|
|
|
| Теория | Методы | НАШИ АВТОРЫ | Ботаническая жизнь Флора | Растительность | Прикладные вопросы НА ГЛАВНУЮ СТРАНИЦУ | Экспедиции | НАПИШИТЕ ПИСЬМО |
ЗАПАСЫ
ФИТОМАССЫ И ПРОДУКТИВНОСТЬ РАСТИТЕЛЬНОГО ПОКРОВА
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
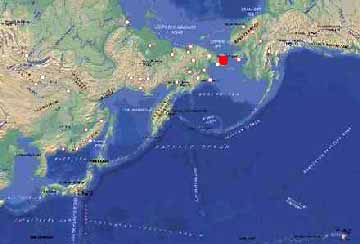
Карта Берингийского сектора Арктики и Субарктики. Красной точкой обозначен район исследования на северо-востоке Чукотского полуострова
Экспедиционные будни на Чукотке. Вездеход основательно застрял. Гусеницы скользят на мерзлоте. Мы несколько часов выбирались из этой трясмны. Болото образовалось на довольно крутом склоне горы северной экспозиции |
ВведениеЗапасы и структура фитомассы являются наиболее существенными экологическими характеристиками растительного покрова, как в пределах отдельных экосистем, так и в пределах целых ландшафтных районов и природных зон. Продукционный процесс рассматривается нами как часть биологического круговорота вещества, поэтому, изучая структуру фитомассы в конкретных растительных сообществах и микроценозах , широко представленных в растительном покрове района, мы, таким образом, оцениваем важные параметры биологического круговорота Галанин и др., 1995). Следует отметить, что как природное явление биологический круговорот, открытый В.И. Вернадским еще в начале ХХ века (Вернадский, 1978), и в наши дни остается недостаточно изученным. Ранее нами (Галанин, 1989,1991) были рассмотрены некоторые возможности оценки биологического круговорота через объем и структуру запаса вещества в локальной экосистеме, скорость обращения вещества в круговороте и степень замкнутости биологического круговорота в локальной экосистеме для элементов минерального питания. Под локальной экосистемой мы понимаем здесь экосистему в пределах конкретной физико-географической фации – одного контурфитоценоза. Запасы фитомассы и продуктивность растительного покрова в зоне тундры изучены явно недостаточно, особенно это справедливо для Северо-Востока Азии, где стационарные геоботанические исследования, начатые в конце 80-х годов, так и не смогли развернуться из-за общего кризиса в России, начавшегося в 90-е годы. В Чукотском автономном округе продуктивность растительного покрова изучали почвоведы И.В. Игнатенко, Б.А. Павлов, А.А. Пугачев, А.Ш. Оганесян, Н.Ю. Смородникова (Игнатенко и др., 1976; Игнатенко, Пугачев, 1979; Пугачев, 1983), однако их исследованиями были затронуты только тундры острова Врангеля, и района Чаунской губы. Из геоботаников продуктивность растительного покрова на юге Магаданской области изучала Т.А. Москалюк ( Москалюк , 1979, 1988, 1990). |
|
На Аляске изучением продуктивности растительного покрова занимались ботаники, почвоведы и экологи. Результаты этих работ обобщены в виде коллективной монографии «Vegetation and Production Ecology of an Alaskan Tundra» (1978). Однако и на Аляске ландшафтное и экологическое разнообразие таких исследований невелико, чаще всего работы проводились в лесных экосистемах внутренней и южной Аляски. В июле-августе 1991 г. авторы участвовали в работе комплексной международной экспедиции Института биологических проблем Севера ДВО РАН на северо-востоке Чукотского п-ова. Одной из задач экспедиции было изучение запаса биомассы в разных типах тундры будущего Берингийского международного национального парка. В связи с тем, что традиционными методами изучения запаса биомассы в столь краткий период времени собрать материал для характеристики целого ландшафтного района в бассейне р. Чегитунь было невозможно, мы были вынуждены использовать нетрадиционную методику сбора фактического материала. На Аляске изучением продуктивности растительного покрова занимались ботаники,почвоведы и экологи. Результаты этих работ обобщены в виде коллективной монографии «Vegetation and Production Ecology of Alaskan Tundra » (1978). Однако и на Аляске ландшафтное и экологическое разнообразие таких исследований невелико, чаще всего работы проводились в лесных экосистемах внутренней и южной Аляски. |
|
Дриадовая тундра на щебнистом склоне. Здесь встречается довольно много лишайников и травянистых растений. Растительный покров куртинный и пятнистый
Пятнистые и куртинные дриадово-лишайниково-разнотравные тундры - обычный тип растительного покрова на гребнях чукотских гор
Типичный ландшафт Восточной Чукотки. Подзона типичных горных тундр | МетодикаРастительный покров на ценотическом уровне организации хорологически неоднороден и представлен ценоячейками или микрогруппировками, которые, сочетаясь друг с другом, и образуют фитоценотическую «ткань» растительного покрова урочищ, местностей и целых ландшафтов (Галанин, 1989, 1991). В связи с этим, метод случайно заложенных мелких учетных площадок, разбросанных в пределах одного контурфитоценоза без учета тонкой ценотической неоднородности растительного покрова, требует очень большой повторности. Вместе с тем, опытный геоботаник в полевых условиях легко различает разные ценоячейки и может закладывать учетные площадки, ориентируя каждую из них на определенный тип ценоэлемента . Зная относительную площадь, занятую в контурфитоценозе ценоячейками разного типа, мы легко можем экстраполировать полученные данные на всю площадь контурфитоценоза , а затем на конкретные типы тундры и, имея геоботаническую карту, на весь ландшафтный район. Разумеется, точность такой оценки запаса биомассы будет далеко не стопроцентной, но она будет не намного ниже точности при рандомизированном распределении учетных площадок при невысокой повторности. Изучая структуру растительного покрова района среднего течения р. Чегитунь , мы пришли к выводу о том, что примерно 80-85% растительного покрова ландшафтного района образованы 35-40 типами ценоячеек . При этом некоторые контурфитоценозы , например, заросли березки тощей ( Betula exilis ) на бровках надпойменных террас, могут складываться из ценоячеек одного типа, другие, например кочкарная пушицевая ( Eriophorum vaginatum ) тундра, состоят из двух типов ценоячеек . Встречаются контурфитоценозы и микрокомбинации , в которых можно выделить три и даже четыре типа ценоячеек , например микрокомбинация грядово-мочажинного комплекса. Запас надземной и подземной фитомассы в бассейне среднего течения р. Чегитунь оценивался в течение 10 дней в первой декаде августа на квадратных учетных площадках размером 25х25см. Каждая такая площадка закладывалась в пределах одной ценоячейки в однократной повторности для каждого типа ценоячеек . При этом на основе визуальных наблюдений выбирался наиболее типичный вариант каждого типа ценоячеек . В связи с этим наши данные не имеют статистических погрешностей и могут рассматриваться только как первое приближение к оценке запаса фитомассы и ее структуры в данном регионе. При выделении ценоячеек в природе мы использовали доминантно-эдификаторный подход. При этом были отобраны те виды доминантов-эдификаторов, которые в растительном покрове данного района встречаются часто и их ценоячейками в общей сложности в ландшафте занято более 80% территории. За центр конкретного ценоэлемента часто принималась особь вида эдификатора , образующая свое мощное фитогенное поле, иногда единое фитогенное поле полидоминантного характера формировала группа особей разных видов. В любом случае учетная площадка размером 25х25см для отбора фитомассы закладывалась в самой типичной части ценоэлемента . С площадок собиралась вся надземная фитомасса , которая сразу же разбиралась по видам и фракциям: живая (трава, листья кустарников и кустарничков, древесина) и мертвая ( мортмасса ). В некоторых случаях при характеристике ценоэлемента кустарниковых ив мы использовали учетные площадки размером 100х100см для сбора надземной фитомассы и площадки размером 25х25см для учета подземной фитомассы. |
|
Подземная фитомасса отбиралась вместе с почвой на тех же учетных площадках по горизонтам: 0-5см, 5-10см, 10-15см, 15-20см и даже 20-25см там, где почва за сезон оттаивала достаточно глубоко. Глубже 25см в почве даже в самых высокопродуктивных типах тундры в нашем районе сосредоточено меньше 5% от общего запаса подземной фитомассы . В отдельных случаях образцы для учета подземной фитомассы отбирались строго по почвенным горизонтам. Подземная фитомасса от почвы отмывалась в полевых условиях на почвенных ситах с самым малым диаметром ячей. При этом терялись только самые мелкие фракции мертвого органического вещества - уже явного торфа. Потери фитомассы при промывке составляли не более 2-3%, что вполне соизмеримо с искажениями, получаемыми из-за не всегда точно отбираемых колонок почвы. В поле образцы надземной и подземной фитомассы высушивались до воздушно сухого состояния, упаковывались в матерчатые мешочки и транспортировались в лабораторию. Здесь они высушивались до абсолютно сухого веса в термостате в течение суток при температуре +1050 С и взвешивались на весах ВЛТК-500. Разбор подземной фитомассы на живую и мертвую не производился. РезультатыНиже в виде очерков будут приведены фактические материалы по каждому проанализированному нами ценоэлементу. Наряду с данными о запасе и структуре фитомассы приводятся сведения о таксономическом составе ценоэлементов и характеристика местоположений, из которых брались образцы для анализа. Ценоэлемент ивы полярной ( Salix polaris ) характерен для нивальных местообитаний, расположенных в ложе очень поздно стаивающих снежников, обычно к концу июля – началу августа. Площадка заложена на берегу термокарстового озера, в нижней части крутого склона юго-западной экспозиции. Почва супесчаная, с большим количеством мелкой гальки, интенсивно гумусированная на глубину 10-15см. Увлажнение средней интенсивности, но во время таяния снежника избыточное. Нивальный режим выражен очень сильно. Обилие видов здесь и далее приведено в трехбалльной шкале и оценивалось визуально. Структура запаса фитомассы в граммах абсолютно сухого вещества на площадку 25х25см следующая: Живые листья и веточки ивы полярной 4,4 Из полученных данных видно, что общий запас надземной фитомассы в этом типе ценоячеек очень низкий, однако практически половина фитомассы приходится здесь на долю компонент с быстрой ротацией вещества в биологическом круговороте. Судя по количеству мортмассы , полный оборот вещества здесь происходит за 1-2 года. Вполне возможно, что значительная часть листьев ивы выедается зайцами и другими животными, что сильно ускоряет биологический круговорот. При низкой продуктивности и небольшом ежегодном накоплении вещества в фитомассе скорость биологического круговорота в этом типе ценоячеек весьма значительная. Запас подземной фитомассы здесь незначительный, чуть меньше наземного. Ценоэлемент ивы сетчатой ( Salix reticulata ) приурочен к полунивальным луговинам и высоким ивнякам из ив шерстистой ( S . lanata ) и красивой ( S. pulchra ). На участках, имеющих незначительное укрытие снегом зимой, ива сетчатая в данном районе не встречается, но и в настоящих нивальных тундрах этот вид крайне редок. Площадка заложена в описании N 3 на берегу небольшого ручья, протекающему по склону южной экспозиции крутизной 15-20о в довольно узкую долину ручья. В этом месте ручей подрезает шлейф отрога хр. Иссэтэн . Увлажнение в данном местообитании нормальное, нивальный режим средней интенсивности, почва щебнисто-суглинистая интенсивно гумусированная без следов оглеения , протаивающая на значительную глубину, в связи с чем дренаж в этом месте интенсивный. Структура запаса фитомассы следующая: Живые листья и сережки ивы сетчатой 8,9 Общий запас надземной фитомассы в этом ценоэлементе в 5 раз больше, чем в предыдущем, однако фитомассы с быстрой ротацией здесь больше только в 2 раза. Судя по соотношению запаса живой фитомассы с быстрой ротацией и запаса мортмассы скорость биологического круговорота здесь примерно в 2 раза ниже, чем в ценоячейке ивы полярной. Полный оборот вещества здесь происходит примерно за 3-4 года. Полный же оборот вещества через древесину длится значительно дольше, по всей вероятности, за 15-20 лет. Снижение скорости круговорота в данной ценоячейке , по сравнению с предыдущей, вполне компенсируется количеством вещества в круговороте так, что в целом мощность потока вещества в круговороте в ценоэлементе ивы сетчатой несколько больше, чем в ценоэлементе ивы полярной. Разумеется, это предварительное предположение нуждается в проверке на более массовом фактическом материале. Запас подземной фитомассы в данной ценоячейке превышает запас надземной фитомассы более чем в 3 раза. Последнее обстоятельство, несомненно, связано с хорошей приспособленностью данного типа растительности к суровым климатическим условиям Арктики, и в первую очередь с дефицитом тепла. Живое вещество в почве более защищено от холода, чем в наземном слое, но из-за многолетней мерзлоты 95% подземной фитомассы сосредоточено в верхнем 10-ти сантиметровом слое. Ценоэлемент иван-чая широколистного (Chamerion latifolium) встречается на галечниках в поймах рек и ручьев с интенсивным аллювиальным режимом. Часто аллювиальные процессы накладываются на процессы нивации , так как узкие долины рек и ручьев зимой забиваются снегом. Учетная площадка заложена в пределах еоботанического описания N 3, но значительно ниже предыдущей, уже в пойме ручья. Почва здесь не сформирована, увлажнение периодически избыточное проточное. Структура запаса фитомассы данной ценоячейки следующая: Живая трава иван-чая широколистного 14,2 Общий запас надземной фитомассы в этой ценоячейке чуть больше, чем в ценоячейке ивы сетчатой, а вот запасы подземной фитомассы практически не отличаются. Живой фитомассы с быстрой ротацией в ценоэлементе иван-чая широколистного примерно в 4,3 раза меньше, чем мортмассы . Следовательно, полный оборот вещества здесь происходит за 4-5 лет, то есть чуть дольше, чем в ценоэлементе ивы сетчатой. Количество вещества в круговороте здесь несколько больше, но скорость оборота чуть меньше, следовательно, мощность потоков вещества в ценоячейках ивы сетчатой и иван-чая широколистного примерно равная. Сучья ивы шерстистой к данной ценоячейке вроде бы не относятся, но вещество, высвобождаемое при их разложении, включается в новый цикл иван-чаем и его спутниками по ценоячейке . Ценоэлемент куропаточьей травы цельнолистной ( Dryas integrifolia) представляет собой дриадовые мохово-лишайниковые подушки на сухих щебнистых склонах карбонатных гор хр. Иссэтэн. Площадка заложена в описании N 4 в щебнистой редкокуртинной дриадовой тундре в средней части склона южной экспозиции крутизной 20о. Почва здесь скелетная карбонатная слабо гумусированная под незадернованной щебенкой, но довольно интенсивно гумусированная под подушками дриады. Увлажнение недостаточное, снег зимой интенсивно сдувается. Выявлена следующая структура фитомассы : Живые ветви и листья дриады узколистной 20,4 Надземная фитомасса в этой ценоячейке превышает подземную примерно в 2 раза. Большая часть надземной фитомассы приходится на компоненты с очень медленной ротацией вещества, а вот фитомассы с быстрой ротацией здесь не больше 5г, даже если к ней отнести многолетние листочки дриады. Примерно такой запас быстро ротируемой живой фитомассы в ценоячейке крошечной ивы полярной. Здесь мало мортмассы , но это легко объяснить молодостью куртины и очень скудным ежегодным отпадом вещества. Складом медленно ротируемого вещества в данной ценоячейке является не мортмасса, а многолетние ветви и листья дриады. В старых отмирающих куртинах дриады структура запаса фитомассы следующая: Живая фитомассы (листья дриады, лишайники) 4,3 Из полученных данных можно заключить, что мощность потока вещества в биологическом круговороте в пятнистой дриадовой тундре на сухих щебнистых карбонатных склонах крайне незначительная, ниже даже, чем в нивальной тундре с доминированием ивы полярной. Ценоэлемент хвоща полевого (Equisetum aarvense) и зеленых мхов характерен для шлейфов склонов сопок, конусов выноса и обширных надпойменных террас и встречается в слабо выраженных депрессиях с периодически избыточным, а периодически нормальным увлажнением. Нивальный режим в таких местах не выражен, снег в значительном количестве не скапливается. Учетная площадка заложена в геоботаническом описании N 5 на пологом шлейфе склона известковой горы южной экспозиции с уклоном 2-3о. Почва суглинистая с примесью карбонатного щебня. Выявлена следующая структура запаса фитомассы : Живые веточки и побеги хвоща 4,6 Запас подземной фитомассы здесь примерно в 5 раз больше, чем надземной. Жизнь здесь пряталась в почве. Значительная часть надземной фитомассы сосредоточена во фракции с быстрой ротацией вещества, масса опада только в 2 раза превышает запас живой фитомассы с быстрой ротацией, поэтому можно предположить, что скорость оборота вещества в этой фракции весьма высокая – примерно 2-3 года. Зеленые мхи в круговороте вещества здесь не играют большой роли, так как запас их фитомассы невелик. Но, обладая большой гигроскопичностью и удерживая много воды, мхи в биологическом круговороте играю роль своеобразного стабилизатора водного и температурного режимов. Задерживая в своих телах воду, мхи тем самым препятствуют выносу подвижных атомов в почве за пределы локальной экосистемы. Ценоэлемент осоки ( Carex lugens) и пушицы ( Eriophorum vaginatum) распространен на шлейфах склонов гор и конусах выноса. В растительном покрове этот тип ценоячеек закономерно сочетается с предществующим типом и вместе в сочетании они образуют контурфитоценозы , представленные геоботаническим описанием N 5. В отличие от предыдущего типа ценоячеек в данном отчетливо выражено застойное переувлажнение, поэтому почва здесь со следами оглеения . Выявлена следующая структура запаса фитомассы: Живая трава осоки и пушицы 9,2 Надземная фитомасса в данной ценоячейке значительная, но все же составляет только 1/3 от запаса подземной фитомассы . Жизнь и здесь как бы стремится укрыться в почве, где перепады температуры и влажности в течение суток и в течение года значительно меньше, чем на поверхности. Не смотря на большой общий запас фитомассы с быстрой ротацией здесь немного, примерно столько же, как в предыдущей ценоячейке . Незначительное количество надземной мортмассы позволяет говорить о довольно быстром обороте вещества в этой ценоячейке . По всей вероятности живая надземная фитомасса здесь интенсивно выедается травоядными животными. Примечательно, что фитомасса мхов в этой ценоячейке в 7 раз больше фитомассы травянистых растений, следовательно их стабилизирующая роль в круговороте вещества в этой ноячейке больше, чем в ценоячейке хвоща полевого. Сравнивая осоково-пушицевый и хвощовый ценоэлементы одного контурфитоценоза следует заметить, что количество вещества и структура запаса фитомассы в них сильно различаются. На наш взгляд такое хорологическое разнообразие биогеохимического процесса в пределах одного контурфитоценоза oптимизирует его и делает более стабильным. Ценоэлемент багульника стелющегося (Ledum decumbens ) распространен в сырых кустарничково-моховых бугорковатых тундрах на высоких надпойменных и обширных нагорных террасах, на пологих слонах увалов и шлейфах склонов гор. Бугры мерзлотного пучения диаметром 1-2м и высотой до 30-40см на таких участках перемежаются довольно глубокими трещинами. Нивальный режим не выражен, напротив, снег сдувается с бугров, оставаясь только в трещинах. Почва чаще всего супесчаная, или легко суглинистая, сильно оторфованная , в нижней части почвенного профиля довольно много гальки и щебня. Учетная площадка заложена в пределах геоботанического описания N 6. Выявлена следующая структура запаса фитомассы : Живые листья ивы красивой 4,4Живые листья ивы голубой 0,9 Живая трава осок и злаков 1,1 Живая трава разнотравья 2,1 Итого живой фитомассы с быстрой ротацией 8,5 Живые веточки багульника с листьями 16,1 Живые ветви ивы красивой 5,3 Живые ветви ивы голубой 3,2 Живые ветви брусники с листьями 3,7 Итого живой фитомассы с медленной ротацией 28,3 Мортмасса 74,4 Итого надземной фитомассы 111,2 Всего подземной фитомассы 163,8 в том числе по горизонтам: 0-5см - 127,0 5-10см - 36,8 Общий запас фитомассы 311,8 На долю фитомассы с быстрой ротацией в этой ценоячейке приходится только 1/11 часть всей надземной фитомассы , - значительно меньше, чем в хвощевой и осоково-пушицевой ценоячейках . В данном случае мы имеем дело с экосистемой с медленным биологическим круговоротом. Несмотря на то, что живая надземная фитомасса здесь невелика, оборот и этого вещества происходит за 3-4 года – примерно во столько раз запас живой фитомассы меньше запаса мортмассы . Большое количество вещества и в этом ценоэлементе сосредоточено в почве. Следует заметить, что данный ценоэлемент является типично тундровым и очень широко распространен в ландшафте. Ценоэлемент брусники малой (Rhodococcum vitis - idaea ) в нашем районе – это один из ценоэлементов плакорной тундры, в которой приурочен к выпуклым участкам – буграм пучения. Почва, снеговой и водный режимы в этом ценоэлементе примерно такие же, как в предыдущем, разве что здесь несколько суше. Учетная площадка заложена в пределах геоботанического описания N 6. Выявлена следующая структура запаса фитомассы : Живые листья ивы сетчатой 1,7 0-15 - 163,9 В этом ценоэлементе доля живого вещества с быстрой ротацией очень незначительна и биологический круговорот очень замедленный . Полный оборот вещества даже через фракцию фитомассы с быстрой ротацией здесь осуществляется примерно за 15-20 лет. Не смотря на то, что общий запас фитомассы в этом ценоэлементе весьма значительный (в пересчете на 1 га около 65,344 т), интенсивность потока вещества в биологическом круговороте здесь крайне низкая. Несложный расчет показывает, что в данном типе ценоэлемента мощность потока вещества через биологический круговорот составляет всего лишь около 25,5 кг-год /га. Основная биогеохимическая активность в такой тундре имеет место не на поверхности, а в глубине почвы. Ценоэлемент ивы красивой (Salix pulchra ) как и два предыдущих, характерен для плакорной бугорковатой кустарничково-мохово-лишайниковой тундры (геоботаническое описание N 6), но приурочен к наиболее дренированным участкам. Усиленный дренаж в таких местах связан с роющей деятельностью суслика длиннохвостого. Почва здесь протаивает глубже, увлажнение нормальное. Трофность в данном местообитании заметно выше, также за счет суслика. Кустики ивы красивой высотой до 0,7м приурочены к трещинам. Структура запаса фитомассы выглядит следующим образом: Живые листья ивы красивой 30,8 В этом ценоэлементе запас надземной фитомассы выше, чем подземной, что характеризует кустарниковую тундру, а не кустарничковую. Таким образом, в пределах одного контурфитоценоза (геоботаническое описание N 6) мы наблюдаем сочетание арктических и гипоарктических ценоячеек, сильно различающихся по типу биологического круговорота. Примерно 30г вещества, накапливаемых за год ивой красивой, ежегодно в этом ценоэлементе поступают на поверхность почвы. Примерно столько же мертвых листьев этого вида находится в опаде . Следовательно, коэффициент ротации вещества через листья этого вида здесь равен единице. Это значит, что разлагается за год столько же листьев ивы красивой, сколько их осенью поступает в опад . Через древесину, мхи и лишайники ротация вещества происходит значительно (в 20-30 раз) медленнее, чем через листья кустарников, кустарничков и траву. |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Ивняки из Salix alaxtysis на галечнике в пойме реки. На восточной Чукотке это самые высокие кустарники. Иногда они достигают высоты 1,5 м |
ЛитератураАндреяшкина Н.И. К методике определения годичного прироста, опада и скорости разложения в сообществах гипоарктических кустарников и кустарничков // Бот . ж урн., 1987. Т. 74, N 4. С. 530-541. Андреяшкина Н.И. Влияние тяжелого гусеничного транспорта на рстительный покров подзоны типичных тундр // Экология, 1997. N 2 . С. 143-145. Игнатенко И.В., Оганесян А.Ш. Растительная масса некоторых типов тундры острова Врангеля // Бот . ж урн., Т. 72. 12. С. 1636-1642. Галанин А.В. Ценотическая организация растительного покрова. Владивосток: ДВО АН СССР, 1989. 162с. Галанин А.В. Флора и ландшафтно-экологическая структура растительного покрова. Владивосток: ДВО АН СССР, 1991. 272 с. Вернадский В.И. Живое вещество. М.: Наука, 1978. 289 с. Игнатенко И.В., Котляров И.И., Нестеренко А.И., Пугачев А.А. Запас и структура общего органического вещества в кедровостланниковых зарослях Охотского побережья // Биология и продуктивность растительного покрова Северо-Востока . Владивосток: СВНЦ ДВО АН СССР, 1976. С. 138-158. |
Игнатенко И.В., Пугачев А.А. Динамика растительной массы и биологический круговорот в горнотундровых и кедровостланиковых ландшафтах Северного Охотоморья // Биологический круговорот в тундролесьях юга Магаданской области. Владивосток: ДВНЦ АН СССР, 1979. С. 92-124.
Москалюк Т.А. Запасы и структура растительной массы в основных типах лиственичников Северного Охотоморья // Биологический круговорот в тундролесьях юга Магаданской области. Владивосток: ДВНЦ АН СССР, 1979. С. 16-27.
Москалюк Т.А. Структура и продуктивность лесов Северного Охотоморья . Владивосток: ДВО АН СССР, 1988. 144с.
Москалюк Т.А. Возрастное развитие и биологическая продуктивность чозениевых лесов на юге Магаданской области // Лесоведение. 1990. С. 46-55.
Пугачев А.А. Запасы и структура фитомассы зарослей кедрового стланика бассейна Верхней Колымы // География и природные ресурсы, 1983. N 2. С . 61-71.
Vegetation and Production Ecology of an Alaskan Tundra (1978).|
|
|
© Беликович А.В., Галанин
А.В. : содержание, идея, верстка, дизайн Все права защищены. Сайт создан весной 2004 г.. |