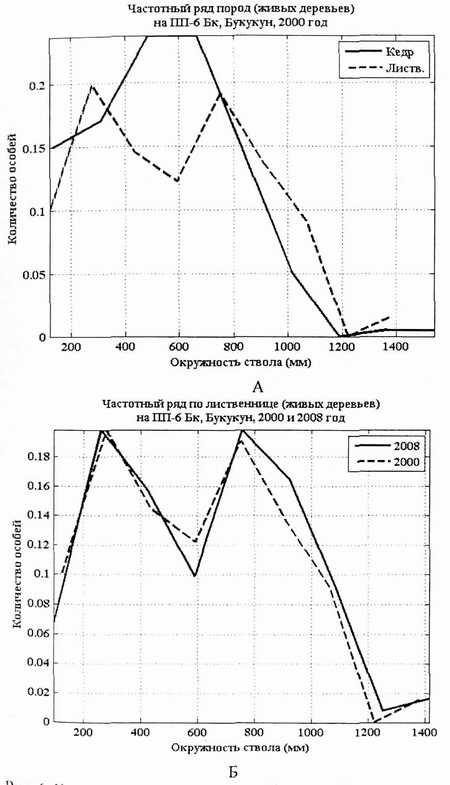
Рис. 6. Кривые распределения кедра сибирского (А) и лиственницы даурской (Б) по длине окружности ствола (мм) на пп-6 в 2000 г. и в 2008 г. |
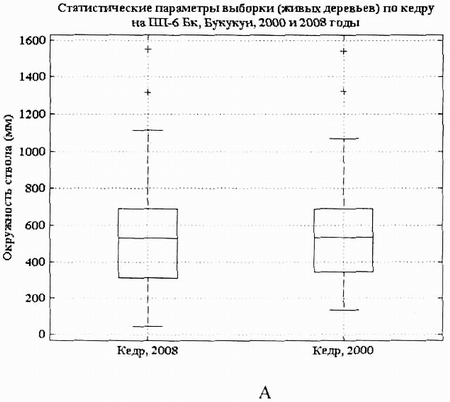
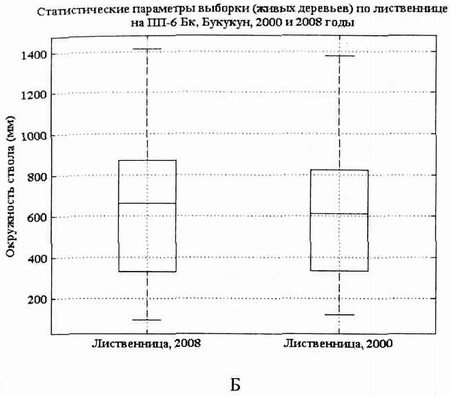
Рис. 7. Статистические параметры выборки живых Pinus sibirica (А) и Larix gmelinii (Б) на пп-6 в 2000 г. и в 2007 г.
|
Диаграммы выборок кедра и лиственницы в виде «ящиков» также подтверждают вывод об относительной стабильности данной фитоценотической системы. Судя по этим диаграммам, позиции лиственницы в данном фитоценозе более прочные, чем кедра. В 2008 г. сохранился уровень медианы у кедра, но увеличился у лиственницы. В ценопопуляции лиственницы в 2008 г. медиана сместилась вверх за счет интенсивного прироста особей в основной части выборки. Нижняя асимметрия «ящика» показывает пополнение популяции за счет тонкомера.
Постоянные водотоки
ПП-17 заложена в 1984 г. А.В. Галаниным и А.В. Беликович. Сложный пойменный кустарниково-разнотравно-зеленомошный елово-кедрово- лиственичный лес. Таксация древостоя с картированием деревьев впервые проведена в 2001 г. Тогда же площадь была разбита на 25 квадратов размером 10x10 м, был выполнен перечет видов сосудистых растений, синузий мхов и эпигейных лишайников по квадратам. Площадь расположена на правом берегу р. Букукун против зимовья «Букукунское озеро» примерно в 150 м от него. Площадь неоднородна, она захватывает берег реки - прирусловую часть поймы и более высокий участок поймы, постепенно переходящей в низкую и слабо выраженную надпойменную террасу, первая фация - это прирусловый разнотравный кустарниковый (ивняковый) лес. Вторая фация отличается почти полным отсутствием высоких кустарников и слабым развитием травяной синузии. Третья фация по пощади незначительная, она с преобладанием кустарничков, зеленых мхов и кустистых эпигейных лишайников. Во второй фации (ерниковой) в последние 20 лет вдет массовое возобновление ели, древостой здесь имеет формулу 9Л1Е, а возобновление 8К1Л1Е. Следовательно, на площади происходит постепенная смена пород - лиственница замещается кедром, активизируется ель. В 2000 г. при переописании растительности на этой площади были отмечены падение уровня грунтовых вод, смена лесообразующих пород (становится больше кедра и ели), в приречной фации происходит обеднение видового состава не только трав, но коснувшееся даже кустарников, в ерниковой фации снизили свое обилие голубика и брусника, исчезли 15 видов разнотравья.
Переописание ПП-17 в 2008 г. позволило построить следующие графические модели структуры древостоя. Вариационный ряд популяций кедра отличает сильная вогнутость (почти ломанность) кривой вариационного ряда, Это говорит об агрессивности данной ценопопуляции, ее активном возобновлении. В ценопопуляции кедра много особей, которые «ждут» момента, чтобы выйти в верхний ярус. Вариационный ряд лиственницы в данном фитоценозе также вогнутый, что говорит об устойчивости данной ценопопуляции в фитоценозе.
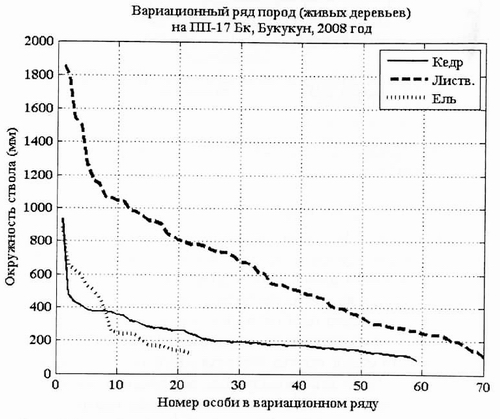
Рис. 8. Вариационный ряд кеда сибирского, лиственницы Гмелина ели на ПП-17 в 2008 г. |
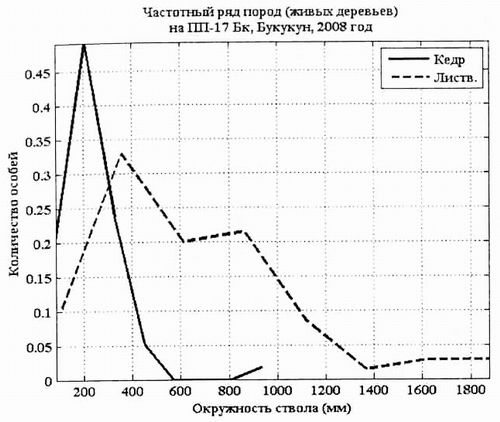
Рис. 9. Кривые распределения кедра сибирского и лиственницы сибирской по обхвату (мм) на ПП-17 в 2008 г. (Б). |
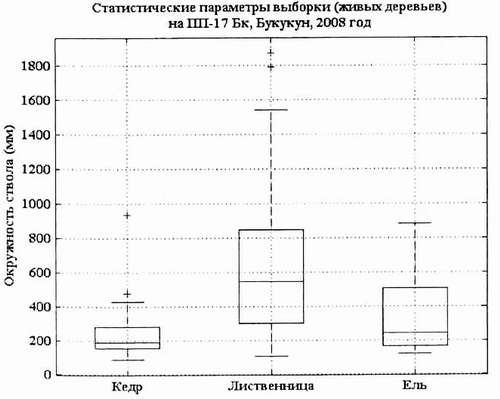
Рис 10. Статистические параметры выборки живых Pinus sibirica, Larix gmelinii и Picea obovata па пп-17, пойма р. Букукун в 2008 г. |
Статистические параметры ценопопуляции кедра и лиственницы, отраженные на диаграмме «ящик» показывают, что фитоценоз находится в относительно устойчивом состоянии, при котором лидирует лиственница, на втором месте кедр и на третьем ель сибирская.
Выводы
Таким образом, горные кедровые и кедрово-лиственничные леса верхнего лесного пояса Сохондинского биосферного заповедника, находящиеся в условиях существенного воздействия флювиальных процессов, отличаются относительной устойчивостью и могут рассматриваться как коренные. Устойчивой структурой обладают кедровые леса в местообитаниях с плоскостным стоком, где условия увлажнения изменяются незначительно.
В устойчивых лесных экосистемах по результатам нашего мониторинга сохраняется тенденция к усложнению ценотической структуры. Устойчивость ценопопуляций кедра и лиственницы в изученных фитоценозах определяется двумя факторами: относительной стабильностью режима
увлажнения и регулярным пополнением ценопопуляций видов деревьев за счет возобновления и за счет пропорционального роста деревьев.
Для выявления более существенных изменений в сообществах коренных кедрово-лиственичных лесов Сохондинского заповедника необходим более длительный мониторинг древостоев на постоянных пробных площадях.
Использованные нами методы статистической обработки данных дают вполне удовлетворительные результаты и способствуют пониманию
процессов, происходящих в прирусловых лесах. |
Литература
Аушев С.В. Экологические зависимости влияния факторов внешней
среды на интенсивность прироста сосны обыкновенной. Автореф. дис. на соиск. учен. степ. канд. биол. наук: - Томск: 2006. - 22 с.
Банникова И.А., Худяков О.Н. Почвенно-растительные подпояса
юго-восточного Хангая // Структура и динамика основных экосистем МНР.
- Л., 1976.-С. 5-134.
Беликович А.В., Галанин А.В. Посинузиальная структура растительного покрова Сохондинского биосферного заповедника // Флора, рас
тительность и растительные ресурсы Забайкалья. Т. 1. - Новосибирск: изд-
во СО РАН, 2000. - С. 47-52.
Беликович А.В., Галанин А.В. Забайкальский харганат как тип растительности // Комаровские чтения. - Владивосток: Дальнаука, 2005. Вып.
39: С. 162-185.
Белый Б.В., Беркович К.М., Зайцев А.А., Рулева С.Н., Чалов Р.С.
Исследование русловых процессов на больших реках Сибири // Геоморфология. - 1977. - № 3. - С. 3-11.
Бернадский Н.М. Речная гидравлика, её теория и методология. Т. 1.
- Л. - М: Госэнергоиздат, 1933. - 148 с.
Вайс А.А. Структурная организация древостоя // Лесные экосистемы Северо-Восточной Азии и их динамика: Матер, междунар. конф. - Владивосток: Дальнаука, 2006. - С. 35-37.
Васильченко З.А. Высокогорная флора Сохондинского заповедника // Природа Сохондинского заповедника. - Чита, 1983. - С. 38-58.
Васильченко З.А. Ботаническая изученность Сохондинского заповедника. // Ботанические исследования в заповедниках РСФСР. - М., 1984,
С. 34-38.
Васильченко З.А. Надземная биомасса травяно-кустарничкового яруса типичных сообществ лесостепного пояса Сохондинского заповедника. // Опыт исследования растительных сообществ в заповедниках. - М., 1988.-С. 72-75
Васильченко З.А. Редкие растения Сохондинского заповедника. //
Редкие виды растений в заповедниках. - М., 1987. - С. 46-56.
Васильченко З.А., Шевырева Н.А. Новые и редкие виды семейства
мятликовых Сохондинского заповедника. // Опыт исследования растительных сообществ в заповедниках. - М., 1988.-С. 115-120.
Васильченко А.А., Галанин А.В. Физико-географические условия и
история создания Сохондинского заповедника. // Природа Сохондинского
заповедника. - Чита, 1983. - С. 3-7.
Виппер П.Б. Взаимоотношения леса и степи в горных условиях
Юго-Западного Забайкалья // Ботан. журн. 1968. Т. 53, № 4. - С. 491-504.
Галанин А.В. Перспективы ботанических исследований в Сохондинском заповеднике. // Ботанические исследования в заповедниках
РСФСР. - М., 1984. - С. 39^3.
Галанин А.В. Ценотическая организация растительного покрова. -
Владивосток: ДВО АН СССР, 1989. -163 с.
Галанин А.В. Флора и ландшафтно-экологическая структура растительного покрова. - Владивосток: ДВО АН СССР, 1991. - 272 с.
Галанин А.В., Беликович А.В. Эколого-флористическая структура
растительного покрова Сохондинского заповедника (Хэнтей-Чикойское
нагорье) // Структурная организация компонентов биогеосистем (сравнительный и количественный анализ). - Владивосток: ДВО АН СССР, 1988.
- С. 11-23.
Галанин А.В., Беликович А.В. Динамика древостоя в субальпийском кедровнике в Сохондинском заповеднике за 24 года // Классификация
и динамика лесов Дальнего Востока. - Владивосток: Дальнаука, 2001.
Галанин А.В., Беликович А.В. Предварительный список видов сосудистых растений Сохондинского биосферного заповедника и его окрестностей // Растительный и животный мир Сохондинского биосферного заповедника. - Чита-Владивосток: БСИ ДВО РАН, СГБЗ,
2002. - С. 79-106.
Галанин А.В., Беликович А.В. Изменения в растительном покрове Сохондинского заповедника по результатам ревизии геоботанических пробных площадей (1983-2001) // Растительный и животный мир Сохондинского биосферного заповедника. - Чита-Владивосток: БПИ ДВО РАН, БСИ ДВО РАН, СГБЗ, 2002. - С. 14-33.
Галанин А.В., Беликович А.В. Изучение фитогенного поля сосны
обыкновенной в Южном Забайкалье // Растительный и животный мир Сохондинского биосферного заповедника. - Чита-Владивосток:
БСИ ДВО РАН, СГБЗ, 2002. - С. 53-62.
Галанин А.В., Беликович А.В. Постоянные геоботанические пробные площади Сохондинского биосферного заповедника. - Чита: Поиск,
2004. - 228 с.
Галанин А.В., Беликович А.В., Галанина И.А. Анализ растительности лиственнично-березового леса в долине р. Агуца// Растительный и животный мир Сохондинского биосферного заповедника. - Чита-Владивосток: БСИ ДВО РАН, СГБЗ, 2002. - С. 34-52.
Галанин А.В., Проскурина Н.С. К вопросу о фенологической периодизации года в Южном Забайкалье // Природные ресурсы Читинской
области. - Чита: Заб. отд. ВГО, 1983. - 126 с.
Галанин А.В., Беликович А.В., Храпко О.В. Флора Даурии. - Владивосток: Дальнаука, т. 1, 2008.- 183с.
Долгалева Л.М. Динамика древостоя кедрово-лиственничного леса
в верхнем лесном поясе Сохондинского биосферного заповедника
// Бюллетень БСИ ДВО РАН, вып. 4, http://botsad. ru/journal/number.htm,
2009а. - С. 50-53.
Долгалева Л.М. Экологическая оценка прирусловых лесных сообществ бассейна р. Букукун в Сохондинском государственном биосферном
заповеднике // Трансграничные территории Азиатской части России и сопредельных государств: геоэкологические и геополитические проблемы и
предпосылки устойчивого развития. - Улан-Удэ: Изд-во БГУ, 20096. -
С.100-102.
Дулепова Д.И. Флора степных и лугово-болотных сообществ и ее
особенности. // Природа Сохондинского заповедника. - Чита, 1983. -
С. 33-37.
Дулепова Б.И. Петрофильные кустарниковые степи, их значение и
охрана // Охрана природы и воспроизводство естественных ресурсов. -
Чита, 1979. - С. 95-97
Дулепова Б.И., Уманская Н.В. Динамика растительных сообществ в Сохондинском заповеднике // Флора, растительность и растительные ресурсы Забайкалья. Т. 1. - Новосибирск: изд-во СО РАН, 2000. - С. 58-66.
Дулепова Б.И., Уманская Н.В. Болота // Природа Сохондинского заповедника. - Чита: СГЗ, 1983.-С. 97-100.
Дылис Н.В., Рещиков М.А., Малышев Л.И. Растительность // Предбайкалье и Забайкалье. - М.: Наука, 1965.- С. 225-281.
Дыренков С.А. Структура и динамика таежных ельников. - Л.,
1984. - 174 с.
Ипатов B.C. Дифференциация древостоя I // Вести. ЛГУ. Биология.
1968. - №21. - С. 59-68.
Ипатов B.C. Дифференциация древостоя II // Вестн. ЛГУ, Биология.
1969,-Вып. З. - С. 43-54.
Ипатов B.C. Исследование структуры растительных сообществ. Авторефер. дис. ... д-ра биол. наук. - Тарту, 1971. - 55 с.
Ипатов B.C., Кирикова Л.А. Фитоценология: учебник. - СПб.: Изд-во С-Петербург. ун-та, 1999. - 316 с.
Касич В.Б. Леса юго-восточной части заповедника // Природа Сохондинского заповедника. - Чита: СГЗ, 1983. - С. 100-104.
Коротков И.А. Типы леса Монгольской Народной Республики / Леса МНР. - М.: Наука, 1978. - С. 47-121.
Колесников Б.П. Состояние советской лесной типологии и проблемы генетической классификации типов леса. - Изв. Сибир. отд-ния АН СССР.- 1958. - № 2. - С. 109-122.
Лесная энциклопедия: В 2-х т., т.2 / Гл. ред. Воробьев Г.И.; Ред. кол.: Анучин Н.А., Атрохин В.Г., Виноградов В.Н. и др. - М.: Сов. энциклопедия, 1986. - 631 с.
Мазинг В.В. Теоретические и методические проблемы изучения структуры растительности: Докл. по опубл. работам, представл. к защите вместо дис. на соиск. учен. степ, д-ра биол. наук. - Тарту, 1969. - 95 с.
Миркин Б.М. Теоретические основы современной фитоценологии. - М.: Наука, 1985. - 136 с.
Паллас П.С. Путешествие по разным провинциям Российской империи 1772 и 1773гг. Кн. I и II. - СПб., 1788.
Растительный и животный мир Сохондинского биосферного заповедника: Труды Сохондинского биосферного заповедника. Вып. 2. - Чита:
Изд-во СБЗ, 2002. - 170 с.
Русских A . M . Климат и микроклимат. // Природа Сохондинского
заповедника. - Чита, 1983. С. 10-12.
Сипливинский В.Н. Флористические находки на горе Сохондо (Забайкалье) // Новости систематики высших растений. - 12, 1968. - С. 287-303.
Соколов Н. П. Описание горы Чеконды на китайской границе //
Собр, сочинений, выбранных из месяцесловов на разные годы. - СПб., 1793, ч. 10. - 337.
Сукачев В.Н. Динамика лесных биогеоценозов // Основы лесной
биогеоценологии, - М., 1964. - С. 458-486.
Тарашкевич А.И. К вопросу о таксации северных лесов, состоящих
из деревьев различных поколений // Лесопромышленное дело. 19236. -
№ 1-16. - С. 18-21.
Таран Г.С. Сукцессионные смены растительности в пойме средней
Оби при свободном меандрировании // Человек и вода. Тезисы докл. к
научно-практич. конф. - Томск, 1990. - С. 138-141.
Таран Г.С. К характеристике пространственной структуры растительности в пойме средней Оби // Флора и растительность Сибири и Дальнего
Востока. Тезисы докл. конференции. - Красноярск, 1991. - С. 130-132.
Таран Г.С. Синтаксономический обзор лесной растительности.поймы средней Оби (Александровский отрезок) // Сиб. биол. журн., 1993, вып.
6. - С. 85-91.
Таран Г.С. Малоизвестный класс растительности бывшего СССР -
пойменный эфемеретум (Isoeto - Nanojuncetea Br.-BI. et Tx. 43) // Сиб. экол.
журн. 1995. Т. 2, №4. - С. 373-382.
Таран Г.С. Осиновые и березовые леса поймы средней Оби ( Spiraeo
salicifoliae-PopuIetum tremulae Taran 1993) // Ботанические исследования
Сибири и Казахстана. Сборник научных статей. Вып. 4 / Под ред.
А.Н. Куприянова. - Барнаул: Изд-во АГУ, 1998. - С. 82-89.
Таран Г.С., Тюрин В.Н. К характеристике флоры и растительности
поймы Оби в окрестностях Сургута // Проблемы изучения растительного
покрова Сибири. Тезисы докладов II Российской научной конференции. -
Томск: Изд-во ТГУ, 2000. - С. 140-141.
Уманская Н.В. Луга // Природа Сохондинского заповедника. - Чита: СГЗ, 1983. - С. 84-97.
Фалалеев Э.Н., Данилин М.А. Строение осиновых насаждений Сибири // Лесное хозяйство. - 1962, №10. - С. 5-9.
Флора Сибири. - Новосибирск: Наука. Сиб. отд-ние. 1990-2000 -
Т. 1-14.
Черкасов А.А. Записки охотника Восточной Сибири. - СПб 1867, 1884.


