|
|
|
|
|
|
Метод графов при составлении дихотомических ключей
|
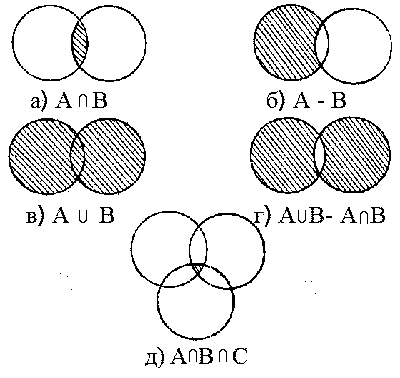 |
В процессе составления дихотомического ключа путем логических операций из элементарных альтернативных признаков могут быть получены сложные альтернативные признаки (рис. слева). Рис. 1. Способы образования сложных морфологических признаков из признаков элементарных с помощью основных логических операций.
Например, имеются два элементарных признака: А - «Листья зубчатые по краю» и В - «Листья сердцевидные». Путем пересечения этих двух элементарных альтернативных признаков – операции умножения на множествах (рис. 1а) - можно получить сложный альтернативный признак А |
Для морфологической характеристики видов рода Лютик флоры Калининградской области нами взято 30 альтернативных признаков. Большинство из них элементарные. Ниже приводится их список с порядковыми номерами. Далее в матрице (табл. 1) и на грвфе (рис. 2) признаки приводятся под теми же номерами.
1. Стебли в основании с клубневидным утолщением.
2. Стебли ползучие (распростертые и в узлах укореняющиеся).
3. Растения однолетние.
4. Чашелистиков меньше пяти.
5. Растения с развитым горизонтальным корневищем.
6. Все листья цельные.
7. Прикорневые листья округло-почковидные в диаметре менее пяти сантиметров.
8. Прикорневые листья округло-почковидные в диаметре около десяти сантиметров.
9. Листья тройчато-сложные.
10. Листья тройчато-надрезанные.
11. Листья рассеченные на пять долей.
12. Доли прикорневых листьев широко-ромбические.
13. Доли прикорневых листьев узкие.
14. Доли стеблевых листьев широкие.
15. Доли стеблевых листьев узкие.
16. Плоды с шипиками.
17. Плоды с бугорками.
18. Плоды голые.
19. Цветки мелкие (до 15 мм в диаметре).
20. Чашелистики снизу опушенные.
21. Цветоложе удлиненное (цилиндрическое).
22. Носик плодов хорошо развит.
23. Носик плодов крючковидно загнут.
24. Цветоножки бороздчатые.
25. Стебли голые.
26. Стебли прижато-волосистые.
27. Стебли оттопыренно-волосистые.
28. Чашелистики во время цветения вниз отогнуты (прижаты к цветоножке).
29. Доли стеблевых листьев явно зубчатые.
30. Доли стеблевых листьев по краю явно гладкие.
 |
Всего на территории Калининградской области встречается 12 видов лютиков (табл. 1). Анализируя гербарные образцы, мы альтернативно охарактеризовали каждый из этих видов по каждому признаку. В табл. 1 (+) заменяет альтернативное значение «да», а пропуск обозначает значение признака «нет». Не все выбранные признаки для нашего множества видов являются делящими или диагностическими. С помощью делящего признака мы можем разделить множество видов на два непустых подмножества. Так, по первому признаку мы можем разделить наши лютики на подмножество с одним видом (R. bulbosus) и подмножество, включающее все остальные 11 видов. Такой признак является единично диагностическим, так как сразу «выщепляет» из множества один конкретный вид.
Табл. 1. Матрица распределения видов рода Ranunculus в многомерном пространстве элементарных альтернативных морфологических признаков. Здесь и далее значения признаков "да" обозначены знаком (+), а "нет" - знаком (-); знак (?) обозначает то, что для данного вида признак не альтернативен: у одних особей он имеет значение "да", у других особей этого вида - значение "нет". |
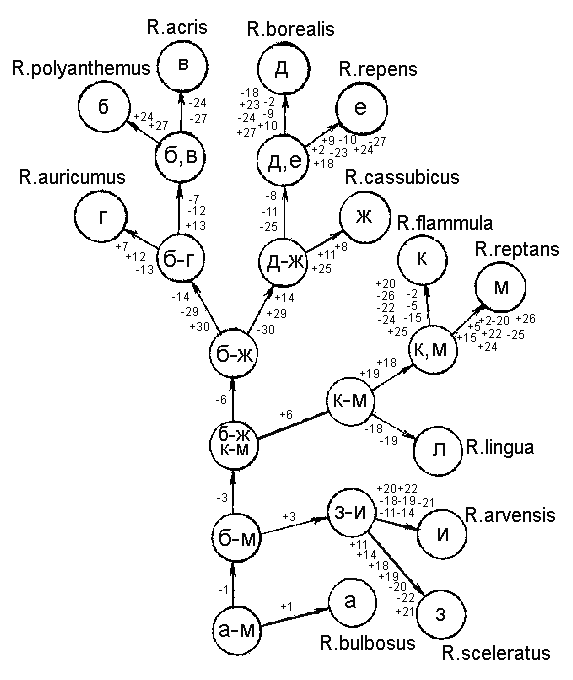 |
Рис. 2а. Графическая модель смешанного ключа для определения видов рода Ranunculus флоры Калининградской области
Признак № 27 диагностический, но не единично, так как делит множество видов на два подмножества, включающие более чем по одному виду: (R. bulbosum, R. polyanthemus, R. borealis, R. arvensis) и (R. acris, R. auricumus, R. repens, R. cassubicus, R. sclerathus, R. flammula, R. linqua, R. reptans). Признаки же № 4 и № 17 для нашего множества делящими не являются и их использование в ключах бессмысленно. Для удобства в дальнейшей работе виды лютиков обозначены строчными буквами (табл. 1). На каждой ступени определительного ключа следует использовать только делящие признаки. Последовательное разбиение множество таксонов с помощью альтернативных признаков удобно изображать в форме ветвящегося графа (рис. 2). Ясно, что на основании одной матрицы можно получить несколько типов разбиения, то есть несколько графов и, следовательно, можно построить несколько ключей. Но при этом одни из них будут более оптимальными, другие – менее. При всех прочих равных условиях боле оптимален граф, основанный на признаках макроскопических и поэтому легко различимых. Второй принцип оптимальности связан с использованием признаков, которые можно проследить на протяжении значительной части сезонного развития растения. Третий принцип оптимальности связан с наличием на каждой ступени деления не одного, а нескольких признаков – диагностических дублей. Наконец, четвертый принцип оптимальности связан с количеством ступеней деления или длиной пути определения. Более оптимален, естественно, граф с меньшей длиной пути определения. Такие графы имеют большее число разветвлений (они более ветвистые) и позволяют в параллельных ветвях повторно использовать одни и те же признаки. |
Признаками дублями являются такие, которые дают абсолютно сходные разбиения исходного множества данной ступени деления. Такие элементарные признаки-дубли на данной ступени деления образуют сложный признак по принципу пересечения, но при полном совпадении. При образовании таких сложных признаков следует использовать только признаки дубли. Нарушение этого правила приведет к нарушению законов формальной логики.
В принципе ключи для определения могут быть трех типов: 1 – моноподиальный, когда на каждой ступени деления выщепляется по одному виду; 2 – полный дихотомический, когда на каждой ступени исходное множество таксонов делится на два равных подмножества; 3 – смешанный, когда совмещаются элементы полного дихотомического деления на одних ступенях с элементами моноподиального деления на других.
Моноподиальное деление возможно только при наличии для множества в целом и исходных подмножеств на каждой ступени делегия единично диагностических признаков. В этом случае для разбиения и таксонов требуется (п – 1) признак, поскольку повторное использование одного признака на разных ступенях деления здесь невозможно, так как каждое ответвление требует отдельный признак.
При полном дихотомическом и смешанном делении возможно повторное использование одних и тех же признаков в разных разветвлениях графа. При полной дихотомии, например, множество из 8 видов можно разделить с помощью всего лишь трех признаков, а множетсво из 128 видов с помощью 7 признаков. Разумеется, эту логическую возможность в полном виде реализовать трудно, так как необходимо дополнительное условие – каждый признак должен делить множество на равные части, первый – на две части, второй – на четыре, третий – на восемь равных частей и т. д.
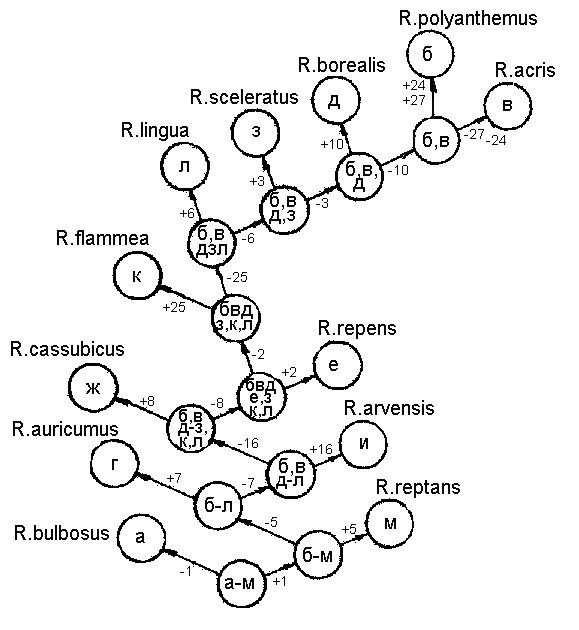 |
Рис. 2б. Графическая модель моноподиального ключа для определения видов рода Ranunculus флоры Калининградской области
Обратимся к нашему конкретному примеру с лютиками Калининградской области (рис. 2а). Исходное множество видов (а-м) на первой ступени с помощью единично диагностического элементарного альтернативного признака №1 делится на два подмножества: (а) и (б-м). Признаки-дубли в этом случае отсутствуют. На второй ступени деления с помощью признака №3 (дублей тоже не имеет) мы получаем подмножества: (з, и) и (б-ж, к-м). Подмножество (з, и) можно разделить на два подмножества ( по одному виду в каждом подмножестве) с помощью нескольких признаков, которые для подмножества (з, и) являются признаками-дублями): №№: 11, 14, 15, 16, 18, 19, 20, 21, 22, 25, 26, 27, 28. Подмножество (б-ж, к-м) с помощью признака № 6 делим на подмножества: (к-м) и (б-ж). Признаки дубли отсутствуют. Теперь подмножество (к-м) с помощью двух признаков-дублей (№ 18 и № 19) делим на подмножества: (к, м) и (л). Подмножество (к, м) легко разделить, пользуясь признаками-дублями №№: 2, 5, 15, 20, 22, 24, 25, 26. Далее делим подмножество (б-ж) с помощью признака № 14 на подмножества: (б-г) и (д-ж). Признаков-дублей в этом случае нет. Для деления подмножества (д-ж) берем признаки-дубли №№: 8, 11 и 25. Получаем подмножества: (ж) и (д, е). Подмножество (д, е) делим с помощью признаков-дублей: №№: 2, 9, 10, 18, 23, 24, 27. Подмножество (б-г) с помощью признаков-дублей №№: 7, 12, 13 делим на подмножества: (г) и (б, в). Последнее подмножество делим с помощью признаков-дублей №№: 24, 27. В итоге мы получили определительный ключ со смешанным (дихотомическим и моноподиальном) делением. В восьми разветвлениях использованы признаки-дубли, что делает эти разветвления вполне надежными. Однако в трех разветвлениях ключа признаков-дублей не существует. Из этих одиночных признаков признак № 1 вполне надежный (макроскопический, прослеживается на протяжении всей вегетации). Признак № 3 менее надежный, а признак № 6 также вполне надежен при определении. |
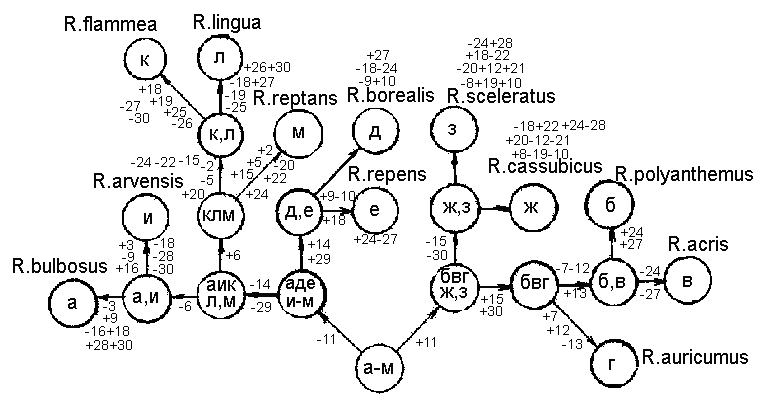 |
Рис. 2г. Графическая модель полного дихотомического ключа для определения видов рода Ranunculus флоры Калининградской области Используя ту же матрицу, можно получить иные схемы деления нашего множества. Так, на рис. 2б мы имеем пример полностью моноподиальной схемы деления, когда на каждой ступени выщепляется сразу один вид. Признаки-дубли здесь возможны только в самом конце графа. В связи с отсутствием признаков-дублей такое деление считать оптимальным нельзя, хотя логически данный граф также непротиворечив. Он более вытянут, чем граф 2а – длина его измеряется 11-тью ступенями или сочленениями (последние в ряду определения виды б и в отстоят от начала графа на 11 разветвлений). Длина графа 2 А составляет всего 6 сочленений и здесь путь распознавания таксонов короче почти в 2 раза. Граф на рис. 2в еще короче (5 сочленений), признаки-дубли здесь отсутствуют только в 2-х разветвлениях. При этом не продублированный признак № 6 вполне надежен так же, как и признак № 11. |
Следовательно, из трех вариантов возможных ключей более оптимален ключ, моделью которого является граф 2В. Именно его мы использовали при словесном оформлении дихотомического ключа для рода. В тех разветвлениях, где признаков-дублей много (больше трех), мы брали в ключ лишь часть из них, а именно те, объединение которых вполне достаточно для четкого разделения подмножества в данном разветвлении.
1. Листья рассеченные или глубоко надрезанные на 5 долей .. 2
- Листья иные ………………………………………………… 6
2. Доли стеблевых листьев узкие (ланцетные или линейные), по краю гладкие ….. 3
- Признаки иные ……………………………………………… 10
3. Прикорневые листья округло-почковидные, в диаметре не более 5 см, доли прикорневых листьев, если имеются, широко-ромбические (узкими не бывают) – R. auricumus L. Сухие луга, обочины дорог, лесные полянки. Часто.
- Прикорневые листья иные, доли их узкие ланцетные или линейные …..4
4. Цветоножки бороздчатые, стебли оттопыренно-волосистые
- R. polyanthemus L. Сухие и нормально увлажненные луга, опушки леса, обочины дорог. Часто.
- Цветоножки не бороздчатые, стебли не оттопыренно- волосистые – R. acris L. Луга всех типов, кроме травяных болот, обочины дорог, опушки лесов, кустарники. Часто.
5. Прикорневые листья округло-почковидные в диаметре около 10 см, чашелистики снизу опушенные, носик плодов хорошо развит, цветоножки бороздчатые, плодики опушенные. – R. cassubicus L. Хвойно-широколиственные и широколиственные леса, кустарники. Часто.
-Все перечисленные признаки иные. – R. sclerathus L.
6. Доли стеблевых листьев широкие явно зубчатые…………….. 7
- Доли стеблевых листьев не широкие, по краю не зубчатые..8
7. Листья тройчато-сложные, плоды голые, цветоножки бороздчатые, стебли не оттопыренно-волосистые. - R. repens L. Cухие луга, поляны, травяные болота, берега осушительных канав, ивняки. Часто.
- Листья не сложные, хотя тройчато-надрезанные, плоды не голые, цветоножки не бороздчатые, стебли оттопыренно- волосистые. – R. borealis Trautv. Хвойные и хвойно- широколиственные леса. Не редко.
8. Все листья цельные ……… 9
- Листья иные ………………… 11
9. Стебли ползучие (распростертые и в узлах укореняющиеся), чашелистиков меньше пяти, носик плодов хорошо развит, цветоножки бороздчатые, чашелистики снизу голые. – R. reptans L. По дну осушительных канав, ручьев, берегам прудов. Не часто.
- Стебли не ползучие, чашелистиков не менее пяти, носик плодов не развит, цветоножки не бороздчатые, чашелистики снизу опушены … 10
10. Плоды голые, цветки мелкие до 15 мм в диаметре, стебли голые, доли стеблевых листьев по краю зубчатые. – R. flammula L. Сухие заболоченные луга, травяные болота, сырые кустарники. Не часто.
- Плоды не голые, цветки крупные более 15 мм в диаметре, стебли оттопыренно-волосистые, доли стеблевых листьев по краю гладкие. – R. lingua L. Сырые луга по берегам рек, ручьев, старых канав. Не часто.
11. Растения однолетние, стебли в основании без клубневидного утолщения, плоды с шипиками, чашелистики во время цветения вниз не отогнутые и к цветоножке не прижатые, доли стеблевых листьев по краю явно гладкие. – R. arvensis L. На полях сорное. Не редко.
- Растения многолетние, стебли в основании с клубневидными утолщениями, плоды без шипиков, чашелистики во время цветения вниз отогнутые, доли стеблевых листьев по краю не гладкие. – R. bulbosus L. Сухие луга на песчаной и супесчаной почве, обочины шоссе. Не редко.
Разумеется, тремя графами возможное количество путей определения на основании исходной матрицы не исчерпывается. Поэтому в дальнейшем потребуется более строгое определение принципов оптимальности деления множества таксонов с тем, чтобы весь процесс можно было выполнять с помощью ПК. Существенно ускорить работу можно и без ПК, используя перфокарты ручной обработки (например, типа К-5 с двумя сотнями пар отверстий краевой перфорации). На каждый вид множества мы заводим перфокарту, пары отверстий нумеруем и каждый признак кодируем одной парой отверстий. При значении признака «да» отверстия выстригаются, призначении признака «нет» краевая перфорация соответствующего номера не простригается. Затем с помощью стальной спицы мы легко и быстро можем делить множество видов по любому признаку в любой последовательности. Признаки-дубли при этом находятся быстро и легко.
Мы считаем, что определители растений должны включать не только ключи (ветвистые графы), но и сами матрицы распределения видов в пространстве элементарных альтернативных морфологических признаков. В этом случае, после процедуры определения, мы можем быстро и легко сравнить определенный нами экземпляр с более полной морфологической характеристикой таксона, приведенной в матрице. При неверном определении можно установить, с каким именно таксоном мы перепутали наш образец. Зачастую, пользуясь региональными определителями, можно не заметить, что имеешь дело с видом, совсем не указанным в данной сводке. Такие экземпляры часто ошибочно подгоняются под один из имеющихся в определителе таксонов.
Матрица избавляет нас от необходимости нагружать определительный ключ кроме диагностических признаков признаками описательными, не являющимися в данном конкретном разветвлении ключа диагностическими. Многие из опубликованных определителей в своих ключах в той или иной степени пытаются совместить функцию распознавания таксонов и функцию их более полного описания, что, конечно, не делает их логически более совершенными.
Таким образом, вся работа по составлению определительных ключей разбивается на несколько последовательных этапов: 1 – определение гербария, изучение литературных источников и составление конспекта видов рода или семейства; 2 – выделение элементарных альтернативных морфологических признаков всесторонне и достаточно полно характеризующих в своей совокупности таксоны данного множества; 3 – установление (по гербарию) характера распределения таксонов в многомерном пространстве элементарных альтернативных морфологических признаков (составление исходной матрицы); 4 – анализ матрицы на предмет отбора удобных при определении диагностических признаков, поиски признаков – дублей для самых первых разветвлений ключа; 5 – построение ветвистых графов-моделей определительных ключей и отбор наиболее оптимальных из них; 6 – составление текста наиболее оптимального ключа.
Предлагаемые здесь ключи и таблицы были использованы при определении растений студентами Калининградского университета на Большом практикуме по ботанике (IV курс) и на полевой практике по ботанике (I и II курсы) в 1985-86 гг. По общему мнению студентов и преподавателей, использовавших эти ключи, они выгодно отличаются от ключей, предлагаемых другими авторами для близлежащих территорий, или террирорий, включающих Калининградскую область.
Дальнейшее развитие метода, как нам представляется, лежит более в строгом определении альтернативных признаков, в стандартизации наборов этих признаков для родов и семейств растений, в составлении баз и банков морфологических и таксономических данных для региональных флор и, в конечном счете, в использовании больших ЭВМ для определения растений. По крайней мере, теоретически и методически компьютеризация процесса определения растений региональной флоры высвечивается вполне определенно. В перспективе морфолого-таксономические базы данных могут сращиваться с базами экологических, ареалогических и ресурсоведческих данных и использоваться во взаимодействии.
Балковский Б. Е. Цифровой политомический ключ для определения растений. Киев: «Наукова думка», 1964, 36 с.
Галанин А. В., Беликович А. В., Проскурина Н. С. Практические работы по биометрии. Выпуск 2. Препринт. Магадан: Издание ИБПС ДВО АН СССР, 1986, 42 с.
Кауфман Н. Московская флора или описание высших растений и ботанико-географический обзор. М., 1889, 761 с.
Маевский П. Ф. Флора средней полосы европейской части СССР. Л.: «Колос», 1964, 880 с.
Определитель высших растений Крыма. Л.: «Наука», 1972, 550с.
Определитель растений Ярославской области. Ярославль: Ярославское книжное издательство. 1961, 450 с.
Победимова Е. Г. Состав, распространение по районам и хозяйственное значение флоры Калининградской области. - В кн.: «Геоботаника», вып. 10, М.-Л.: «Наука», 1956, с. 225 – 329.

|
|
© Беликович А.В., Галанин
А.В. : содержание, идея, верстка, дизайн Все права защищены. 2007 |

