Изучение временных процессов в растительном покрове: перспективы использования графовых моделей. II
А.В. Беликович
Понятие слабых мест в структуре РП
Слабые места – это узлы, связи, группы вершин, которые в наибольшей степени определяют изменения в структуре графа. Проблема слабых мест в основном обсуждалась лишь в работах по изучению технических систем (Войтов и др.,1998), и во всех из них анализ слабых мест связывается с близостью к пределу статической или динамической устойчивости этих систем. В общих чертах проблему слабых мест можно свести к поиску узлов (элементов системы или набору этих элементов), связи которых быстрее всего рвутся при возмущениях внешней среды (увеличении нагрузки). При исследовании РП проблему слабых мест можно сформулировать как поиск классов фитохор или скоплений вершин на графах, для которых при временных перемещениях (или со временем) наиболее вероятен разрыв флористических связей с соседними, наиболее близкими элементами. Вероятно, определение слабого места можно осуществить и путем сравнения структур, выявленных при разной степени антропогенной нагрузки, однако при этом очень сложно найти два полностью аналогичных района.
По-видимому, возможен расчет таких слабых мест и аналитическим путем. Однако постановка такой задачи – дело будущего, когда будет отработан аппарат моделирования ситуаций и состояний РП при разного рода возмущениях среды. Для того, чтобы подойти к этой проблеме, понадобится анализ и систематизация графовых моделей РП с очень многих районов и типов ландшафта, взятых в разных временных интервалах.
Мы же на начальном этапе предлагаем способ нахождения критических (слабых) по статической устойчивости элементов на основе упрощенного метода. В нашем случае слабые места – это те группы вершин на графах, которые со временем поменяли свою позицию. Фактически это узловые ядра флористической структуры, изменение внутри которых можно расценивать как качественные перестройки РП. Причем эти узловые ядра следует отличать от сенсорных элементов – точек наибольшей чувствительности флористического состояния элементарных фитохор.
Так, в разбираемом нами примере слабыми местами являются площади №№ 3,4,6,8,19 и 20, резко поменявшие свою позицию на графе. Заметим, что сенсором (в принимаемом здесь смысле этого слова) из них является лишь площадь № 8. На ней не только сильно поменялся видовой состав, но она и перешла в другой класс за счет идущего на месте гари процесса «отундровения» РП. Таким образом, площадь №8, находящаяся на верхней границе леса, может служить репером для мониторинга за колебаниями этой границы.
Разберем, какие еще фитомезохоры оказались «слабыми местами» флористической структуры РП ландшафта. Площади №3 и 20 поменяли свои позиции из-за смены гидрологического режима. Площадь №3 превратилась из лиственничного леса заболоченного сфагнового в сухой мохово-лишайниковый. На площади №20 (ложбина мерзлоты и ледяных линз под мореной резко усилился поверхностный сток, в связи с чем режим полузастойного избыточного увлажнения сменился проточным режимом, вызвав исчезновение Eriophorum vaginatum и появление множества приручьевых травянистых мезофилов (Seseli condensatum, Epilobium davuricum, Equisetum arvense, Luzula parviflora) и вида-поселенца песчаных наносов Alopecurus aequalis.
Площадь №4, благодаря выпадению из ее РП многих кустарников и специфических видов россыпей, таких как Ribes altissimum, R. nigrum, Cotoneaster melanocarpus, Bergenia crassifolia (по-видимому, в связи с долговременной засухой), потеряла статус самостоятельного класса и подсоединилась к классу кедрово-лиственничных лесов и редколесий. Площадь №19, представляющая фитомезохору моренного озера, выделилась в самостоятельный класс, так как на суглинистых пятнах песчаного берега поселились некоторые виды, нехарактерные для кедрово-лиственничных лесов верхнего лесного пояса. На площади № 6 прошел слабый низовой пожар, затронувший центральную часть исследуемого участка (по лощине), благодаря чему выгорели многие кустарники и кустарнички (Ribes nigrum, Spiraea media) и стали поселяться новые виды, характерные уже для более сухой и каменистой лощины (Berberis sibirica, Rhododendron dahuricum, Carex vanheurckii и др.), а также виды, свойственные производным лесам (Betula platyphylla, Salix caprea).
Таким образом, 6 вершин из 21 в графе поменяли свою кластерную принадлежность, что свидетельствует о действительно крупных изменениях в структуре флористической неоднородности РП данного ландшафта. Причинами этих изменений, вероятно, следует считать усиливающуюся аридизацию климата Центральной Азии, которая приводит к ксерофитизации РП даже высоко в горах. Этот вывод находится вполной противоположности с результатами Б.И. Дулеповой (Дулепова, 2000; Дулепова, Уманская, 2000), доказывающей большую стабильность растительных сообществ Восточного Забайкалья, особенно их флористического состава. Количественный же состав, по данным этих авторов, меняется значительно, особенно в годы метеорологических аномалий. Разница в наших выводах может быть связана с двумя факторами. Во-первых, Б.И. Дулепова исследовала динамику сообществ до 1988 года, то есть, вероятно, до периода наступления интенсивных подвижек в климате, во-вторых, отдельные заложенные ею площадки не образовывали систему реперов, охватывающих весь ландшафт, а были привязаны к степным и лесостепным участкам. Возможно, что фитохоры степного характера как раз и являются самыми стабильными центрами в организации РП забайкальских ландшафтов.
Анализ различных состояний флористической структуры (нескольких временных срезов) помогает оценить устойчивость данной структуры к возмущениям среды и спрогнозировать возможные пути трансформации этой структуры. Благодаря индикационной роли растений, модели ландшафтной флористической структуры РП одного и того же района, взятого в разные промежутки времени, можно использовать для мониторинга состояния биосферы в данном районе. Кроме сенсоров и слабых мест, анализ графовых моделей структуры РП может выявить и такие узлы в структуре РП, которые полностью инвариантны к возмущениям. Их расположение на графовой модели не зависит от типа и силы возмущений, несмотря на то, что видовой состав самих участков может меняться в ту или другую сторону. Такими инвариантными узлами в нашем примере являются классы I (3 площади на нагорных террасах), IV (шахтные отвалы), V (заросли кедрового стланика с лужайками на уступах нагорных террас), подкласс II C (приручьевые кустарниковые кедрово-лиственничные леса) и центральные звенья в классе II – связка из площадей 16,18 (мохово-лишайниковые ерниковые кедрово-лиственничные леса) и связка из площадей 5,7 (кустарничковые кедрово-лиственничные леса с подлеском из кустарников).
Распознавание когерентности флористических элементов
Когерентность в физике и математике – это степень близости реакций или их соотношений на одно и то же возмущение. Аналогично в геоботанике можно считать когерентными те элементы, которые проявляют сходные реакции в процессе развития РП. В силу флористической неоднородности РП ландшафтного района реакции на возмущение его элементов оказываются различными. Однако есть некоторые группы элементов, которые ведут себя более сходно по отношению друг к другу, чем к остальным элементам. При этом возникает возможность агрегирования элементов, то есть их некоторая классификация по отношению к предположительным сценариям возмущения РП. Вероятно, можно в качестве таких когерентных объектов выбирать на графовых моделях кластеры, более мелкие объединения вершин – узлы, или сами вершины.
Так как в качестве примера у нас рассматриваются графы с малым количеством вершин и ребер, то в данном случае, вероятно, удобнее всего выявлять когерентные вершины. Смысл когерентности будет зависеть от того, какие реакции РП мы будем считать сходными. Так как мы исследуем явление флористической неоднородности РП, то основной целью нашего анализа является вопрос о таксономическом аспекте биоразнообразия. Снизился ли уровень этого разнообразия на конкретных участках (альфа-разнообразие РП)? Проанализируем, в каких мезофитохорах снизилось видовое разнообразие, а где оно повысилось.
Для визуального сравнения рядом с каждой вершиной на графе можно проставить стрелки (вверх – повысилось, вниз – снизилось). Заметим, что на площадях 2,3,10,10а,16 и 18 уровень видового разнообразия практически не изменился, и ни одна из этих площадей не является сенсором. Возможно, что такое совпадение не случайно. На площадях 6,7,8,11,12,19,20 уровень видового разнообразия изменился слабо (на 2-4 вида), что может быть объяснено чисто случайными причинами. Зато все остальные площади можно разбить на две группы: где количество видов резко снизилось (4,17,21,24) и где количество видов резко повысилось (1,8,9). В этих двух группах фитомезохоры когерентно реагируют на изменение внешней среды.
В первой группе снижение таксономического разнообразия в фитохорах происходит в связи с иссушением местообитания и выпадением большого числа травянистых мезофилов. То есть когерентность изменений связана с проявлением одного и того же фактора. Во второй группе, напротив, повышение видового разнообразия вызывается на разных фитохорах разными причинами. Так, на площади 1 идет первая стадия процесса бурного зарастания сильно нарушенного участка – появились Rubus sachalinensis, Ribes fragrans, Pentaphylloides fruticosa, возобновление Betula platyphylla, Populus suaveolens, Salix rorida , S. divaricata и др. Причем и в 1984, и в 2000 г. мы имели здесь дело с несформированным РП, поэтому и в дальнейшем следует ожидать повышения видового разнообразия. О начале формирования сомкнутого РП с ценотическими отношениями будет говорить снижение и стабилизация видового разнообразия на площади. На площади 8 параллельно идут два процесса: происходит медленное в подгольцовье зарастание участка после старой гари (появились Angelica decurrens, Aegopodium alpestre, Berberis sibirica) и идет проникновение видов, типичных для тундр – Empetrum nigrum, Carex obtusata, C. scirpoidea, С. stenocarpa, Kobresia myosuroides, Aster serpentimontanus, Schultzia crinita и др. Опять же, здесь трудно говорить о сформировавшемся РП. Что же касается площади 9, то повышение видового разнообразия здесь удивительно, так как на площади идет массовое усыхание кедрового стланика. Анализируя происходящие перестройки в травянистом ярусе, замечаем, что здесь появился ряд видов, свойственных бореальной тайге (Pedicularis labradorica, Carex globularis, Pyrola incarnata, Moehringia lateriflora, Rubus arcticus). Однако, несмотря на значительное число новых видов (9), общий процент изменений не так и высок (27%).
Таким образом, анализируя когерентность реакций разных фитохор на изменения среды, следует понимать, что в основе такой когерентности могут быть разные причины. Тем не менее, выявление когерентных групп может многое дать для распознавания основных направлений динамики РП данного региона. Качественно сходные реакции разных фитохор говорят об однонаправленности возмущений, то есть об определенной векторности изменения окружающей среды.
Различная степень синфазности (когерентности) реакций разных элементов РП на воздействия среды – одно из важнейших проявлений флористической неоднородности РП. Поэтому идентификация групп в большей или меньшей степени когерентных элементов является одним из способов исследования неоднородности РП. Одна из перспектив изучения когерентности в РП – исследование сходства реакций не фитохор, а целых классов фитохор или узлов отношений между фитохорами. Подобные исследования можно делать только на массовом материале с построением многовершинного графа, то есть в ситуации, когда большую роль начинают играть стохастические процессы, и правомерно применение статистических методов. При этом вероятно получение оригинальных данных о различных стратегиях разных типов растительности и разных типов пространственной неоднородности РП при возмущениях среды.
Оценка устойчивости структуры РП к возмущениям
Существует небольшое число публикаций о системно-структурном подходе к исследованию устойчивости экосистем (Крауклис, 1983), и все они сугубо теоретические, не имеют дела с конкретными ситуациями природных состояний среды. Можно сказать, что в наших работах структурный подход применен к изучению неоднородности РП ландшафтного уровня. Основными моментами этого структурного подхода являются определение показателей коэффициента сходства-различия между элементами РП, идентификация на основе этих коэффициентов ординационных кластеров (объединений флористически сходных элементов), и построение графовой ординационной модели РП для последующего детального изучения ее динамических свойств.
Основными проявлениями динамической неоднородности РП следует считать сенсорность и когерентность. Большая или меньшая сенсорность фитохор определяется их положением в разных физико-географических элементах и степенью гомеостаза того типа растительности, который преобладает в фитохоре. Количество и положение слабых мест в ландшафтной флористической структуре РП определяется не только флористическими связями фитохор между собой, но и их разными функциями в задании структуры РП и жизнедеятельности РП ландшафта. Наличие когерентных элементов в структуре РП означает развитие РП в определенном направлении и помогает определить вектор этого развития.
Таким образом, в задачу геоботанического мониторинга должно входить выявление сенсорных элементов, нахождение когерентных узлов и прослеживание направления флористических процессов в данном РП. Рассмотрим пример с мониторингом РП в Сохондинском заповеднике. Наличие большой когерентной группы фитомезохор показывает, что все изменения во флористической неоднородности РП здесь могут объясняться единой причиной. Главный фактор в данном случае – аридизация климата, иссушение местообитаний, большая протайка многолетнемерзлого слоя.
Исходя из предложенных нами определений, следует ожидать, что наибольшая реакция на возмущения внешней среды должна возникать в сенсорных элементах и в тех элементах, которые являются слабыми местами во флористической структуре РП. За этими элементами необходимо регулярное слежение и повышенное внимание со стороны специалистов по мониторингу. Возможно, после проведения предварительного периода общего мониторинга в заповеднике, который позволит выявить наиболее чувствительные к внешним воздействиям элементы (сенсоры) и определить их связи с другими элементами РП, следует перейти к более оптимальной системе мониторинга, когда слежением будет охвачена только сеть сенсоров и слабых мест. Это позволит проводить мониторинг с меньшими затратами и более углубленно.
Рассматривая пример с Сохондинским заповедником, мы видим, что локализация сенсоров на графовых моделях флористической структуры РП практически не зависит от положения «слабых мест». Однако, самую интересную информацию о динамике РП дает расчет сходства графовых моделей флористической структуры РП. Вопрос о том, насколько похожи полученные две структуры, не столь прост. В общем случае расчет сходства двух графов сделан нами ранее (Галанин и др.,1986). В обоих графах по 21 вершины, в первом графе 17 ребер, во втором 18. Девять ребер в обоих графах занимают одинаковое положение.
Максимальное (возможное) число ребер (Max) в 21-вершинном неориентированном графе, не имеющем кратных ребер, равно n(n-1)/2, то есть 420/2=210. При случайном распределении ребер одинаковое положение должны занимать P1·P2·Max ребер (где P = n/Max), то есть 1,5 ребра, что в 6 раз меньше наблюдаемого. Коэффициент сходства двух графов равен:
Pн – Рт
К = -------------- , то есть в нашем случае 0,83
Р
н
Оценка этого сходства по критерию Пирсона показывает, что оно достоверно больше случайного. Действительно, графы весьма сходны, то есть структура флористической неоднородности РП осталась инвариантной, несмотря на то, что половина ребер изменила свое положение.
Нахождение сенсоров и слабых мест - самое необходимое для того, чтобы научиться управлять или контролировать РП на данной территории. При долговременном мониторинге, имея значительную базу данных по изменению флористической неоднородности РП, становится возможен прогноз различных сценариев развития РП. Становится возможен ответ и на вопрос о том, снизилось или повысилось общее разнообразие фитохор в данном районе и неоднородность структуры. Для обоснованного ответа на этот вопрос проведем расчет критерия Пирсона для разницы между наблюдаемым и теоретически ожидаемым количеством вершин с установленным числом исходящих из них ребер. Для графа с 21 вершиной легко определить, каким он должен быть, если ребра распределились между вершинами случайно. Тогда в первом графе вершин, из которых не исходит ни одно ребро, должно быть 4,8; из которых исходит 1 ребро - 8,0; 2 ребра – 6,3; 3 ребра – 3,2; 4 ребра – 0,6; 5 ребер – 0,03. Из наблюдаемого графа можно взять аналогичные наблюдаемые числа. Они довольно резко отличаются от теоретических, то есть наш граф сильно неоднороден. Это хорошо видно на рисунке ниже, где на графиках черными линиями обозначены наблюдаемые распределения вершин с разным количеством исходящих из них ребер, а серыми – теоретически ожидаемые при случайном «разбросе» ребер по вершинам. Величина критерия Пирсона при этом равняется 40,1. Та же процедура, примененная для второго графа, дает другую картину и другой критерий Пирсона, равный 6,3. Это значит, что уровень неоднородности за 16 лет сильно снизился. Граф структуры 2000 г. гораздо более равномерен.
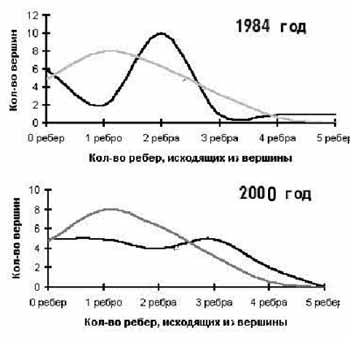 |
Анализ неравномерности графовых моделей флористической структуры РП таежно-гольцового ландшафта Сохондинского заповедника: вверху– в 1984 г., внизу – в 2000 г.
По оси абсцисс - количество ребер, отходящих от вершины; по оси ординат - количество вершин с определенным числом исходящих из них ребер. Сплошная линия - наблюдаемое распределение ребер по вершинам в графе; пунктирная - теоретическое рассчитанное распределение при полной равномерности графа с тем же количеством вершин |
Проведем аналогичную процедуру для другого уровня сходства в 10%, когда количество связей на графах увеличивается до 182 и 178 ребер соответственно. Хотя полученные структуры более неравномерны (критерий Пирсона для первого графа 86,2, а для второго 69,2), тенденция сохраняется: граф 2000 г. более равномерный. Аналогичные расчеты для других уровней сходства (здесь не приводятся) еще раз подтвердили подобный вывод. Заключение может показаться удивительным, но оно обосновано статистическими расчетами: за 16 лет мы видим снижение структурного биоразнообразия РП, то есть утрату неоднородности РП.
Этот вывод перекликается с выводами палеонтологов, установивших, что в периоды катастроф резко снижается таксономическое разнообразие РП (Красилов, 1977; Маркевич, 1995). По-видимому, в периоды катастрофических изменений климата происходит снижение и структурного разнообразия, о чем наглядно свидетельствует пример Сохондинского заповедника.
При сравнении двух пространственно совмещенных моделей РП возникают и вопросы более общего плана, связанные с явлением динамической неоднородности РП. Каким образом можно ее определить? Можно ли считать разброс в величине флористического изменения элементов РП (отдельных фитохор, их классов) и количество когерентных групп степенью динамической неоднородности? Здесь мы вступаем в мало разработанную область формализованного описания динамических процессов в РП. Для постановки таких задач необходимо накопление данных об изменении флористической структуры РП во времени, а для этого необходимо вести длительные мониторинговые исследования на одной и той же системе реперов (участков РП). К сожалению, подобные исследования трудно реализовать из-за быстрой сменяемости поколений исследователей и невозможности существования этой системы вне охраняемых территорий. Единственная возможная база для накопления подобной информации – заповедники. Регулярное переописание пробных площадей, замаркированных на местности, может дать уникальные данные для проведения сравнительного анализа динамических процессов в РП естественных ландшафтов.
