Устойчивость и динамика экосистем
Галанин А.В., Беликович А.В.
Ботанический сад-институт ДВО РАН
 |
© OCR А.В. Галанин, Беликович А.В. Публикуется по: А.В. Галанин, Беликович А.В. Устойчивость и динамика экосистем // Экосистемы Сибири и Дальнего Востока. Находка: ИТИБ, БСИ ДВО РАН, 2010. С. 7-20.
В основу статьи положены результаты 30 летних исследования авторов в Даурии, Севере и юге Дальнего Востока.
В статье рассматриваются некоторые основные понятия теории экосистем - такие, как структура, биоразнообразие, динамика, сукцессиоиные смены, необратимые изменения растительных сообществ, климатогенные изменения флоры и растительности. Анализируются результаты длительного мониторинга структуры древостоя на постоянной пробной площади в Сихотэ-Алинском биосферном заповеднике. Проведен сопряженный анализ ареалогической и эколого-ценотической структуры дендрофлоры Даурии, определены главные векторы развития растительного покрова Даурии. |
Экосистемы Дальнего Востока России и Сибири изучены очень неравномерно и явно недостаточно. Это постоянно сказывается тогда, когда требуется предсказать экологические последствия того или иного крупного строительства, будь то прокладка нефтепровода из Сибири в Приморье и Китай или крупное строительство на острове Русском во Владивостоке. Мы до сих пор точно не знаем, как поведет себя экосистема Южного Приморья в результате длительного и постоянного прохождения лесов низовыми пожарами или в результате устойчивого глобального изменения климата (Krupa, 1997).

Фото 1. Пихтовый (Abies nephrolepis) лес на юге острова Сахалин. Фото А.В. Галанина. |
Общепризнано, что биоразнообразие играет большую роль в поддержании устойчивого состояния в биосфере и отдельных экосистемах. Именно поэтому поддержанию и сохранению высокого биоразнообразия экосистем уделяется такое огромное внимание. Создание Красных книг, охрана редких растений и животных в заповедниках и ботанических садах, создание банков семян - все это подтверждает сформулированный выше тезис.
Однако, несмотря на очень большое число публикаций на тему биоразнообразия, повсеместное издание региональных Красных книг, теоретические аспекты этой проблемы раскрыты весьма поверхностно. Разработка количественных методов для оценки биоразнообразия - это только первый робкий шаг в сторону разработки теории биоразнообразия. Чтобы создать такую теорию, необходимо ответить, по крайней мере, на следующие вопросы: 1) Что такое устойчивость экосистем? 2) С помощью каких критериев можно отличать устойчивые экосистемы от неустойчивых? 3) Всегда ли высокая устойчивость экосистем связана с их высоким биоразнообразием? 4) Почему в одних биоценозах много видов, а в других мало? 5) Почему в сообществах обильными являются немногие виды, в то время как многие встречаются единично или в малом обилии? 6) Как связано изменение биоразнообразия в биосфере и конкретных экосистемах с процессами видообразования и биологической эволюцией?
Чтобы убедительно ответить на эти вопросы, необходимы конкретные исследования многих специалистов. В одной небольшой статье можно лишь |
наметить основные подходы к обозначенным этими вопросами проблемам. Важнейшую, я бы сказал, базовую роль в экосистемах и в биосфере в целом играет растительный покров или растительный мир, который, как известно, является в ней восходящей ветвью биологического круговорота (а именно, синтез и накопление органического вещества). Устойчивой будет такая экосистема, в которой восходящая ветвь биологического круговорота равна нисходящей. Баланс вещества в биологическом круговороте - вот ключ к пониманию сути устойчивости экосистем. Если баланс биологического круговорота нарушить, то равновесие в экосистеме нарушится, ее фитоценоз, зооценоз, микоценоз и микробоценоз начнут и будут изменяться до тех пор, пока не наступит новое состояние равновесия между восходящим и нисходящим потоками вещества.
Устойчивость является одним из ключевых свойств сообществ. Это состояние в существовании сообщества достигается, с одной стороны, разнообразием входящих в его состав членов, с другой - наибольшей их приспособленностью как друг к другу, так и условиям существования. Видовое и ценотическое разнообразие приводит к более устойчивому положению растительного сообщества, позволяя ему существовать длительное время в состоянии равновесия. Любые нарушения, вызванные внешними факторами или изменениями внутренней среды, приводят к закономерной динамике структуры сообщества.

Фото 2. Дубовый (Quercus mongolica) лещиново-леспедециевый (Corylus mandzhurica + Lespedeca bicolor) лес в Приморском крае на Борисовском плато. А.В. Галанин читает лекцию аспирантам о биоразнообразии. Фото Е.Н. Роенко. |
Оценка эколого-ценотического разнообразия растительного покрова позволяет дать характеристику состояния экосистемы в конкретный момент времени, а при периодических наблюдениях определить направленность происходящих в ней процессов. Очевидно, что существующее сообщество сохраняет свою уникальность, то есть чем-то отличается от других таких сообществ и сохраняет эти отличия во времени. Именно в процессе динамики идет выработка упорядоченности в структуре сообществ растений и животных и выявляются закономерности процесса самоорганизации живой материи (Ипатов, 1969). Одним из элементов, составляющих лесную экосистему и лесное растительное сообщество, является древостой, структура которого в основном определят всю структуру фитоценоза. Структура в данном случае понимается как некоторая упорядоченность компонентов биоценоза: особей, жизненных форм организмов и видов. Разнообразие частей, составляющих систему, - это условие возникновения, сохранения и развития структуры. Образно говоря, структура это ловушка для негэнтропии, поступающей в систему из окружающей среды: в структуре системы негэнтропия накапливается, сохраняется и превращается в информацию.
В естественных древостоях структурирование не ограничивается различием особей по их принадлежности к определенным различным видам. Более широкая амплитуда различий растений в пределах вида по фенотипическим признакам поддерживается генетически и проявляется в индивидуальной силе роста и дифференциации особей по мере их роста, которая сопровождается отпадом слабых экземпляров. Эта дифференциация и является основой устойчивости всего фитоценоза. В результате дифференциации особей одного вида в древостое в ценопоггуляции вида возникают разные |
совокупности (группы) деревьев по толщине, высоте, мощности крон, биохимической эффективности. Динамика лесного фитоценоза и процесс дифференциации его элементов, кроме различия в генотипе особей, определяются также и характером их размещения на площади относительно друг друга. Анализ структурных образований коренных и производных насаждений дает неизмеримо большую информацию о природе изучаемой лесной экосистемы и подлинном разнообразии сообщества живых организмов. Этот метод более полно отражает ценотическую структуру сообщества, чем метод суммарного описания типа леса в виде различных усредненных индексов (средней высоты и диаметра, состава и полноты древостоя). Фактические данные о ценотических отношениях в описанных растительных группировках являются первым этапом изучения многолетней динамики ценотической структуры естественных растительных группировок. Поэтому целью эколого-ценотического исследования следует считать закономерности

Фото 3. Маньчжурская разнотравная прерия на юге Приморского края. Борисовское плато. Фото А.В. Галанина. |
процесса дифференциации особей в ходе восстановительной динамики, а также особенности пространственного размещения особей в пределах фитогенных полей их соседей.
Одним из основных вопросов фитоценологии является изучение процессов самоорганизации растительных сообществ как части более сложных систем - биоценозов и экосистем. Для растительного сообщества, так же как и для всех открытых нелинейных систем, характерны устойчивость относительно внешних воздействий, самообновляемость, возможность к самоусложнению, росту, развитию и согласованность всех составных частей. Изучая сложные самоорганизующиеся системы, необходимо делать акцент на внутренние свойства как на источник саморазвития и познавать функционирование элементов. Например, благодаря дискретности жизни и эколого-биологической неравноценности особей растений даже на совершенно однородном участке формируется неоднородная система, микроструктура которой обеспечивает более оптимальный ход биогеохимического процесса экосистемы в целом (Галанин, 1989). Факторы формирования внутренней неоднородности фитоценозов, связанные с жизнедеятельностью растений, могут быть названы фитогенными, топическими, ценотическими, фитоценотическими. Ценотические отношения могут проявляться в изменении состояния взаимодействующих растений, в изменении успешности размножения, в неслучайном распределении особей разных видов относительно друг друга, в изменении видового состава и количественных соотношений между видами в фитоценозе (Галанин, 1989). |
Образование новых элементов в растительном сообществе есть результат упорядочивания ценофункциональных связей, а наличие таких элементов составляет ценотическую организацию растительного покрова. Такую внутреннюю структуру с ее элементами и связями, которая стремится к самосохранению в некоторых границах допустимых возмущений, по-видимому, можно считать одним из «аттракторов» (или целей системы), способных при достаточном наборе видов сформировать на данном участке биогеохимически оптимальный фитоценоз (Галанин, 1989, 1991).
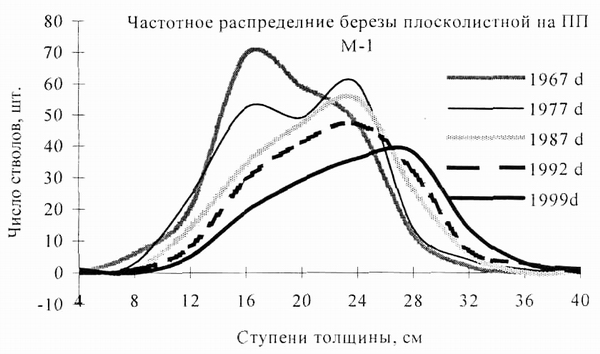
Рис. 1. Динамика дифференциации ценопопуляции березы плосколистной на постоянной пробной площади в Сихотэ-Алинском заповеднике (по Н.А. Васи ленко, 2008) |
На рис. 1 приведена серия кривых частотного распределения особей березы плосколистной на пробной площади в Сихотэ-Алинском заповеднике, заложенной в окрестностях кордона Майса в 1967 г. Были проведены 4 ревизии пробной площади в 1977, 1987, 1992 и 1999 гг. Отчетливо видно, как изменялась ценопопуляция березы по мере восстановления лесной экосистемы, которая в 50-е гг. прошлого века была выведена из состояния равновесия верховым лесным пожаром, уничтожившим кедрово-широколиственный древостой. В 1967 г. ценопопуляция пионерного вида березы плосколистной уже вполне сформирована: средний диаметр особей около 20 см. В ценопопуляции березы началась дифференциация особей по толщине: можно выделить два модальных значения диаметра - 16 см и 23 см. В 1977 г. степень дифференциации березы в результате межвидовой конкуренции на пробной площади достигла максимума: сформировались две совокупности деревьев - с толщиной ствола 16 см и 24 см. Через 10 лет, в 1987 г. особи, отставшие в росте (диаметр ствола 16 см), элиминировали, и соответствующая деформация кривой исчезла. Самые успешные особи увеличили диаметр, но модальное значение, равное 24 см, не изменилось, в результате чего кривая распределения стала асимметричной. Ценопопуляция березы в период с 1977 по 1978 гг. стала деградировать, этот процесс продолжался и в дальнейшем. В 1992 г. модльное значение |
диаметра ствола остается прежним, но численность особей в ценопопуляции заметно снижается. Наконец в 1999 г. численность ценопопуляции березы снизилась еще больше, но оставшиеся особи прибавили в росте, в результате чего модальное значение диаметра ствола стало равно 28 см, и правая часть кривой заметно сместилась вправо. Активизация роста особей березы в период с 1992 по 1999 гг. произошла в связи с ослаблением внутривидовой конкуренции (много особей березы к этому времени элиминировали), а также в связи с климатическими изменениями, в результате которых экологические условия для березы плосколистной на данной пробной площади стали более благоприятны.

Фото 4. Забайкальский край. Вершина священной горы Алханай. Лиственичная редина (Larix dahurica) и заросли кедрового стланика (Pinus pumila). Фото Л.М. Долгалевой. |
Выведенная из состояния равновесия локальная экосистема со временем восстанавливается (вновь возвращается в состояние, близкое к исходному), проходя при этом некоторый ряд этапов (или цикл демутаций), или не возвращается, а переходит в новое состояние, отличное от исходного по главным параметрам (в том числе и по набору видов и соотношению их обилий, по количеству вещества, вовлеченного в биологический круговорот, по скорости оборота этого вещества, по количеству энергии, которую растительное сообщество захватывает в процессе фотосинтеза и которую в дальнейшем используют животные, грибы и бактерии, обитающие в данной экосистеме). Выводя экосистему из состояния равновесия, мы должны знать тот предел воздействия, после которого экосистема не может возвратиться к исходному состоянию, а перейдет в новое.
В геоботанике хорошо изучены сукцессионные смены растительных сообществ в пределах экосистемы, выведенной из состояния равновеия. Они соответствуют циклам демутаций локальной экосистемы, после которых система возвращается в состояние, близкое к исходному - к так называемой коренной растительности. Санитарные и выборочные рубки в лесу являются примером таких воздействий на экосистему, после которых она может восстановить утраченное динамическое равновесие. Естественные экосистемы из состояния динамического равновесия чаще всего выводятся при лесных и степных пожарах, рубках сплошного пользования и др. Коренная растительность в результате демутаций восстанавливается, но далеко не всегда. Если внешняя среда была изменена очень существенно (смыло почвенный слой, изменилась крутизна склона, снизился уровень грунтовых вод, изменился климат и т.д.), |
прежняя экосистема и ее компоненты (фитоценоз, зооценоз, микоценоз и микробоценоз) восстановиться не могут. Новое коренное сообщество на этом участке будет отличаться от коренного первоначального. Другая причина невозможности восстановления исходного коренного растительного сообщества может быть связана с отсутствием в данной локальной флоре источника диазачатков видов, составлявших коренное сообщество до его разрушения. Это может случиться тогда, когда коренная растительность уничтожается на очень большой территории, соизмеримой по размерам с площадью локальной флоры. Например, верховым пожаром уничтожен сосновый лес на площади в 2-3 тыс. км . При этом сосна как вид исчезла на этой территории, источника семян нет, и прежний тип леса здесь восстановиться не может даже по прошествии нескольких сотен лет.
В ряде случаев достижение исходного состояния растительности невозможно в связи с нестабильностью климатических условий. Климат региона изменяется быстрее, чем длится цикл демутаций коренной растительности. В таких условиях коренная растительность просто не успевает восстанавливаться. Компонентом экосистем в этом случае постоянно является пионерная растительность, а сами экосистемы находятся в перманентном неравновесном состоянии. Примером такой растительности является современная растительность Дальнего Востока, находящаяся в условиях неустойчивого муссонного климата. Похоже, что именно перманентная нестабильность биологического круговорота вещества в экосистемах и невозможность восстановления коренной растительности в течение длительного времени являются движущей силой процессов автохтонного видообразования и дальних миграций видов и целых эколого-ценотических флористических комплексов (Галанин и др., 2008).
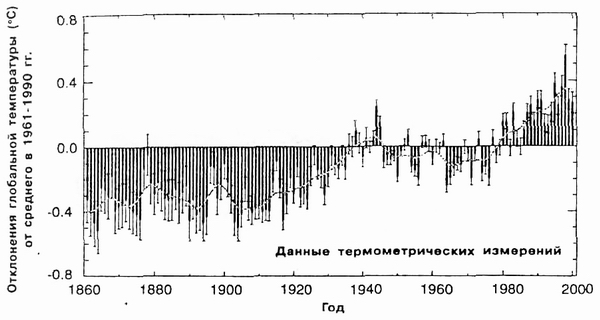
Рис. 2. Рост глобальной температуры приземного слоя воздуха с учетом территории суши и морской поверхности (по И. С. Касимову и Р. К. Клиге, 2006) |
На рис. 2 приведен график изменения одного из показателей климата - средней температуры января - за последние 100 лет. Хорошо видно, что в 1860 г. температура была -0,4°С, такой она продержалась до 1920 г., затем повысилась и в период с 1930 г. по 1980 г. колебалась около 0°, после 1980 г. температура начала снова повышаться и в 2000 г. достигла +0,4°С. Следовательно, за 140 лет температура приземного слоя воздуха на нашей планете увеличилась на 0,8 градуса. Некоторые ученые считают, что повышение температуры на Земле произошло из-за повышения в атмосфере концентрации углекислого газа, т.е. вследствие усиления парникового эффекта, суть которого в том, что световые солнечные лучи свободно проходят сквозь атмосферу, она для них прозрачна. А вот инфракрасные тепловые лучи, которые испускает нагретая солнцем поверхность, сквозь богатую углекислым газом атмосферу проходят с трудом, они отражаются назад, рассеиваются в атмосфере, дополнительно разогревая ее. Таким образом, заметное потепление на Земле началось в 1920 г.: это хорошо видно на рис. 2.
На основании длительных геоботанических и флористических исследований и анализа огромного фактического материала из разных регионов северного полушария (Галанин, 1991; Беликович, 2001; Галанин, Беликович, 2004; Флора ..., 2004) мы пришли к |
выводу о том, что практически во всех локальных фитоценозах имеется значительное таксономическое сверхразнообразие, когда из всего набора видов, встречающихся в данном фитоценозе, только треть принимают заметное участие в биологическом круговороте. Обилие остальных видов фитоценоза столь незначительное, что говорить о них как полноценных компонентах экосистемы не приходится. В связи с этим правомерен вопрос: почему эти виды вообще растут в этом фитоценозе? Ответ прост: в соседних фитоценозах эти виды обильны, производят много диазачатков, и эти диазачатки распространяются в пределах ландшафтного района широко, попадают в места на пределе толерантности этих видов, и здесь эти виды, хотя и растут, но находятся в угнетенном состоянии. Интенсивный семенной дождь - причина того, что некоторые виды растут в пределах ландшафтного района не только в оптимальных для них экосистемах, но и в не свойственной для них обстановке.

Фото 5. Массивы реликтовых сосновых (Pinus densiflora) лесов среди дубовых (Quercus mongolica) лесов на юге Приморского края в окрестностях с. Комиссаровка. Фото А.В. Галанина. |
Но проблема малообильных в фитоценозах видов этим не исчерпывается. В локальных флорах очень много видов, которые ни в одном конкретном фитоценозе данного района не являются обильными. Как попали в эту конкретную флору такие виды? Почему они не вытесняются из фитоценозов видами, которые находятся в этом районе в климатическом и эдафическом оптимуме? Почему в одних фитоценозах в пределах одного ландшафтного района редких и малообильных видов много, а в других мало? Почему в одних локальных флорах редких и малообильных видов больше, чем в других?
Обычно принято считать, что чем больше в сообществе редких и малообильных видов, тем оно устойчивее, тем больше его резистентность. Думаем, что это не столь однозначно. В том случае, когда климатические условия изменяются циклически, в одном сообществе могут сосуществовать виды разного экологического склада: мезофиты и ксерофиты в степях-прериях Даурии, мезофиты и криофиты в луговинных тундрах Южной Чукотки (Беликович, 2001), бореальные и неморальные виды в смешанных лесах Приморского края (Флора ..., 2004). Циклические изменения климата длительностью в несколько сотен лет приводят к тому, что для одной (холодной) стадии цикла оптимальны виды бореального склада, а для другой (теплой) стадии - неморальные виды. В течение климатического цикла более обильными становятся то неморальные виды, то бореальные. Такой бореально-неморальный маятник раскачивается в Приморье, по крайней мере, в течение нескольких тысячелетий. Аридно- |
гумидный маятник, характерный для растительного покрова Даурии, по нашим данным, раскачивается в течение нескольких десятков тысяч лет (Галанин, Беликович, 2004).

Фото 6. Лиственично-тополевый (Larix dahurica + Populus suaveolens) кустарниковый прирусловой лес в Алханайском национальном парке в Забайкальском крае. Фото Л.М. Долгалевой. |
Таким образом, значительное число видов разного (противоположного) эколого-ценотического склада, встречающихся в одном типе растительных сообществ в пределах площади одной конкретной флоры, говорит о том, что климатические условия в этом районе изменяются циклически,
что для растительного покрова района характерны изменения типа эколого-ценотического маятника. Нами установлено, что разные типы растительных сообществ в пределах площади одной конкретной флоры имеют разную степень устойчивости. При изменении климата один тип сообществ в ландшафтном районе (щебнистые тундры на крутых склонах гор на Чукотском полуострове) может измениться очень сильно, в то время
как кочкарные равнинные тундры в этом районе в это же время практически не изменятся. Например, мало изменяются при климатических пертурбациях сообщества верховых болот (виды доминанты в них сохраняются в течение 15 тыс. лет и более), в то время как сообщества на склонах холмов успевают за это время измениться практически полностью несколько раз (так, сосна многоцветковая на юге Приморья на склонах сменилась дубом монгольским, ель аянская на севере Приморья - кедром корейским).
Ареалогический анализ многих конкретных флор показал, что в одной конкретной флоре встречаются виды с разными типами географических ареалов. Так, например, во флоре Ононской Даурии на юге Читинской области встречаются виды с маньчжурским типом ареала, для которых долгота Хэнтей-Чикойского нагорья является западным пределом их ареалов, а также виды с центральноазиатским типом ареала, для которых долгота Большого Хингана - восточный предел их ареалов. О чем это говорит? Можно предположить, что в разные климатические эпохи на территорию Ононской Даурии |
осуществлялись миграции то с востока, то с запада и юго-запада. С востока в гумидные эпохи в Даурию расселялись маньчжурские виды, а с юго-запада в аридные эпохи «приходили» центральноазиатские и южносибирские виды. Судя по соотношению между восточным и западным ареалогическими элементами во флоре Ононской Даурии, можно заключить, что преобладали миграции видов с востока. Следовательно, Даурию следует рассматривать
 |
как продолжение Маньчжурии, а не Южной Сибири (Галанин, Беликович, 2006). Следует заметить, что в разных семействах во флоре Ононской Даурии соотношение видов маньчжурского и центральноазиатского склада существенно различается. Эти различия явно связаны с центрами происхождения разных семейств или же с центрами их видового разнообразия.
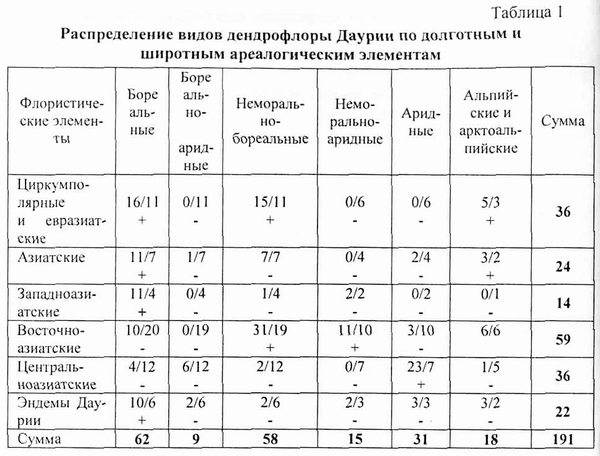
Значимые результаты дал сопряженный анализ структуры дендрофлоры Даурии (Галанин, Роенко, 2009) методом Пирсона (Галанин, 1987, 1991). На основании конспекта дендрофлоры было подсчитано количество видов и рас в каждом элементе пересечения широтных ареалогических групп с долготными ареалогическими группами и ареалогических групп с эколого-ценотическими группами (табл. 1 и 2). Рассчитаны суммы видов по широтным и долготным группам и суммы видов по ареалогическим и эколого-ценотическим группам Общее количество учтенных видов и рас 191. Рассматривая краевые суммы каждого столбца и каждой строки как независимые события, мы можем рассчитать частоту этих событий. Например, частота события «леса верхнего лесного пояса» может быть получена как дробь 25/191 (соответствующая краевая сумма, деленная на обшее количество учтенных таксонов). Эта частота измеряет вероятность попадания произвольного вида дендрофлоры Даурии в данный эколого-ценотический элемент. Частота события «восточноазиатские виды» равна Дроби 62/191, она измеряет вероятность попадания вида локальной флоры в данный ареалогический элемент. |
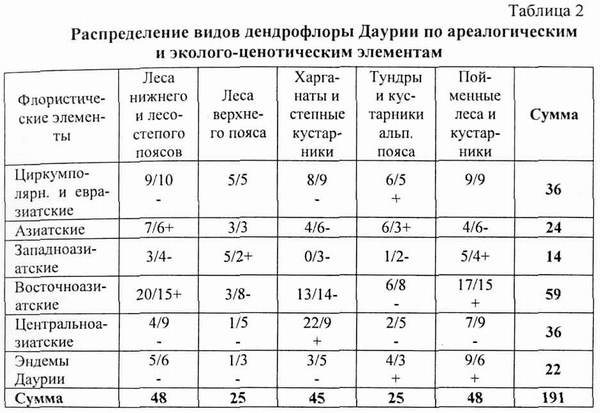 |
Перемножая частоты независимых событий в таблице, мы можем рассчитать ожидаемую частоту условного события. Например, частоту события «восточноазиатские виды при условии, что они окажутся и неморально-бореальными», можно рассчитать так: 62/191 х 57/191 = 19/191. Следовательно, в данном элементе пересечения при условии независимого (случайного) распределения таксонов дендрофлоры Даурии ожидается 19 видов. Сравнивая наблюдаемое количество видов в данном элементе пересечения (31) с ожидаемым (19), можно сказать, что в данном случае наблюдается положительная корреляция восточноазиатских видов с неморально-бореальными. При небольшом различии ожидаемого числа видов и наблюдаемого говорить о наличии достоверной корреляции нельзя. Однако, используя табличные значения критерия Пирсона хи-квадрат при двух степенях свободы, мы определили, что в нашем случае достоверными могут считаться отклонения не менее чем на 5 видов. Более подробно использование метод Пирсона для анализа структуры флор изложен в нашей публикации (Галанин, 1987 и на данном сайте).
Описанным выше способом были рассчитаны ожидаемые числа видов в каждом элементе пересечения (табл. 1 и 2), они показаны в знаменателе в каждой клетке обеих таблиц. В числителе указаны |
наблюдаемые числа видов. Сопряженный анализ позволяет выяснить, насколько и с каким знаком долготные ареалогические элементы дендрофлоры Даурии скоррелированы с широтными ареалогическими элементами или насколько долготные элементы той же флоры скоррелированы с эколого-ценотическими элементами. Из табл. 1. следует, что заладноазиатские виды положительно скоррелированы с бореальным флористическим элементом. Это значит, что с запада наша флора обогащалась бореальными видами, которые могли мигрировать в Даурию в холодные гумидные эпохи климатического цикла. Напротив, восточноазиатские виды положительно скоррелированы с бореалыю-неморальным элементом, что позволяет утверждать, что миграции видов с востока становились активными в теплые гумидные фазы климатического цикла. Центральноазиатские виды положительно скоррелированы с аридным элементом, что говорит о том, что миграции видов с юга активизировались в аридные фазы цикла (Walter et aL, 1983).

Фото 7. Закладка постоянной пробной площади для изучения динамики лиственичного (Larix dahurica) кустарникового леса в Алханайском национальном парке в Забайкальском крае. Фото Л.М. Долгалевой. |
Интересно то, что альпийский и арктоалышйский элементы в нашей флоре с долготными элементами скоррелирован весьма слабо. Здесь мы не видим существенного превышения наблюдаемых чисел видов над ожидаемыми ни в одном из элементов пересечения. Это позволяет утверждать, что дендрофлора альпийского пояса гор Даурии весьма устойчива в геологическом времени. Ее устойчивость можно связать с существенной изолированностью гольцов Хэнтея от гольцов северного Забайкалья, Восточного Саяна и Прибайкалья. Эта изолированность нарушалась только в эпохи очень сильных похолоданий так, что во второй половине плейстоцена дендрофлора гольцов Даурии развивалась автохтонно. Именно поэтому здесь так много эндэмичных подвидов и рас широко распространненных в Азии альпийских и арктоальпийских видов.
Восточноазиатские виды отрицательно скоррелированы с бореально-аридными и бореальными, что подтверждает сделанный выше вывод о климате, при котором в Даурию активизировались миграции видов из Восточной Азии. Циркумполярные и евразиатские виды в даурской дендрофлоре отрицательно скоррелированы с бореально-аридными и неморально-аридными. Это значит, что в аридные эпохи климатического цикла миграции видов в Даурию с севера и запада были резко ограничены. Центральноазиатские виды отрицательно скоррелированы с бореальными, неморально-бореальными, неморалыю-аридными и бореально-аридными. Из этого следует, что Центральная Азия очень давно не производила никаких видов, кроме аридных. Следовательно, аридизация климата Центральной Азии произошла еще в третичном периоде.
Сопряженный анализ ареалогической и эколого-ценотической структур (табл. 2.) показал, что эти структуры между собой скоррелированы в гораздо меньшей |
степени, чем долготная ареалогичсская структура с широтной ареалогической структурой. Вполне естественно, что центральноазиатские виды положительно скоррелированы с харганатами и степными кустарниковыми сообществами (сообщества ксерофитных местообитаний) и отрицательно - с лесами верхнего лесного пояса. Восточноазиатские виды положительно скоррелированы с лесами нижнего лесного и лесостепного поясов и отрицательно - с лесами верхнего лесного пояса. Циркумполярные и евразиатские виды достоверно не скоррелированы ни с одной из эколого-ценотических групп. Но вот азиатские виды недостоверно положительно скоррелированы с тундровыми сообществами альпийского пояса, западноазиатские - с лесными сообществами верхнего лесного пояса, а эндемы Даурии - положительно сопряжены с сообществами речных долин.

Даурская ботаническая экспедиция в Аргунской Даурии (Забайкальский край) в 2006г. Сбор гербария позволяет выявить таксономическое разнообразие растительного покрова. Фото В.А. Галанина. |
Сопряженный ареалогический и эколого-ценотический анализ дендрофлоры Даурии показывает, что она относится к восточноазиатским флорам. Эта дендрофлора возникла в результате постепенной трансформации флоры гумидного маньчжурского склада. Мы считаем, что эта трансформация длилась достаточно долго, настолько долго, что широколиственные маньчжурские леса успели превратиться в новый тип растительности - харганат (Беликович, Галанин, 2005). Миграции видов с запада, севера и с юга в становлении дендрофлоры Даурии играли весьма важную роль. Однако и автохтонное видообразование в становлении этой дсндрофлоры было весьма заметным. Из 191 таксона дендрофлоры Даурии 22 таксона или 11,5% - это узкие эндемы либо Даурии в целом, либо ее отдельных провинций. Мы считаем, что при более детальном изучении флоры Даурии количество эндемов, особенно в ранге подвидов, может увеличиться.
В заключение следует сказать, что феномен таксономического сверхразнообразия в растительном покрове требует дальнейшего осмысления и изучения. Связь таксономического сверхразнообразия фитоценозов с устойчивостью экологических систем не столь проста, как принято считать. Кроме таксономического разнообразия и разнообразия жизненных форм при анализе растительного покрова конкретных территорий, в том числе и конкретных фитоценозов, следует учитывать разнообразие эколого-ценотических и ареалогических флористических элементов. Состав флор зависит от того, насколько близок растительный покров района к климаксовому состоянию, имеются ли в растительном покрове района эколого-ценотические маятники, связанные с циклическими изменениями климата. Важнейшей экологической задачей является установление |
основных тенденций в динамике вековых смен растительности. Для этого необходимо изучить восстановительные и возрастные смены лесной растительности в связи с антропогенными воздействиями, установить закономерности и особенности формирования ценотической структуры в наиболее распространенных коренных и вторичных лесах, выявить основные закономерности распределения видов растений и фитоценозов в разных типах ландшафтов Дальнего Востока. Нам и нашим коллегам в последние 10 лет удалось проанализировать результаты мониторинга растительности в Хинганском заповеднике, Сихотэ-Алинском и Сохондинском биосферных заповедниках, создать системы постоянных геоботанических площадей для длительного мониторинга ценотической структуры лесной растительности Сихотэ-Алинского, Сохондинского заповедников, Алханайского национального парка, Онон-Бальджинского национального парка (Монголия) и юга острова Сахалин. В связи с изучением устойчивости экосистем важно установить степень соответствия ритмов сезонного развития аборигенных видов фанерофитов особенностям климатического ритма юга Приморского края.
Литература
Беликович А,В. Ландшафтная флористическая неоднородность растительного покрова (на примере модельных районов Северо-Востока России). - Владивосток: БПИ ДВО РАН, 2001. - 248 с.
Беликович А.В., Галанин А.В. Забайкальский харганат как тип растительности // Комаровские чтения. - Владивосток: Дальнаука, 2005. - Вып.52. - С. 98-126.
Василенко Н.А. Самоорганизация древесных ценозов. - Дальнаука,
2008. - 171с.
Галанин А.В. Ценотическая организация растительного покрова. -
Владивосток: ДВО АН СССР, 1989. - 164 с.
Галанин А.В., Беликович А.В. Даурия как подобласть Дауро-Маньчжурской ботанико-географической области // Комаровские чтения.
- Владивосток: Дальнаука, 2006. - Вып. 53. - С. 9-31.
Галанин А.В. Опыт сопряженного анализа типологических структур конкретных флор // Теоретические и методические проблемы сравнительной флористики. - Л.: Наука, 1987. - С. 23-56.
Галанин А.В. Флора и ландшафтно-экологическая структура растительного покрова. - Владивосток: ДВО АН СССР, 1991. - 272 с.
Галанин А.В., Беликович А.В. Постоянные геоботанические пробные площади Сохондинского биосферного заповедника. - Чита: Поиск,
2004. - 228 с.
Галанин А.В., Роенко Е.Н. Конспект деидрофлоры Даурии // Бюлл.
БСИ ДВО РАН [Электронный ресурс] : науч. журн. / Ботан. сад-институт
ДВО РАН. - Владивосток, 2009. - Вып. 3. - С. 5-23. - http://botsad.ru/journal/ number3.htm
Галанин А.В., Беликович А.В., Галанина И.А., Роенко Е.Н., Яковченко Л. С. Растительность Даурии и великие переселения народов //
Вестник СВНЦ ДВО РАН, 2008. - № 4. - С. 41-53.
Ипатов B . C . Дифференциация древостоя II // Вести. ЛГУ. Биология.
1969. - Вып. З. - С. 43-54.
Касимов Н.С., Клиге Р.К. Введение. Проблемы глобальных изменений // Современные глобальные изменения природной среды, Т. 1. - М:
Научный мир, 2006. - С. 19-34.
Флора Сихотэ-Алинского биосферного заповедника (сосудистые
растения). - Владивосток: БСИ ДВО РАН, 2004. - 301 с.
Krupa S. Global climate change: Processes and products - fh overview //
Environ. Monit. Assess. 1997. V. 46. № 1-2. P 73-88.
Walter H., Box E. O., Hilbig W. The deserts of Central Asia // Ecosystems of the world 5: Temperate deserts and semi-deserts. N. E. West (ed.). New
York: Elsevier Scientific Publishing Company, 1983. P. 193-235.
|


