 |
Анализ графов флористической структуры растительного покрова
двух ландшафтных районов Чукотки А. В. Галанин © 2010
© OCR - А.В. Галанин, 2010. Воспроизводится по тексту: Галанин А.В. Анализ графов флористической структуры
растительного покрова двух ландшафтных районов Чукотки // Метод графов в экологии. Владивосток: ДВО АН СССР, 1989. С.6-24.
Эта статья мало известна геоботаникам, и новый метод анализа структуры растительного покрова на ландшафтном уровне организации с помощью
графовых моделей, которому и посвящена статья, до сих пор не используется исследователями. Метод позволяет ответить на вопрос, насколько
структурирован растительный покров ландшафта, насколько его структура определяется внешними факторами, а насколько внутренними ценотическими.
Сформулировано представление о равномерности - неравномерности структуры, предложен алгоритм для расчета графа с стохастическим распределением
ребер по вершинам. С той поры метод был усовершенствован нами, создана программы для ПК, позволяющая строить графовые модели при разных пороговых
уровнях сходства вершин и сравнивать полученные графы с их стохастическими моделями при том же количестве вершин и ребер. |
Введение
Под флористичсской структурой растительного покрова мы понимаем систему отношений сходства-различия, выявленную на множестве типов фитоценозов (групп ассоциаций) в пределах одного элементарного ландшафтного района, когда оценивается степень общности видового состава этих фитоценозов. Флористическая структура растительного покрова в пределах района с одной конкретной флорой в некоторой степени соответствует его экологической структуре в понимании В. Б. Сочавы (1978).
В процессе многолетних исследований методом конкретных флор нами изучался растительный покров разных регионов Советского Союза: среднее течение р, Эргувеем и окрестности Чаунского стационара на Чукотке (1971— 1974 гг.), нижнее течение р. Преголи в Калининградской области (1973—1975 гг.), среднее течение р. Вычегды в Коми АССР (1983 – 1984 гг.) В каждом случае тщательно исследовалась, конкретная флора высших сосудистых растений (Галанин, 1987), а для выявления разнообразия растительности в каждом районе составлялось 200 - 300 геоботанических описаний на пробных площадках размером 100 кв. м для травянистой и кустарничковой растительности и 400 кв. м для древесной и кустарниковой растительности. Пробные площади рассредоточивались в пределах площади конкретной флоры (зона радиусом около 10 км) так, чтобы максимально полно охватить основное ландшафтно-экологическое разнообразие в 3-4-кратной повторности. Затем все описания группировались в типы местообитаний или группы ассоциаций. В одну группу ассоциаций (тип местообитаний) объединялись фитоценозы, сходные по положению в рельефе, характеру и интенсивности увлажнения, характеру почвы и подстилающей горной породы, набору обильных видов и общему флористическому составу.
Для каждой группы ассоциаций составлялся полный список встреченных видов высших сосудистых растений. Эти списки использовался при составлении сводных таблиц или матриц распределения видов по группам ассоциаций. Группы ассоциаций в пределах каждого ландшафтного района сравнивались попарно для оценки их флористического сходства по Жаккару. Результаты сравнения записывались в виде матрицы попарного (бинарного) сходства групп ассоциаций.
Ординация и дальнейший анализ этих матриц выполнялись графически. Группы ассоциаций (типы местообитаний) изображались кружками с соответствующими номерами. Сходные типы располагались по возможности ближе друг к другу и соединялись линиями, толщина и характер которых обозначали уровень флористического сходства между соединенными типами местообитаний. Графы изображались на плоскости, поэтому передать их многомерную структуру без искажения было невозможно. Неискаженным, однако, остается основное свойство графа - распределение множества ребер на множестве вершин. Именно это свойство и следует использовать в дальнейшем анализе.
На рис. 1 и 3 изображены графы флористического сходства типов местообитаний двух ландшафтных районов Чукотки. Характеристика типов местообитаний дана в соответствующих разделах. Более полную характеристику можно найти в более ранних публикациях (Галин, 1977, 1980, 1987).
Мы убеждены в том, что метод' графов - это рациональный способ отображения и анализа структуры сложных объектов и явлений, каким является растительный покров ландшафтного района. С помощью графов растительный покров можно представить как некий континуальный процесс, совершающийся в экологическом пространстве. К сожалению, представления о пространственных процессах, которыми являются континуальные структуры, развиваются чрезвычайно медленно. Попрежнему как в геоботанике, так и в зоогеографии преобладают попытки выявить и проанализировать закономерности организации растительного покрова и животного населении путем типизации и классификации отдельных пространственных состояний объекта. Конечно, традиционный способ анализа пространственных процессов дает немало; но все же многое при этом остается как бы «за кадром», в частности, такой вопрос, как соотношение дискретности и непрерывности структуры. Мы мало задумываемся о том, может ли континуальная структура быть иерархичной или это свойство лишь дискретных структур. Открытым, на мой взгляд, остается и вопрос о том , исчерпывается ли разнообразие всех пространственных процессов дискретными и континуальными типами структур, и различными их сочетаниями. Подобные вопросы не являются схоластическими, как может показаться на первый взгляд. Было бы чрезвычайно интересно выяснить, какая структура — дискретная или континуальная — экологически более оптимальна в разных условиях.
При изучении ландшафтно-экологической структуры растительного покрова, а также, повидимому, и животного населения, и всей бноты в целом непрерывность их хорологической и типологической структуры следует рассматривать не как помеху, затрудняющую выделение типов или классов состояний пространственного процесса, а как важнейшее свойство этого процесса, имеющее фундаментальное экологическое значение, К сожалению, арсенал методов работы с континуальными структурами сегодня невелик и сводится в основном к нахождению числа осей, пространства ординации и определению разнообразия состояний в направлении этих осей.
В настоящей статье с помощью вероятностных методов. мы попытались проанализировать некоторые, свойства графов как пространственных процессов. По аналогии с процессами во времени пространственные процессы также могут быть равномерными и неравномерными, непрерывными и дискретными, циклическими и линейными. Одним словом, особенности структуры графов нельзя описать лишь разными сочетаниями дискретности и непрерывности.
Методика
Пусть некий граф G имеет n вершин (х1, х2, х3, ..., хn). Вершины пронумерованы, то есть мы имеем дело с помеченным графом. На множестве вершин n распределено множество ребер m (у1, y2, y3, ..., ym). Структура графа определяется распределением ребер по вершинам, следовательно, характеристика структуры графа должна быть характеристикой этого распределения.

Следовательно, в равномерно связном графе с п вершинами и m ребрами из каждой вершины должно исходить по 2m/n ребер. В этом случае мы можем утверждать, что граф описывает абсолютно равномерный пространственный процесс, который можно назвать равномерным континуумом. Повидимому, к такому графу применять методы дискретного анализа нельзя, так как выделяемые группы и типы не будут естественными, реально существующими.
Обозначим события, когда из вершины исходит определенное количество ребер, как аk. Тогда полное поле всех событий будет: а0, a1, а2, a3, ..., аk,..., а(n-1). Здесь a0 - событие, когда вершина изолирована и из нее не исходит ни одного ребра; а1 — из вершины исходит одно ребро; а2 — из вершины исходит два ребра и т. д. Предположим, что событие аn состоит в том, что из вершины исходит среднее ожидаемое число ребер — 2m/n. При независимом распределении ребер по вершинам вероятность этого события должна быть максимальной, а вероятность отклонения и в большую, и в меньшую стороны — меньшей.
Зная вероятности прямого и обратного событий Р(у) и q(y), мы
можем построить модель биномиального распределения вероятности событий а0, а1, a2, a3, ..., ak, ..., a(n-1). Находятся они по разложению бинома Ньютона (p-q) в степени n, где р и q - вероятности прямого и обратного событий:
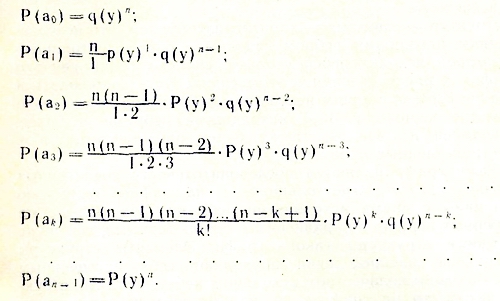

Второй тип элементов структуры графа можно условно назвать «гантели». Это пары вершин, соединенные между собой одним ребром и изолированные от остальных структурных элементов. На графиках распределения числа вершин в зависимости от числа исходящих из них ребер преобладание структур типа «гантелей» будет выражаться в явном преобладании числа вершин с одним исходящим ребром. Если в структуре графа «гантелей» больше, чем можно ожидать при случайном распределении, то такой пространственный процесс можно классифицировать как дискретный.
Третий тип структур — «звезды» — характеризуется тем, что несколько вершин с одним исходящим ребром связаны с одной центральной вершиной, из которой исходит ребер столько, сколько с ней связано вершин. На графиках распределения в этом случае будет наблюдаться явное преобладание вершин с одним исходящим ребром при значительно меньшем количестве вершин с 3-4 исходящими ребрами и более. Граф, распадающийся на звезды, также будет представлять собой дискретный пространственный процесс.
Если элементами графа являются последовательности соединенных вершин — «цепочки», «петли», «треугольники», то на графике будут явно преобладать вершины с двумя исходящими ребрами. Чем больше на графике двухреберных вершин по сравнению с однореберными, тем ниже степень дискретности структуры такого графа. Элементы структуры «плеяды» на графике распределения проявляют себя повышенным числом вершин с большим количеством исходящих ребер. Если тип структуры графа сетчатый, то на графике должно отразиться повышенное количество 3-4-реберных вершин. Распознавание форм структуры графов и их классификация, несомненно, имеют большое значение, так как разными формами структур и являются в конечном счете разные
подмножества — типы, надклассы, классы местообитаний.
При построении графов структуры растительного покрова обычно используются флористические связи разного уровня. Чрезвычайно интересно выяснить, на каком уровне сходства-различия дискретность и иерархичность структуры выявляются. наиболее полно. В связи с этим для каждого графа строились теоретические и эмпирические распределения чисел вершин в зависимости от количества исходящих из них ребер при разных уровнях сходства-различия. Учитывая связи только высокого уровня сходства, мы получаем графы с малым числом ребер. На таких графах даже при случайном распределении ребер будет много изолированных вершин. Учитывая же и более слабые связи, мы увеличиваем число ребер и снижаем таким образом вероятность существования случайных изолированных вершин, так как связность графа при этом увеличивается.
Результаты
С помощью описанной выше методики проанализированы графы флористической структуры растительного покрова двух ландшафтных районов.
Район Рау-Чуа на Западной Чукотке
Расположен в 60—70 км к юго-западу от южной оконечности Чаунской губы и характеризуется среднерасчлененным горным рельефом. Отдельные горные массивы имеют высоту более 1300 м н.у.м. и несут на себе альпийские черты. Большинство сопок - высотой 600 — 1100 м, с плоскими щебнистыми вершинами и сильно террасированными склонами. Вершины соседствующих сопок связаны седловинами в цепи и крупные массивы, которые перемежаются широкими горными распадками. Многочисленные ручьи и речки во второй половине лета обычно пересыхают.
|

Район среднего течения р. Рау-Чуа
|

Поселок Бараниха, в окрестностях которого производились исследования
|
Склони сопок довольно резко переходят в обширные суглинистые шлейфы. В местах перехода склон/шлейф имеются выровненные участки (подсклоповые террасы), которые, несмотря на хороший дренаж, интенсивно увлажняются благодаря подтоку грунтовых вод со склона сопки. На шлейфах имеются щебнистые гряды шириной от 2 до 10 м и высотой до 0,6 м, орнептированные вниз по склону, а в некоторых местах, обычно в средней и нижней частях, встречаются широкие (20 — 30 м) корытообразные ложбины стока. На склонах сопок и нагорных террасах им соответствуют каменистые полосы временных водотоков.
Отдельные наиболее высокие вершины имеют останцы, возле которых наблюдается очень большое количество грубообломочного материала. На крутых склонах сопок довольно часто встречаются щебнистые и каменистые россыпи.
В целом в районе преобладают кислые оеадочные и магматические породы: гранитониы, песчаники, сланцы. Породы основного состава отсутствуют. В долинах рек хорошо выражены пойменные террасы, переходящие в надпойму с многочисленными болотами и озерами.
Климат района континентальный, арктический, с продолжительной зимой. Снежный покров устанавливается в сентябре и лежит до июня. Средняя температура января -35, июля 10 градусов Цельсия. За год выпадает около 200 мм осадков, причем большая часть их приходится па осень, так что в конце лета даже в кочкарпых тундрах наблюдается иссушение почвы, Снеговой покров незначительный, ветрами снег сдувается с выпуклых участков рельефа и скапливается в ложбинах, местах перегибов склонов, узких распадках, долинах ручьев и речек. Однако благодаря теплой солнечной погоде в мае и нюне к началу июля все снежники стаивают и эффект нивальности проявляется не так сильно, как, например, на Восточной Чукотке.
Согласно Б. А. Юрцеву (Юрцсв, 1973; Юрцев и др. 1977), район наших исследований относится к Западно-Чукотскому округу континентально-чукотской подпровинции Арктической флористической области, подзоне южных гипоарктических тундр. Г.В. Горбацкий (1967) относит его к подзоне южных материковых тундр Притихоокеанского океанического сектора Арктики и к Чукотско-Аляскинской физико-географической области.
Флора района насчитывает 320 видов и рас сосудистых растений. Наиболее богатыми по числу видов являются (%): злаковые (12), сложноцветные (10), осоковые (10), гвоздичные (8), ивовые (6), камнеломковые (5), розоцветные (5), норичниковые (4), вересковые (3,5), ситниковые (3), бобовые (2,5). На остальные семейства приходится треть видов флоры.
В пределах ландшафтного района было выделено 59 типов местообитании или групп ассоциаций. Флористическая структура растительного покрова отражена на рис. 1.
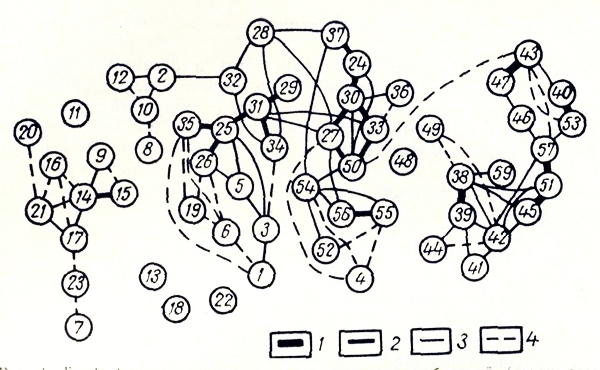
Рис. 1. Граф флористического сходства типов местообитаний (групп ассоциаций) района среднего течения p. Pay-Чуа. Флористическая структура растительного покрова ландшафтного района, 1 - сходство между типами местообитаний более 70%, 2 - сходство более 50 и менее 70%, 3 - более 30 и менее 50%, штриховые - более 20 и менее 30%.
|
14, 15. Галечники в поймах крупных рек с редкотравной луговой растительностью образуют ядро класса. Они отличаются друг от друга но степени заилепности субстрата.
9. Пойменные галечники небольших речек с редкотравными лугами.
17. Сухие участки, занятые ивняками, в поймах крупных рек. По сравнению с галечниками растительность здесь более сформирована ценотически.
21. Старые галечники первой надпойменной террасы крупных рек, с пятнистыми и куртинными разнотравно-кустарничковыми тундрами. В отличие от галечников пойм заметную роль здесь играют криогенные процессы.
23. Сухие заиленные песчапные наносы в поймах крупных рек, занятые высокотравными вейниковыми лугами. Луга ценотически неустойчивы и при незначительном усилении аллювиального процесса замешаются ивняками. В сукцессионном ряду этим лугам, по-видимому, предшествуют редкотравные луга заиленных песчаных пляжей.
20. Сухие выпуклые участки обширных надпойменных террас крупных рек. Растительность в виде кустарпичковых тундр с хорошо сформированной ценотической структурой. От старых галечников отличаются меньшей дренированноетью и лучшим развитием мохового покрова. |
16. Сырые заиленные берега временных проток и рукавов в поймах крупных рек. Растительность — сырые поименные ивняки. Местами высота кустов достигает 3 — 4 м, и их кроны образуют сомкнутый полог.
11. Сухие участки в поймах небольших речек, занятые низкорослыми ивняками. От ивняков в поймах крупных рек отличаются обедненным видовым составом, мелкоконтурностыо.
7. Заиленные берега стариц небольших речек с растительностью в виде разнотравно-осоково-пушипевых заболоченных луговин с плотной дерниной. Увлажнение избыточное, дренаж ослаблен за счет развития мерзлотных процессов.
26, 25, 35. Сырые ложбины между грядами в нижней части шлейфов склонов сопок и па склонах равнинных увалов. Кочкарные осоково-пушицево-кустарничково-моховые тундры. Увлажнение избыточное в связи с развитием мерзлоты, часто застойное, но периодически полупроточное. Преобладают делювиальные процессы, приводящие к накоплению мелкозема.
29, 31, 34. Сырые участки в верхней части шлейфов склонов сопок (подсклоповые террасы) и на конусах выноса с интенсивным минеральным подтоком. Сырые разнотравно-кустарничково-моховые звтрофные тундры. От предыдущих местообитаний отличаются степенью эвтрофикации. В облике растительности это проявляется большим участием видов разнотравья.
5. Сырые заболоченные участки надпойменных террас небольших
речек, занятые куетарничково-осоково-пушициево-моховыми тундрами.
Степень трофности высокая. Увлажнение избыточное, застойное.
3. Бровки коренного берега небольших речек и ручьев. Это хорошо дренируемые сухие торфяники, занятые ерниковыми тундрами. Один из вариантов олиготрофных местообитаний.
1. Сырые поймы ручьев в среднем и верхнем течении, занятые осоково-пушицево-моховыми тундрами. Увлажнение избыточное, проточное. Один из вариантов олиготрофного местообитания.
32. Широкие корытообразные ложбины стока в средней части шлейфов склонов. Осоково пушицево-разнотравно-моховые тундры, перемежающиеся низкорослыми сырыми ивняками. Увлажнение периодически избыточное. Уровень трофности повышенный.
28. Межгрядовые понижения в верхней части склонов с разнотравно-кустарничково-моховым тундрами. Уровень трофности повышенный.
19. Вогнутые заболоченные участки первой надпойменной террасы крупных рек. Увлажнение избыточное, застойное. Растительность в виде пушицево-осоково-кустарничково-моховых болот.
6. Заболоченные надпойменные террасы небольших речек, запятые
пушицево-осоково-моховыми болотами.
22. Заболоченные участки в пойме р. Рау - Чуа. Разнотравно-осоково-моховые минеральные эвтрофные болота. Высокая трофность сочетается с аллювиалыюстью и ослаблением криогенных процессов.
18. Заболоченные берега озер первой надпойменной террасы Рау-Чуа. Растительность в виде разнотравно-осоково-пушицево-моховых заболоченных луговин.
13. Старичные озера. Прибрежно-водная растительность.
8. Редко затопляемые галечники надпойменных террас небольших речек , притоков Рау-Чуа, в узких распадках, с нивальным режимом. Растительность в виде куртинных и пятнистых кустарничково-разнотравных нивальных луговинных тундр. Этот тип местообитаний характеризуется сочетанием аллювиального и нивального режимов.
10. Невысокие надпойменные террасы небольших речек в узких распадках с полунивальным режимом. Растительность в виде довольно высокорослых разнотравных ивняков. От предыдущего отличается ослаблением аллювиальности.
12. Заиленные галечники в поймах небольших речек . Растительность куртинная, разнотравно-кустарничковая, с элементами нивалыюсти.
24, 37. Сухие грядообразные повышения на склонах равнинных увалов. Заняты пятнистыми разнотравно-кустарничково-мохово-лишайниковыми тундрами. Увлажнение несколько недостаточное. Колювиальные процессы сочетаются с делювиальными.
27, 30, 33, 50. Сухие щебнистые участки на шлейфах склонов сопок. Пятнистые разнотравно-кустарничково-лишаи пиковые тундры. По сравнению с 24, 37 сильнее выражены колювиальные процессы. Увлажнение периодически недостаточное.
55, 56. Щебнистые пологие склоны сопок северной экспозиции, занятые пятнистыми разнотравно-кустарничково-лишайниковым и щебнистыми тундрами. Колювиальные процессы явно преобладают над делювиальными. Увлажнение большую часть сезона недостаточное, лишь периодически нормальное. Очень хорошо выражены криогенные процессы
54. Первые надпойменные террасы небольших речек. Старые галечники. Разнотравно-кустарничковыетундры.
52. Высокогорные террасы на высоте более 800 м н.у.м. Разнотравно-кустарничково-мохово-лишайниковые тундры.
36. Щебнисто-суглинистые уступы на шлейфах склонов, занятые сухими разнотравно-кустарничковыми тундрами.
48. Берега ручьев в нижней и средней частях некрутых склонов
сопок южной экспозиции, с растительностью в виде разнотравно-мохово-кустарничковых тундр.
43, 47. Сухие щебнистые нагорные террасы в нижнем и среднем
частях склонов, запятые пятнистыми разнотравно-кустарничково-мохово-лишайниковыми тундрами. Колювнальпые процессы здесь, видимо,
незначительно преобладают над делювиальными. Увлажнение недоста
точное.
40. 53. Некрутые склоны сопок южной экспозиции. Пятнистые
кустарничково-лишайниковые щебнистые тундры.
45, 51, 57. Привершинные части склонов сопок. Куртинные и фрагментарные разнотравно-лишайниковые тундры. Колювиалыше процессы развиты очень сильно. Увлажнение явно недостаточное.
38, 39. Слегка выпуклые участки в нижней части склонов сопок южной экспозиции. Разнотравно-кустарничково-лишаппиковые тундры со слабыми элементами остепнения. Мелкозем накапливается. Увлажнение почти постоянно недостаточное.
44. Каменистые россыпи в нижней части склонов южной экспозиции. Здесь развиты куртинные, фрагментарные разнотравные тундры.
42. Временные каменистые водотоки в нижней и средней частях южных склонов сопок. Растительность в виде весьма высокопродуктивных куртинных разностравно-кустарничково-лишайниновых тундр.
59. П лоские вершины сопок, занятые куртинными разнотравно-лишайниковыми тундрами. Субстрат щебнистый и шебнисто-каменистый. Мелкозем активно выносится, но дренаж из за развития криогенных процессов незначительный.
41. Щебнистые, слегка вогнутые участки в нижней части южных
склонов сопок. Зимой хорошо укрываются снегом, летом, благодаря
южной экспозиции и положению в нижней части склонов, рано протаивают. Почвы интенсивно гумуснрованы. Растительность разнотравно-злаковая, с явственными элементами остепнения. Тундровые гипоарктичсские кустарнички отсутствуют. В ландшафте представлены очень
мелкими контурами.
46. Каменистые водотоки в верхней части склонов сопок (курумы). Растительность в виде куртинных разнотравно лишайниковых тундр. Колювиальные процессы здесь развиты в наибольшей степени.
49. Сусликовпны. Щебнисто-суглннистые бугры в нижней части склонов
южной экспозиции имеют зоогенное происхождение. Растительность в виде сухих ксеромезофитных луговин. Специфика этих местообитаний
связана с деятельностью длиннохвостого суслика. Усиленный дренаж,
глубокое протанвание гpунтa, занос семян многих видов, удобрение
экскрементами (повышенная трофность).
На рис . 1 типы местообитаний, образующие «плеяды» вершин, формируют классы местообитаний. Типы местообитаний, слабо связанные с выделенными классами, либо уникальны флористически за счет видов, встреченных только в этом местообитании (13; 18; 29), либо представляют собой флористически сильно обедненный вариант более богатого типа местообитания (II), либо специфичность их флористического разнообразия обусловлена сочетанием видов с весьма разными экологическими амплитудами (при этом виды не обязательно редки в районе) (48) (рис. 1). Однако во всех трех случаях флористическое несходство типов местообитаний связано со спецификой экологических условий, т. е. с особенностями экологических режимов.
В одних и тех же фитоценозах могут произрастать виды и с узкой, и с широкой экологической амплитудой. В нашем случае это наблюдается в нескольких типах фитоценозов: 8; 10; 12; 49.
На основе графической модели флористической структуры растительного покрова ландшафтного района разработана шкала оценки эколого-ценотических амплитуд видов (Галанин, 1977):
• Стенотопные виды. Встречаются в одном-двух местообитаниях, относящихся к одному классу. Если вид встречен в двух—трех местообитаниях, то флористическое сходство между ними очень велико (относятся к одному ядру).
• Гемистенотопные виды. Встречаются в большинстве местообитаний
одного класса и в одном-двух местообитаниях соседнего класса.
• Гемиэвритопные виды. Встречаются в большинстве местообитаний
двух классов и в отдельных местообитаниях третьего класса.
• Эвритопные виды. Встречаются в большинстве типов местообитаний трех классов и некоторых типах местообитаний четвертого класса.
Оценка эколого-ценотической амплитуды видов по этой шкале имеет ландшафтно-экологическое обоснование и облегчает сравнение экологических амплитуд у видов с разной экологией в пределах одного района. Например, у обитателей пойм и обитателей щебнистых склонов. Наряду с широтой эколого-ценотической амплитуды имеет смысл оценивать степень соответствия (адекватности) эколого-ценотического распределения конкретного вида в пределах ландшафтного района интегральной флористической структуре растительного покрова этого района. Было показано, что виды могут иметь «тривиальное» и «оригинальное распределение» в пределах ландшафтного района (Галанин, 1977).
Полученные данные показывают, что изменения растительного покрова в пределах ландшафтного района связаны с такими экологическими факторами, как интенсивность и характер увлажнения, уровень трофности, уровень нивальности, подверженность субстрата перемещениям, развитие криогенных процессов и др. Однако названные, экологические факторы не могут служить осями многомерной прямоугольной, системы координат, характеризующей экологическое пространство ландшафтного района, так как все эти показатели довольно тесно скоррелированы друг с другом. Это значит, что изменение одного из факторов, например степени увлажнения, немыслимо без изменения других факторов, например уровня трофности. В качестве же осей координат экологического пространства должны быть выбраны какие-то иные характеристики, непосредственно связанные с объемом, скоростью и степенью замкнутости биологического круговорота химических элементов, величиной потока энергии, проходящего через экосистему за год и, возможно, с уровнем негэнтропии экосистемы.
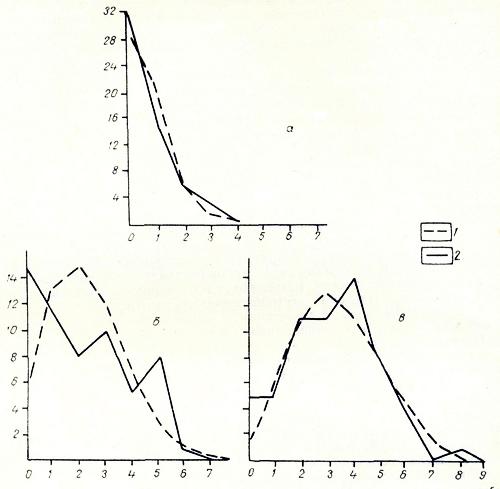
Рис 2. Распределение числа вершин в зависимости от числа исходящих из них ребер графа флористического сходства типов местообитаний района среднего точения р. Рау-Чуа: а - сходство 70%, б - сходство 50%, в - сходство 30%; 1 - распределение ожидаемое, 2 - распределение наблюдаемое
|
На рис. 2 изображены, кривые распределения числа вершин графа флористического сходства типов местообитаний района Рау-Чуа. Вначале мы учитывали только ребра, соответствующие флористическому сходству более чем на 70%. При данном числе вершин n = 58 и данном числе ребер m =18 весьма велика вероятность существования в структуре графа «изолятов» и «гантелей». Эмпирическое распределение вершин в зависимости от числа исходящих ребер отличается от теоретического очень слабо (хи-квадрат = 2,29). Следовательно, учитывая только наиболее высокое сходство, мы получаем структуру весьма близкую к равномерной, ее можно охарактеризовать как равномерный континуум.
Приняв более низкий уровень сходства (более 50% и менее 70%), мы увеличили число ребер до 59, т. е. в среднем на одну вершину теперь приходится 2,1 ребра а не 0,7 как в предыдущем случае. Структура графа становится явно дискретной, так как число «изолятов» существенно выше, чем ожидается теоретически. Кроме того, значительно больше число вершин с пятью исходящими ребрами. Пространственный процесс, моделируемый этим графом, имеет явно неравномерный характер ( хи-квадрат = 30,12).
При уровне сходства больше 30%т менее 50% эмпирическая кривая очень слабо отличается от теоретической (хи-квадрат = 11,03). Тем не менее мы можем говорить о слабой дискретности структуры («изолятов» несколько больше
ожидаемого) и некоторой неравномерности пространственного процесса. Причина дискретности флористической структуры растительного покрова Рау-Чуа на уровне сходства выше 50%, по-видимому, связана с контрастностью рельефа и резкой континентальностью климата. В таких условиях участки с различным типом экологического режима оказываются резко разграниченными. С другой стороны, район Рау-Чуа характеризуется давно сложившейся флорой и растительностью, развитие которых происходило в основном преемственно (Галанин, 1977). В этом случае эколого-ценотические амплитуды большинства видов пришли в соответствие с градиентами экологической структуры ландшафта. Дискретность растительного покрова может иметь и ценотическое происхождение, когда в результате межвидовой конкуренции виды как бы «загоняются» в оптимальные для них местообитания и «изгоняются» из обитаний неоптимальных. |
В нашем случае ценотический фактор, видимо, не является решающим, так как в ландшафте много сообществ с невыработанной ценотической структурой. Говоря о неравномерности флористической структуры растительного покрова района, следует всегда отмечать, на каком уровне сходства групп ассоциаций она наблюдается. В связи с этим есть основание оценивать не только степень неравномерности структуры, но и уровень этой неравномерности.
Район окрестностей Чаунского стационара (Западная Чукотка)
Чаунская низменность включается в общий контур Анюйско-Чукотской горной области Верхояно-Чукотской мезозойской складчатой страны (Баранов, Бискэ, 1964) и представляет собой незатопляемую морем окраинную часть области шельфа, простиравшегося в позднечетвертичное время значительно севернее о-ва Врангеля. Горнодолинные ледники среднечетвертичного и позднечетвертичного оледенений не покрывали эту территорию (Баранов, Бискэ, 1964).
Рельеф района очень однообразен. Это совершенно плоская озерно-аллювиальная равнина, расчлененная множеством русел и представляющая собой дельтовую область рек, текущих с Анюйского и Чукотского нагорий. Гидрографическая сеть здесь настолько густа, что площадь суши едва ли больше площади, занятой текущими водами и озерами. Течение рек медленное, оно периодически подпруживается приливами и нагонными ветрами с моря. По мере удаления от устья реки аллювиальные осадки становятся крупнее и примерно в 14—20 км вверх по течению наряду с песчаными пляжами встречаются обширные галечники. Поверхность равнины поднята над урезом воды на 2—3 м в нижнем течении и на 3—5 м в 15—20 км от устья. Неудивительно, что русла рек Чаунской низменности непрерывно меняют свое положение и вся равнина покрыта слоем современных илистых, песчаных и галечных наносов. Перемещение русел происходит в половодье.

Чаунский биологический стационар
|
Во время северных и северо-западных штормовых ветров в реку нагоняется так много воды, что все устье заливается на 10—15 км. Благодаря этому приморские галофиты заходят далеко вверх по реке. Часть озер периодически затопляется водой приливов, что угнетающе действует на многие виды растений. Во время южных ветров уровень воды в реке падает на I —2 м и обнажаются огромные песчаные отмели площадью в десятки и сотни гектаров. Эти отмели совершенно свободны от растительного покрова. В такие моменты заросли прибрежно-водных злаков — арктофилы и дюпонции — похожи на роскошные сухие пойменные луга.
Вся низменность в районе стационара представляет собой широкую речную долину. В мезорельефе выражены многочисленные округлые углубления, вероятно, термокарстового происхождения, с торфяным дном, почти до краев наполненные водой. Уровень воды в таких озерках на 2 — 3 м выше, чем в реке. Наряду с явно термокарстовыми широко распространены старичные озера с песчаным дном, которые к тому же ориентированы в виде цепочек и соединены друг с другом постоянными или временными протоками. Во время паводков эти озера сообщаются с основным руслом.
Изредка над равниной возвышаются торфяные бугры — балгуньяхи (гидролаколиты) высотой 7—8 и до 30 м в диаметре, на вершинах которых можно встретить растения, обычно характерные для сухих горных тундр. |
Кроме тсрмокарстовых процессов в Чаунекой низменности широко распространено морозное растрескивание субстрата, в результате которого образуются многоугольники диаметром от 2 до 30 м и более. Таким образом, несмотря на однообразный макро- и мезорельеф, аллювиальные и термокарстовые процессы, явления мерзлотного выпучивания и растрескивания субстрата, а также периодическое засоление во время штормов создают довольно своеобразный и обширный спектр экологических режимов.
В районе стационара хорошо различаются две полосы: 1 — полоса, подверженная засолению, тянется от побережья залива вверх по течению на 10—'15 км; 2— полоса, не подверженная засолению, начинается в 10—15 км от побережья и простирается к югу. Граница между этими полосами весьма нечетка, так как уровень воды в заливе поднимается на разную высоту в зависимости от силы штормов.
На западе, юге и востоке Чаунскую низменность обрамляет полого-увалистая равнина с отдельными сопками высотой до 600 м н.у.м. Одна из таких сопок (гора Нейтлин) находится в 20 км к югу от стационара. Пологий склон горы в нижней части сложен глинистыми сланцами, далее сланцы сменяются роговиками, окружающими гранитную вершину (батолит). Склоны горы сильно террасированы, ступени террас около 30 м высотой. Как небольшие террасы, так и склоны в верхней части сложены глыбами гранита. Лишь на обширных нагорных террасах в нижней части склонов сопки скапливается мелкозем и развиты разнотравно-кустарничково-моховые тундры, в которых задернованность поверхности достигает 70—80%. Со всех сторон гору окружают обширные шлейфы, занятые бугорковатыми гипоарктическими осоково-кустарничковыми тундрами.
Для Чаунской низменности характерен низкоарктический климат с продолжительной морозной зимой (средняя температура января - 35° С) и относительно теплым для данной широты летом (средняя температура июля 10° С). Осадков за год выпадает около 200 — 300 мм, причем 100 мм из них в жидком виде (Клюкин, 1960). Вегетационный период длится 3—3,5 мес.
В системе ботанико-географического и геоботаничсского районирования район Усть-Чаунского биологического стационара занимает такое же положение, как и район среднего течения р. Рау-Чуа.
По нашим данным, флора района насчитывает 252 вида и расы высших сосудистых растений. Ведущими по числу видов семействами являются (%): злаки (13,5), осоковые (12,3), слржноцветные (10), ивовые (6), гвоздичные (5,5), лютиковые (5), крестоцветные (4,8), норичниковые (4,8), камеполомковые (4), розоцветные (6), гречишные (3) и бобовые (2,8).
Всего в пределах ландшафтного района выделено 30 типов местообитаний, или групп ассоциаций. Флористическое сходство по Жаккару рассчитывалось, между всеми возможными парами типов местообитаний. На основе матрицы бинарного (попарного) сходства типов местообитаний построен граф (рис. 3). Нумерация типов местообитаний на графе соответствует их нумерации в приведенном ниже конспекте.
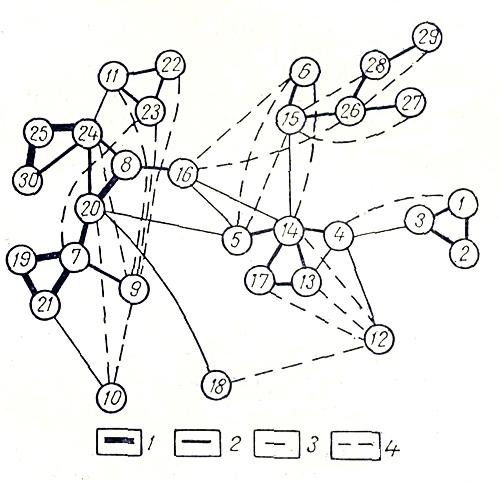
Рис. 3. Граф флористического сходства типов местообитаний (групп ассоциаций) района окрестностей Чаунского биологического стационара. Флористическая структура растительного покрова ландшафтного района. 1 - сходство более 50%; 2 - сходство более 40%, но менее 50%; 3 - сходство более 30%, но менее 40%, 4 - сходство более 20%, но менее 30%.
|
1. Слегка вогнутые участки приморской террасы, часто заливаемые
морскими водами. Растительность в виде низкотравных луговин
(марши). Увлажнение постоянно слегка избыточное.
2. Относительно сухие, чуть выпуклые участки приморской террасы,
часто заливаемые морскими водами. Сухие пизкотравные луговины
(марши).
3. Песчаные, сильно заиленные пляжи в устье реки. Растительность в виде редкотравных луговин (марши). На этих местообитаниях
засоление сочетается с аллювиальными процессами.
12. Сырые заиленные пляжи вне полосы засоления. Растительность
в виде редкотравных пойменных лугов.
4. Сухие песчаные речные пляжи, заливаемые только во время
сильных паводков. Растительность в виде рсдкотравных
лугов с примесью галофитов.
13. Сухие песчаные пляжи и галечники вне полосы засоления.
Редкотравные пойменные луга. |
14. Обрывы коренного берега вне полосы засоления. Куринные и фрагментарные пойменные луга. Субстрат обычно торфянисто-минеральный. Уровень трофности за счет хорошей минерализованности
торфа несколько повышенный. Увлажнение периодически недостаточное.
17. Задернованные берега стариц и временных проток. Растительность в виде комплекса перемежающихся ивняков, ольховников и лужаек. Аллювиальный процесс ослаблен. Дренаж довольно интенсивный.
5. Обрывы коренного берега в полосе засоления. Растительность куртинная кустарничково-травяиистая. Субстрат обычно торфянисто-минеральный. Увлажнение периодически недостаточное.
18. Задернованные днища стариц вне полосы засоления. Осоково-пушицевые тундровые болота. Флористически сильно обособлены.
23. Тсрмокарстовые озера вне полосы засоления. Водная и прибрежно-водная растительность.
22. Старичные озера вне полосы засоления. Водная и прибрежно-водмая растительность.
11. Старичные озера в полосе засоления. Водная и прибрежно-водная растительность.
15. Наиболее обдуваемые, хорошо дренированные участки коренного берега с бугорковатым микрорельефом. Растительность в виде кустарничкопых мохово-лишайпиковых тундр. Почва торфянистая, увлажнение периодически недостаточное.
26. Щебнистые выходы на шлейфах склонов сопок. Как правило, это невысокие уступы на шлейфе. Растительность в виде разнотравно-кустарничковых мохово-лшпайниковых пятнистых тундр. Наряду с
делювиальными процессами здесь хорошо развиты и колювиальные,
но они слабо преобладают над делювиальными.
28. Сухие щебнистые склоны сопок в нижней части, занятые
пятнистыми разнотравно-кустарничковыми тундрами. Увлажнение периодически недостаточное. Колювиальные процессы преобладают.
29. Крупнокаменистые склоны в верхней и средней частях с куртинной
разиотравно-кустарничковой тундровой растительностью. Наблюдается
явное преобладание выноса мелкозема. Хорошо развиты колювиальные
процессы.
27. Относительно сухие участки у подножия склонов сопок.
Нивальные кассиоповые мохово-лишайниковые тундры. Зимой здесь
скапливается снег, но не сильно, так что степень нивальности довольно
слабая.
6. Наиболее высокие, обдуваемые и хорошо дренированные
краевые участки коренного берега рек в полосе заселения. Микрорельеф
бугорковатый. Растительность в виде кустарничковых мохово-лишайниковых тундр. Эти местообитания характеризуются пролювиальным
режимом.
10. Незаболоченные, нормально увлажненные песчаные берега в полосе засоления. Ивнячково-злаковые сообщества.
21. Сырые заболоченные берега озер вне полосы засоления. Осоковые тундры со значительной примесью разнотравья. Трофность несколько повышенная.
19. Вогнутые участки надпойменных террас. Полигонально-вадиковые
тундровые болота. Увлажнение избыточное, застойное. Почва торфянисто-
глеевая.
7. Вогнутые участки надпойменных террас в полосе засоления.
Полигонально-вадикрвые тундровые болота. От предыдущего типа отличается только периодическим затоплением и слабым засолением.
9. Заболоченные берега озер в полосе засоления. Сырые осоковые тундры с участием видов-галофитов.
20. Слегка выпуклые участки надпойменных террас в полосе не
подверженной засолению. Буторковатые мохово-кустарничковые тундры с участием кустистых лишайников. Увлажнение периодически избыточное, периодически недостаточное. Почвы торфянистые.
8. Слегка выпуклые участки надпойменных террас в полосе засоления. Бугорковатые мохово-кустарничковые тундры с участием кустистых лишайников.
24. Нижние и средние части шлейфов склонов. Осоково-кустраничково-моховые тундры.
16. Небольшие депрессии в краевой части коренного берега рек, стариц и временных проток (неглубокие овраги) вне полосы засоления. Растительность и виде зарослей ольхового стланика со значительным участием разнотравья. Увлажнение нормальное. Зимой скапливается снег, но нивальпый режим не выражен.
25. Верхние части шлейфов склонов и конусов выноса. Осоково-куетарничково-разнотравные тундры. Увлажнение слегка избыточное.
Трофность повышенная.
30. Обширные нагорные террасы, занятые разнотравно-кустарничковыми моховыми сырыми тундрами. Режим увлажнения весьма непостоянный: в течение вегетационного периода увлажнение может быть избыточным, нормальным и недостатичным. Режим элювиально-пролювиальный.
Граф флористической структуры растительного покрова района окрестностей Усть-Чаунского биологического стационара (рис. 3) показывает, что выявленная структура, как и в районе Рау-Чуа, иерархична и наряду с типами местообитаний здесь также могут быть выделены классы меетообитаний. Однако, и отличие от района Рау-Чуа, в районе Чаунского стационара имеется класс местообитаний, связанных с особым экологическим режимом — засолением. Это маршевые луга и луговины. В самостоятельный класс выделились также озера с прибрежно-водной растительностью, широко распространенные в Усть-Чаунской низменности. Особенность растительности Чаунской низменности — наличие ольховников (16), занимающих промежуточное положение между четырьмя классами местообитаний. По всей вероятности, это реликтовый тип растительности, который в эпоху голоце нового оптимума занимал здесь доминирующее положение (Галанин, 1980). Эта гипотеза была недавно подтверждена пашей находкой в районе оз. Эльгыгытгын кусков древесины кустарниковой ольхи в погребенном торфянике. Радиоуглеродная датировка образцов показала, что даже на высоте около 500 м н.у.м. в истоках рек Чауна, где сегодня ольховник совершенно отсутствует, примерно 9,3 тыс. лет назад встречались его заросли.
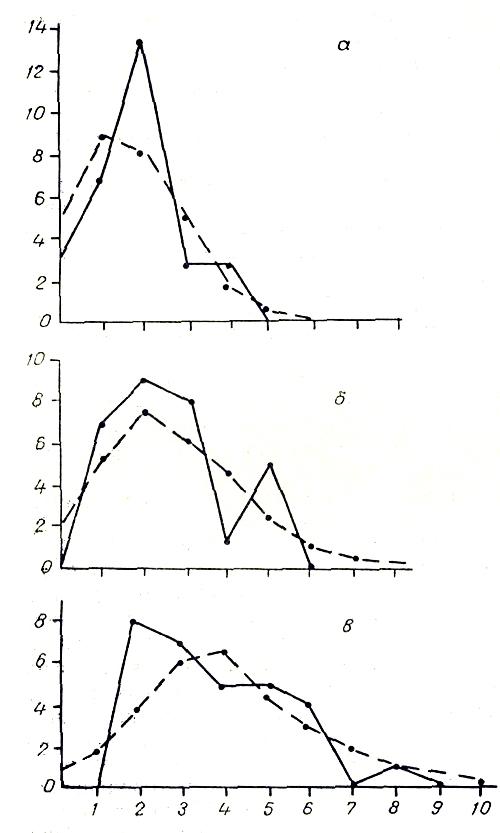
Рис. 4. Распределение числа вершин в зависимости от числа исходящих из них ребер графа флористического сходства типов местообитаний района Усть-Чаунского стационара: а - сходство болеее 40%; б - сходство более 30%, но менее 40%, в - сходство более 20%, но менее 30%.
|
Так же как и на Рау-Чуа, в районе стационара четко выделился класс пойменных местообитаний с аллювиальным экологическим режимом, класс местообитаний равнинных тундр с двумя подклассами по градиенту увлажнения и класс местообитаний сухих горных пятнистых разнотраино-кустарничковых тундр. Если тип экологического режима последнего класса можно в целом охарактеризовать как колювиальный, то тип экологического режима в классе местообитаний в основном определяется преобладанием делювиальных процессов. Пролювиальный режим также отражается в характере растительного покрова.
Таким образом, с помощью графов флористического сходства типов местообитаний можно выявить основные экологические градиенты растительного покрова, внутри ландшафтного района. В Усть-Чаунской низменности изменение растительного покрова на лаидшафтно-акологическом уровне организации связано с изменением интенсивности и характера увлажнения, степени засоленности, уровня трофности, нивальности. Здесь мы выделяем следующие тины экологических режимов: маршевый, аллювиальный, делювиальный, колювиальный, пролювиальный и нивальный. Последний тип режима выражен слабо.
На рис. 4 изображены эмпирические и теоретические распределения вершин в зависимости от числа исходящих из них ребер в графе флористического сходства типов местообитаний района Усть-Чаунекого стационара. При уровне сходства более 40% на 30 вершин приходится 28 'ребер (по 1,9 ребра на одну вершину). Фактическое распределение вершин в этом случае отличается от случайного незначительно (хи-квадрат = 6,78). Правда, имеется достоверное превышение фактического числа вершин с двумя исходящими ребрами над их ожидаемым числом, что говорит о сильно выраженной континуальности структуры графа. Неравномерность или гетерогенность данного пространственного процесса выражена чрезвычайно слабо, так как ход кривой довольно плавный. При уровне сходства более 30% эмпирическое распределение становится почти прерывистым. Здесь больше ожидаемого вершин с одним, двумя, тремя и пятью ребрами, но меньше ожидаемого вершин с четырьмя ребрами. В целом пространственный процесс сильно неравномерный (хи-квадрат = 9,65), но континуальный, так как фактическое число «изолятов» не превышает ожидаемого.
При уровне сходства более 20% эмпирическая кривая существенно отличается от теоретической (хи-квадрат = 12,21). Однако континуальность структуры при этом усиливается, так как вершин с одним исходящим ребром значительно меньше ожидаемого. Несмотря на то что при учете связей выше 40% множество вершин графа распалось на восемь абсолютно изолированных подмножеств, мы можем говорить лишь о слабой неравномерности структуры. Это связано с тем, что при малом числе ребер связность графа мала и вероятность его случайного распада на подграфы очень велика. Сравнивая графы, можно заметить, что наиболее полно и четко неравномерность структуры графа выявляется на уровне сходства 30%. Именно этот уровень был выбран нами при выделении классов местообитаний (Галаниин, 1980).
Сравнивая флористическую структуру растительного покрова районов Рау-Чуа и Чаунского стационара, можно заметить, что последняя характеризуется значительно более высокой континуалыюстыю, что, несомненно, связано с меньшей контрастностью рельефа района. Кроме того, континуальность растительности здесь может усиливаться постепенным изменением степени затопляемости дельты морскими соленными водами, а этот фактор, несомненно, играет значительную роль и распределении растений. |
Выводы
• Графы попарного (бинарного) флористического сходства групп
ассоциаций (или типов местообитаний), являются моделями флористической структуры растительного покрова, отражающими экологическую
структуру ландшафтного района.
• Графические модели флористической структуры растительного покрова можно использовать при осуществлении ландшафтно-экологического ботанического мониторинга. В этом случае пробные площади должны
быть постоянными и отмаркированными на местности. Периодические
переописания растительности этих площадей 1 раз в 15-20 лет позволят
получить временной ряд графовых моделей структуры растительного
покрова, что и составит основу мониторинга.
• Анализ степени равномерности структуры графов показал, что
понятиями «континуальность» и «дискретность» свойства структуры
растительного покрова не исчерпываются. Дискретность растительности
должна пониматься как крайняя степень неравномерности ее структуры,
а континуальность, напротив, как крайняя равномерность распределения
флористического сходства между единицами растительности.
• При оценке неравномерности флористической структуры растительного покрова целесообразно находить и учитывать уровень
флористического сходства, при котором неравномерность имеет максимальное значение.
• Сравнение структуры растительного покрова двух ландшафтно-ботанических районов Чукотки показало, что степень неравномерности
их структур неодинакова. Более неравномерна флористическая структура района Рау-Чуа.
• Для растительного покрова района Рау-Чуа характерна слабая
дискретность типологической структуры, особенно при уровне флористического сходства между группами ассоциаций более 50%.
Литература
Баранова Ю.П., Бискэ С.Ф. История развития рельефа Сибири и Дальнего Востока // Северо-Восток СССР. М.: Наука. 1964. 290 с.
Галанин А.В. Aнализ распределения растений по типам местообитаний (на примере района равнинно-горного ландшафта среднего течения р. Рау-Чуа Западной Чукотки // Ботан. журн. 1977. Т. 62, N 8. С. 1045-1060.
Галанин А. В. Флора и растительность Усть-Чаунского биологического стационара (Западная Чукотка) // Ботан. журн. 1980. Т 65. N 9. С. 1171-1187.
Галанин А.В. Опыт сопряженного анализа типологических структур конкретных флор // Теоретические и методологические проблемы сравнительной флористики. Л.: Наука. 1987. С. 167-189.
Горбацкий Г.В. Физико-географическое районирование Арктики. Л.: Изд-ко ЛГУ. 1967. Полоса материковых тундр. Ч. I. 221 с.
Клюкин Д.Ф. Климатологический справочник по Северо-Востоку СССР. Магадан: Кн. изд-во, 1960. 173 с.
Сочава В. Б. Внедение в учение о геосистемах. М.: Наука, 1978. 320 с.
Юрцев Б.А. Ботанико-географическая зональность и флористическое районирование Чукотской тундры // Ботан. журн. 1973. Т. 58. N 7. С. 786-792.
Юриев Б. А.. Толмачев А.И., Ребристая О.В. Флористическое ограничение и разделение Арктики // Арктическая флористическая область, Л.: Наука, 1978. С. 9-104.
Фотографии взяты с сайта Института биологических проблем Севера и сайта Чукотского национального округа. Авторам фотографий приношу свою благодарность.

