УДК 581.41
Базальные и медиальные девиации в эволюции жизненных форм высших растений
А.П. Хохряков, М.Т. Мазуренко
© OCR - М.Т. Мазуренко, 2010. Воспроизводится по тексту: А.П. Хохряков, М.Т. Мазуренко. Базальные и медиальные девиации в эволюции жизненных форм высших растений // Журнал общей биологии, 1991. T. 52, N 1.
До настоящего времени примеры модусов филэмбриогенезов у растений не касались эволюции их онтогенеза в целом, а лишь морфогенеза отдельных органов. На примере эволюции жизненных форм растений можно выявить их модусы эволюции, так как они проходят стадии онтогенеза целиком. Основным модусом эволюции онтогенезов жизненных форм растений являются анаболии (надставки) в сочетании с базальной аббревиацией, т.е. ускорением прохождения первых стадий. Материал по девиациям приводится главным образом на примере эволюции стланиковых жизненных форм. Начинаясь в качестве факультативных, отклонения от типичного хода онтогенеза с помощью стабилизирующего отбора могут в конце концов закрепиться наследственно, что приводит к выработке новых облигатных жизненных форм и новых таксонов разного ранга. Возможен также комбинированный (девиации-анаболии) ход эволюции онтогенезов.
Введение и постановка вопроса
Базальные и медиальные девиации – так называемые модусы морфологической эволюции. Проблемы связанные с этим кругом вопросов уже долгое время не перестают волновать биологов, но к сожалению, как правило лишь зоологов. Ботаники уделяют им очень мало внимания. Лишь А.Л. Тахтаджян в свое время пытался применить подходы «теории филэмбриогенеза» А.Н. Северцова к растительному миру (Тахтаджян,1943, 1964) Однако, как показала М.М. Лодкина (1983) при этом им совершенно недостаточно была учтена специфика растительных организмов сравнительно с животными, их высокая степень метамерии, органообразование в течение почти всей жизни, высокая модификационная способность, гораздо меньшая интегрированность разных частей и т.д. В результате приводимые А.Л. Тахтаджяном примеры тех или иных модусов (в том числе считавшегося основным у растений–неотении) оказались присущими не столько онтогенезу растений в целом, сколько отдельным органам, многократно образующимся в течение их жизни. Сюда относятся явления рудиментации венчика, андроцея или гинецея, листьев, корней и других органов метамеров, зигоморфия цветка, ассиметрия листьев и другие девиации ранних и поздних стадий морфогенеза (но не онто-).
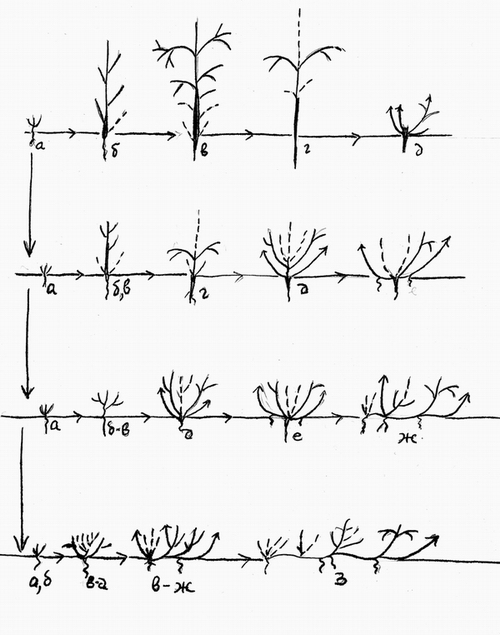
Рис.1. Эволюция жизненных форм растений путем анаболий (надставок). Онтогенезы.1-дерева, 2- аэроксильного кустарника. 3- геоксильного кустарника или кустарничка, 4- кустарничка, полукустарничка или вегетативно-подвижного травянистого растения. Штрихом обозначены отмирающие ветви, со стрелками – побеги формирования. Тонкие горизонтальные стрелки – направления онтогенеза, направленные вниз толстые – направления филогенеза, маленькие русские буквы – стадии онтогенеза. |
Особенно в этом отношении показательны метаморфозы листьев в филлодии у акаций и серии переходов между лепестками и тычинками у кувшинок. Трактуемые обычно как девиации средних стадий онтогенеза серии метаморфозов обычных листьев в филлодии у некоторых африканских и австралийских акаций могут повторяться в кроне дерева неоднократно, часто на каждой новой серии побегов заново, а иногда и в обратной последовательности (от филлодиев к нормальным листьям). Это явление, а также переходы между лепестками и тычинками у кувшинок) объясняются М.М. Лодкиной тем, что «в онтогенезе современного растения» одновременно присутствуют девиации разных стадий морфогенеза органа. Полностью соглашаясь с этим, не можем не отметить, что и М.М. Лодкиной не приведены примеры каких-либо модусов филэмбриогенеза у растений как целостных организмов. Считаем, что в качестве таковых могут служить и жизненные формы (ЖФ), в понимании И.Г. Серебрякова (1963). В частности у высших растений их ЖФ – собственно тело растения, совокупность их побеговых (предпобеговых) теломных и корневых систем.
В свое время один из авторов данной работы (Хохряков, 1973, 1975а, б) основным способом эволюции онтогенезов ЖФ растений считал надставки (анаболии по А.Н. Северцову) в сочетании с базальной аббревиацией. Мы не будем подробно останавливаться на этом вопросе, отсылая читателя к указанным источникам. Упомянем лишь, что по нашему мнению, путем надставок шла эволюция таких жизненных форм семенных растений, как аэроксильных кустарников (от деревьев), полукустарников, лиан (от кустарников), кустарничков (от кустарников). Корневищных трав (от стержнекорневых трав и от геоксильных кустарников). Геоксильных кустарников (от аэроксильных кустарников) . Эволюция путем анаболий изображена на рис.1.
Даже травянистые растения, по нашему мнению и вопреки мнению А.Л. Тахтаджяна, произошли от деревянистых и именно путем анаболий – все более раннего и более интенсивного развития, вначале у деревьев, а затем у кустарников, деревец и полукустарников – «отравяненых» водяных побегов (побегов формирования), а не путем неотении, как это сейчас общепризнано. Путем надставок (в сочетании с базальной аббревиацией. Т.е. ускорением развития первых фаз онтогенеза) шло и формирование основных ЖФ водорослей нитчатой на базе одноклеточной, разнонитчатой на базе гомотрихальной, пластинчатой на базе нитчатой.
|
Создавалось впечатление, что все основные ЖФ растений обязаны своему возникновению анаболиям и пролонгациям конечных стадий. Однако постепенно стал накапливаться и иной материал. Работая над проблемой лесных стлаников и стланичков М.Т. Мазуренко (1978) обратила внимание на то, что многие обычные лесные кустарники, попадая в условия сильного затенения уже с самого начала онтогенеза сильно отклоняются от обычного хода онтогенеза и вместо того чтобы расти «нормально», превращаются в стланики и стланички. Отклонения ранних стадий онтогенеза древесных растений описаны как у кустарников, так и деревьев (см. Белостоков, 1983), однако с модусами филэмбриогенеза их не связывали. Мы в данной работе поставили перед собой именно эту цель. Однако прежде познакомим читателя с предложенной нами периодизацией онтогенеза древесных растений.
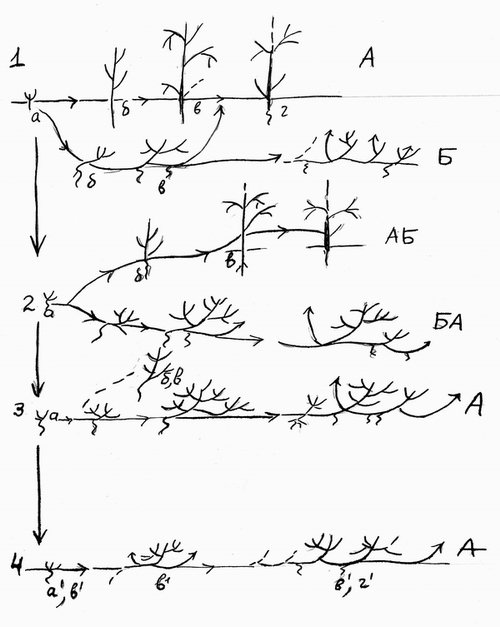
Рис.2 Эволюция жизненных форм растений путем девиаций. Онтогенезы: 1 дерева и факультативного стланца. 2 – дерева (деревца) и полуоблигатного стланика. 3 – облигатного стланика и факультативного деревца, 4 – облигатного стланичка и кустарничка, А – прямой путь онтогенеза, Б-уклоняющийся. Штриховые стрелки – вероятные пути онтогенеза. |
Периодизация онтогенеза древесных растений
Одной из отличительных черт растительного организма в сравнении с животными, не отмеченной М.М. Лодкиной является то, что растения не только всю жизнь растут, но всю жизнь, образно выражаясь, умирают. В этом, вероятно, выражается колониальная природа растительного организма, подмеченная еще Э. Геккелем. Чуть ли не с самого прорастания подавляющее большинство растений «сбрасывает» или меняет те или иные части, органы, вначале – семядоли и первичные корешки, затем – нижние листья и затем – слабые побеги. Разумеется, поскольку растение не отмирает полностью, а продолжает расти процессы отмирания вначале сильно отстают от процессов роста, но с течением времени (неоднократно у разных видов и в разных условиях) сначала наступает равновесие между ростовыми и деструктивными процессами, а затем эти последние начинают преобладать (теоретически), что и приводит к смерти (далеко однако не всегда – особенность растений в сравнении с животными).
На основании изложенного мы (Мазуренко,1980,1986) делим онтогенез высших растений на три основных периода. 1) с преобладанием ростовых процессов (период «рост»), 2) со сбалансированностью процессов роста и отмирания (период «стабилизация»). 3) с преобладанием деструктивных процессов (период "отмирание").
Наряду с тремя периодами онтогенеза высшего растения подразделяем и на фазы (стадии) отличающиеся друг от друга морфологическими (по строениюЖФ) и физиологическими признаками. У других авторов они носят название возрастных состояний (Смирнова и др. 1976) и теоретически различаются способностью к продуцированию генеративных органов. Вопрос о соотношении фаз и возрастных состояний разобран ранее ( Мазуренко,1980). Подчеркнем принципиальную разницу между периодами и фазами: первые определяются соотношением процессов роста-отмирания, вторые морфологическими признаками ЖФ. Вследствие этого как имеющие биоморфологическую специфику разные стадии онтогенеза растений именуются нами онтобиоморфами (Мазуренко,1986). |
|
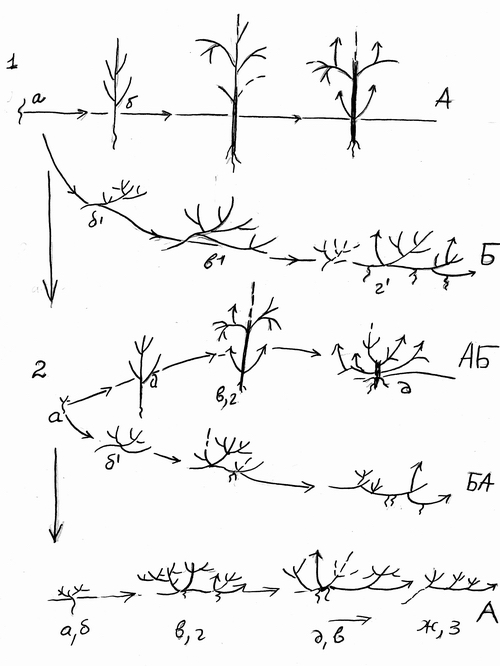
Рис.3. Эволюция жизненных форм растений путем комбинированного действия анаболий и девиаций. Онтогенезы: 1 – дерева и факультативного стланца. 2. – деревца (аэроксильного кустарника) и стланика (геоксильного кустарника) 3– кустарничка или облигатного лесного стланичка |
Первый период онтогенеза самый ответственный. В это время неокрепший молодой организм подвергается наибольшему давлению со стороны среды, абиотически ее факторов и биотических (конкуренция). Следовательно молодое растение в этот период наиболее уязвимо и подвержено модификационному влиянию среды, хотя интенсивно протекающие ростовые процессы всеми силами стараются им противостоять. Отсюда так называемая поливариантность онтогенеза, столь характерная для растений в отличие от животных.
Во втором периоде онтогенеза растение находится в расцвете сил. В условиях экологического оптимума оно достигает наибольших определяемых его генотипом размеров, наибольшей биомассы и лучшим образом противостоит всем изменениям среды обитания, следовательно проявляет минимум модификационных возможностей. Те отклонения которые растение, возможно, испытало в первом периоде, во втором «приходят в норму». Организм как бы возвращается на свою «прямую онтогенетическую траекторию» (Белоусов, 1979). В этом периоде, следовательно, в наибольшей степени проявляется гомеостаз (гомеорез) онтогенеза (Мазуренко, 1987). На наш взгляд ожидать каких либо эволюционных преобразований, которые бы начинались в этом периоде, можно с наименьшей долей вероятности.
В третьем периоде происходит заметное и все более прогрессирующее ослабление функций роста. Усиливающиеся процессы деструкции приводят ко все более прогрессирующему ослаблению организма, который снова может стать добычей разнообразных отклонений (девиаций) и уклонов, правых и левых. У животных это редко приводит к иному результату, чем смерть без всяких эволюционных последствий, тем более что животные в этом периоде, как правило теряют свою генеративную функцию. Растения же продолжают сопротивляться смерти до конца, и даже уже обреченное растение производит в некотором количестве и молодые побеги, и генеративные органы. Это и служит основой их геронтоморфоза, т.е. эволюции путем возникновения и прогрессирования надставочных фаз развития. Таким образом девиации у растений в эволюции из ЖФ всего более вероятны именно как базальные, как отклонения от онтогенетической прямой на ранних стадиях онтогенеза, в первом его периоде и на первых стадиях (онтобиоморфах).
|
Возникновение и эволюция лесных и иных стлаников
Множество древесных и кустарниковых пород как в Средней России так и по всей лесной зоне (Дальний Восток, Кавказ, Средняя Азия) в условиях глубокого затенения образуют стланиковые и кустарниковые биоморфы. Особенно это характерно для липы (Tilia cordata L.) Подлесок из кустарниковой формы липы – обычное явление во всех широколиственных и смешанных лесах. Эта форма образуется уже на первых этапах онтогенеза в первом его периоде так как онтобиоморф второго периода при этом обнаружить, как правило не удается. Однако кустарниковая форма липы почти не цветет из-за сильного угнетения и потому не может явиться базой для возникновения облигатной стланиковой формы липы.
Большинство лесных кустарников (калины, жимолости, смородины, бересклеты) наряду с прямостоячей формой роста образуют и стланиковую, названную нами биоморфой «лесного стланика» (Мазуренко, 1978) Это факультативные ЖФ подобны и кустарниковой форме липы. Некоторые из них, например жимолость съедобная (Lonicera edulis Turcz. ex Frein) на Дальнем Востоке, бересклет Купмана (Evonimus coopmanii Lauche) в Средней Азии бересклет низкий (E. nana Bieb.) имеют как бы параллельно существующие две почти равноценные биоморфы развивающиеся одинаково хорошо как на более освещенных местах в качестве прямостоячего кустарника или кустарничка, так и в условиях более сильного затенения как лесные стланички. И, наконец, имеется достаточно много облигатных лесных стлаников и стланичков не развивающих прямостоячих биоморф ни в каких благоприятных условиях, как например широко распространенная во всей таежной бореальной полосе линнея северная (Linnea borealis L.), барвинок малый (Vinca minor L.) в широколиственных лесах юго-восточной Европы падуб, морщинистый (Ilex rugosa Fr. Schmidt) в темнохвойных лесах Сахалина, гаультерия Шалон (Gaulteria shalon Pursh) в темнохвойных лесах Орегона, ардизия зубчатая (Ardisia crenata Sims.) в горных лесах юго-восточной Азии.
|

Ольха кустарниковая (Alnaster fruticosa) древовидной формы на севере Корякского нагорья в закрытом от ветров распадке.
Фото А.В. Галанина |

Ольха кустарниковая (Alnaster fruticosa) стланиковой формы в виде кустарника (на заднем плане) и в виде шпалерного кустарничка (на переднем плане). Север Корякского нагорья, берег бухты Угольная.
Фото А.В. Галанина
Форма кустов ольховника зависит от мощности снегового укрытия зимой. Там, куда снег надувается, кусты ольхи более высокие, а там, где сдувается - низкие. Древовидная форма и форма в виде шпалерного кустарничка у ольховника на Чукотке и в Корякии встречаются очень редко. |
Какова генетическая сторона биоморфологической эволюции в этих случаях, мы в точности не знаем. Вероятно, облигатные лесные стланички – результат стабилизирующего отбора, который допускает некоторое время параллельное существование двух аллеломорф, кодирующих два варианта биоморф: первоначальную – прямостоячего растения и новую – стланиковую.
Как и в лесах, все стадии перехода от облигатных прямостоячих растений к облигатным стланиковым можно наблюдать и на верхнем пределе лесов, и при переходе от таежной зоны к арктической. Разница лишь та, что при формировании ЖФ лесного стланика ведущим фактором выступает затенение, а здесь – похолодание и наличие снегового покрова. Рассмотрим образование субальпийских и полярных стлаников на некоторых примерах.
|

Дуб монгольский (Quercus mongolica) на западном пределе географического распространения часто имеет форму кустарника. Фото Е.Э Малкова. |

Пихта сибирская (Abies sibirica) на верхнем пределе распространения на горе Сохондо (южное Забайкалье) нередко образует форму стланника. Растут нижние ветви, которые укрываются снегом, а все, что выше снежного укрытия, погибает от холода. Фото Л.М. Долгалевой |
В Японии, на Сихотэ-Алине и Южных Курильских островах обитает один из видов рододендронов – рододендрон Фори (Rhododendron fauriei Franch) который обычно имеет форму небольшого деревца до 6-7 м. Однако на крутых склонах он начинает стелиться вниз по склону, приобретая стланиковую ЖФ. Правда, как только этот стланик «выползает» на ровное открытое пространство, он дает прямостоячую ветвь, превращающуюся затем в прямостоячий ствол,т.е. возвращается к прежней исходной ЖФ (Мазуренко, 1980). Стланиковая форма для этого рододендрона – факультативная, хотя она цветет и плодоносит.
У кавказского рододендрона понтийского (Rhododendron ponticum L.) соотношение биоморф обратное. Только в очень благоприятных условиях (при посеве вне конкуренции с другими растениями и без повреждений морозами) он развивается первое время как небольшое деревцо 3-5 м высотой. Обычно же это гигантский стланик достигающий в нижнем горном поясе 5-6 м высоты. На верхней границе леса он становится гораздо ниже –1,5-2 м.
Две формы роста – кустарниково-стланиковую и древесную образуют и некоторые другие породы близ верхней границы леса на Кавказе: береза Медведева (Betula medwedevii Regel), клен Траутветтера (Acer trautvetteri Medw.) бук восточный (Fagus orientalis Lipsky) и др. Нет сомнений, что кустовидные и стланиковые их формы – производные древовидной прямостоячей поскольку эта последняя более отвечает оптимуму существования вида. Кустовидно-стланиковая форма у них – отклонение от обычного хода онтогенеза в более экстремальных условиях. В то же время в субальпах Кавказа близ верхней границы леса существуют облигатные кустовидно-стланиковые древесные породы, такие как понтийский дуб (Quercus pontica Koch). В настоящее время мы не находим его близких древовидных форм, однако нет сомнений, что именно таковые формы являлись его предками, поскольку именно они населяют лесной пояс Кавказа, и кустарниково-стланиковые формы – производные от древовидных (Серебряков, 1963). Подобный вывод никем не оспаривается. Поэтому важно подчеркнуть, что кустарниково-стланиковые формы субальпийских криволесий – результат девиаций предковых форм. Таково же происхождение по всей видимости стланиковых сосен (Pinus mugo Turra – на западе и P. pumila Pall. – на востоке). Этот последний вид еще не потерял способности к возвращению к исходной древовидной форме в особо благоприятных условиях. Схема эволюции жизненных форм путем девиации дана на рис. 2.
Девиации и таксономия
Понтийский дуб – вид того же самого рода Quercus L. что и его вероятные предковые формы. Но в природе существуют и рода, которые можно себе мыслить как кустарниково-стланиковые производные первичных древовидных форм. Таковы например Alnaster Spach (= Duscheckia Opiz) – ольховники кустарники и стланики, производные от древовидных видов ольхи, свойственные субальпийским криволесьям Европы и Дальнего Востока (а Alnaster fruticosa Rupr. Занимает и огромные площади в Субарктике), небольшой реликтовый и эндемичный для Сихотэ-Алиня род Microbiota Kom. – явное производное рода Biota L., состоящего из больших и маленьких деревец, линнея – небольшой род, явное производное кустарниковых абелий.
|

Лиственница даурская (Larix dahurica) на вершине горы Алханай в южном Забайкалье нередко принимает стланниковую форму, - развиваются нижние ветви, которые зимой укрываются снегом, а все, что выше снегового укрытия, гибнет от мороза. Фото Л.М. Долгалевой |

Ива изменчивая (Salix divaricata) на горе Сохондо в южном Забайкалье в нормальных условиях растет в форме кустарника высотой до 1-1,5 м , но на вершине на высоте 2500 м н.у.м. это корявый низкий кустарничек, стелющийся по субстрату. Фото А.В. Галанина
Таким образом, девиации (по видимому и анаболии) начинающиеся с отклонения от прямого хода онтогенеза лишь побеговых систем ЖФ, с параллельного существования двух (вероятно и более вариантов онтогенеза ЖФ, заканчиваются формированием хороших таксонов вначале ранга вида, а затем и более высокого ранга (Мазуренко, Хохряков,1981). Так, если выделять в особый род Terrorodion (Maxim.) Small. – рододендроны камчатский и Редовского, следует иметь ввиду что отличия их от рода рододендрон в основном биоморфологического порядка (стланиковый рост, способность развивать подземные корневища, кистевидные малоцветковые соцветия) которые возникли, по всей видимости в процессе приспособления к субарктике первоначально кустовидных форм в качестве девиации. |
Связь родов Arbutus L., Arctostaphylos Adams и Arctous (A. Grey) Niedenzu не такая тесная, как предыдущей пары, однако тесное их родство неоспоримо (они относятся к одному подсемейству и всегда ставятся рядом. Отличаются они между собой главным образом биоморфологическими признаками: первая пара – формой роста (Arbutus – деревья и деревца, часто кустовидные или крупные кустарники. Arctostaphylos – кустарники, в основном стелющиеся или типичные стланики и стланички, (как например широко распространенная толокнянка – A . uva-ursi L.). Вторая пара – в основном листопадностью (Arctous – листопадные кустарнички).
Правда, с помощью какого модуса – девиации или анаболии – произошли Arctous от Arctostaphylos, не совсем ясно, так как листопадность у вечнозеленых растений усиливается при всяком ослаблении, как в начале онтогенеза, так и в конце. Точно также обстоит дело и с кущением. Как мы выяснили (Мазуренко, 1980) у Rhododendron mucronulatum Turcz. На юге Приморского края в течение онтогенеза наблюдается две вспышки кущения, т.е. образования мощных побегов формирования от основания куста: первая – в первом периоде онтогенеза, когда растение испытывает угнетение со стороны окружающей биоты, вторая – в оптимальных условиях широколиственных лесов. Эта фаза – факультативная, и во втором периоде растение представляет собой деревце. Но в третьем периоде (пятая фаза) кущение вновь усиливается и растение становится крупным кустом.
Явно производные от рододендрона остролистного рододендроны даурский (Rhododendron dauricum L.) и мелколистный (Rhododendron parvifolium Adam) уже очень редко (первый) или никогда (второй) не развивают деревцевидной формы (точнее – не возвращаются к ней) и, таким образом возникновение ЖФ куста у них можно мыслить себе двояким образом: как результат девиации (отклонения на ранней стадии) или как результат анаболии (добавление в конце онтогенеза). Вероятен еще один комбинированный путь как девиация и как анаболия начавшись с первых стадиий онтогенеза как девиация длительно осуществляемое кущение сталкивается с тем кущением, которое шло от конечной фазы онтогенеза. Ведь путь анаболии также предполагает вначале факультативность, а затем и облигатность надставочных стадий, которые становятся все более продолжительными. Комбинированный путь эволюции изображен на рис.3.
Можно последний пример считать и отклонением всего хода онтогенеза (тотальной девиацией). Следует лишь обратить внимание, что у растений он отнюдь не обязательно начинается с первых стадий (не является архаллаксисом по А.Н.Северцову), так как не связан ни с какими изменениями эмбриональных структур. В этом проявляется еще одна особенность растительных организмов в сравнении с животными. Эволюционные изменения у растений эмбриональных структур, как например у паразитных, не являются результатом их автономной эволюции (Терехин, 1977) а есть результат девиации или анаболии, ставший тотальным в ходе дальнейшей эволюции (Хохряков, 1981).
Вполне вероятно, что такова же природа тех изменений зародыша, которые имели место при становлении типа однодольного растения от типа двудольного, что и привело к мысли о неотении как о модусе их эволюционных преобразований (Тахтаджян, 1943, 1964). Однако возможно однодольные происходят непосредственно от голосеменных предков, у которых однодольность уже была развита. Однако эта проблема – не предмет настоящего исследования.
Заключение
Онтогенез растений весьма существенно отличается от онтогенеза животных постоянным возникновением почти в течение всей жизни все новых и новых серий метамеров и их систем, благодаря чему морфогенез этих органов, как правило, к онтогенезу организма не имеет такого прямого отношения, как у животных. Тем не менее модусы эволюции онтогенеза растений могут быть те же что и у животных, по крайней мере частично, а именно девиации и анаболии. Искать эти модусы нужно в эволюционных преобразованиях жизненных форм растений. Основным модусом онтогенеза у растений являются анаболии; путем девиации эволюционировали главным образом стланиковые и стланичковые жизненные формы. Возникнув на средних или поздних стадиях онтогенеза девиации у растений имеют тенденцию «спускаться» к его началу, захватывая в конце концов и эмбриональные стадии. Так может создаться иллюзия существования у растений неотении. Преобразование онтогенетической траектории сопровождается и таксономическими изменениями разного ранга.
Список литературы
Белостоков Г.П. Системы стеблесложения подроста древесных растений // Бюлл. МОИП Отд.биол. 1983. Т. 88. №1. С.87-104.
Белоусов Л.В. Целостные и структуро-динамические подходы к онтогенезу // Журн. Общей биологии 1979 Т. 40 №4 С. 514-521.
Лодкина М.М. Черты морфологической эволюции растений, обусловленные спецификой их онтогенеза // Журн. Общей биологии 1983. Т. 44 №2. С. 239-253.
Мазуренко М.Т. О жизненной форме стелющихся лесных растений // Ботан.журн.1978. Т. 63. №4. С. 593-603.
Мазуренко М.Т. Рододендроны Дальнего Востока:структура и морфогенез. М: Наука, 1980. 231 с.
Мазуренко М.Т. Биоморфологические адаптации растений Крайнего севера. М.: Наука, 1986, 208 с.
Мазуренко М.Т. О гомеостазе онтогенеза древесных растений // Материалы 5 Всесоюзной школы по теорет. Морфологии растений. Львов. Инстит. ботаники АН УССР, 1987. С. 65-69.
Мазуренко М.Т., Хохряков А.П. Биоморфологическая изменчивость и ее связь с таксонообразованием у растений (на примере семейства вересковых) // Жизненные формы: структура, спектры и эволюция. Н.: Наука. 1981. С. 12-30.
Серебряков И.Г. Экологическая морфология растений. М.: Высш.шк. 1963. 378 с.
Тахтаджян А.Л. Соотношение онтогенеза и филогенеза у высших растений. Этюды по эволюционной морфологии // Тр.Ереванского университета. 1943. Т. 22. С. 71.
Тахтаджян А.Л Основы эволюционной морфологии покрытосеменных. Л.: Изд-во АН СССР. 1964. 236 с.
Терехин Э.С. Паразитные цветковые растения. Л.: Наука. 1977. 219 с.
Хохряков А.П. Основные закономерности эволюции онтогенеза у растений // Бюл МОИП Отд. биол. 1973. Т. 78. Вып.4. С. 37-49.
Хохряков А.П. Закономерности эволюции растений. Новосибирск: Наука, 1975а. 202 с.
Хохряков А.П. Соматическая эволюция однодольных М.: Наука.1975б. 195 с.
Хохряков А.П. Эволюция биоморф растений. М.: Наука.1981. 169с.
Смирнова О.В. Заугольнова Л.В.,Торопова Н.А. Фаликов Л.Л. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф // Ценопопуляции растений: основные понятия и структура Ч.1 М.: Наука.1976. С.14-44.

