Варианты реализации онтогенетической траектории
дуба черешчатого (Quercus robur L.) в условиях Самарской областиА.В. Иванова, М.Т. Мазуренко © 2012
The ontogeny of specific plant species has its own peculiar ontogenetic trajectory. Deviations from the ontogenetic trajectories are caused by changes in habitat conditions. The polymorphism of ontogeny is connected with habitat changes. When plant organism has deviations, it tends to maintain and reproduce pattern and characteristics of its own ontogenetic trajectories. Deviations and tendencies of maintaining of ontogenetic trajectories were studied on the example of oak (Quercus robur L.) on leewage slopes of Samara Region, Russia. It is assumed that when extreme factors are prolonged, the plants can consolidate genetically deviating ontogenetic trajectory.
Введение
В течение большого жизненного цикла (онтогенеза) определенный вид растений проходит ряд этапов развития, возрастных состояний, этапов по определенному вектору развития. Обладает свойственной виду онтогенетической траекторией. Отклонения от онтогенетической траектории вызываются изменением условий обитаний. С этим связан полиморфизм онтогенеза. При отклонениях растительный организм стремится к восстановлению главной онтогенетической траектории. Отклонения и стремление к восстановлению онтогенетической траектории исследованы на примере дуба обыкновенного (Quercus robur L.) в Самарской области на обдуваемых склонах. Предполагается, что при длительном воздействии экстремального фактора растения могут закрепить наследственно отклоняющуюся онтогенетическую траекторию.
Большой жизненный цикл растения (онтогенез) от прорастания семени до его отмирания - претерпевает ряд кардинальных перестроек - возрастных состояний (Уранов, 1967). Возрастные состояния растений, их положение в ценопопуляциях, исследованы на очень многих объектах. В настоящее время на этот счет имеется большая и уже труднообозримая литература (Жукова, 1983, 1995, 2001; Смирнова и др., 1976 и мн. др). Возрастные изменения отражаются на внешнем облике растений. И. Г. Серебряковым (1962) они названы фазами онтогенеза. При смене одной фазы онтогенеза последующей – меняется биоморфа растений. Особенно наглядно это выражено у древесных растений, в частности у деревьев (Заугольнова, 1968; Вахромеева 1975; Полтинкина, 1985; Смирнова, Чистякова, 1980, 1982 и др.) - наиболее крупных биоморф, завоевывающих в процессе онтогенеза большое воздушное пространство. У деревьев по мере нарастания, ветвления структура дерева с возрастом усложняется и соответственно этому меняется его жизненная форма названная нами онтобиоморфой (Мазуренко, 1980,1986). Ряд последовательно сменяющихся в онтогенезе онтобиоморф определенного вида имеют свой вектор развития, свою онтогенетическую траекторию (Белоусов, 1983; Мазуренко, 1987).
Онтогенетическая траектория наследственно закреплена и обладает определенной устойчивостью, характеризующей онтогенез определенного вида в оптимальных для него условиях обитания. Однако у разных видов степень жесткости этого закрепления (норма реакции) может быть различной. У многих видов она может изменяться в значительной степени в зависимости от условий окружающей среды. Особенно это выражено в экстремальных условиях. Например на границах ареала. Онтогенетическая траектория может отклоняться при различной степени затенения, под влиянием холода, в засушливых условиях при дефиците воды, особенно обдуваемости ветрами и т.д. (Мазуренко, 1976, 1978, 1986 и др.).
В этом случае растение, отклоняясь от свойственной виду траектории на разных стадиях онтогенеза, особенно в молодом возрасте, может приобретать совершенно иную биоморфу, не свойственную для типичной онтогенетической траектории вида. Устойчивость нарушается. Отклонение выражается тем сильнее, чем сильнее иные, по сравнению с типичными, оптимальными, условия обитания вида. С явлением отклонения от главной онтогенетической траектории связана поливариантность онтогенеза. Описываются многочисленные варианты протекания онтогенеза у одного и того же вида. Многочисленные варианты связаны с изменением экологических условий, особенно на границах ареала вида, давлением фитоценоза.
Многочисленные работы по исследованию возрастных состояний определенных видов в ценопопуляциях посвящены именно поливариантности онтогенеза разнообразных жизненных форм растений (Махатков, 1991; Жукова 2001, Савиных, 2004 и мн. др). Эта поливариантность в определенном онтогенетическом состоянии отражает степень колебания, отклонения в сторону от онтогенетической траектории и амплитуду этого колебания, встраивание в новые для вида экологические (географические) условия.При отклонении от главного вектора - онтогенетической траектории, растение на определенных стадиях жизненного цикла может быть представлено совершенно иными биоморфами, нежели в обычных для него условиях произрастания. Отклонения не закреплены наследственно. И при описании вариантов всегда ссылаются на основной путь его течения, основной вектор – его главную онтогенетическую траекторию. Отклонения вызваны иными условиями.
Объект исследования
Дуб черешчатый (Quercus robur). Фото с сайта: http://www.ecmo.ruПри вынужденных отклонениях растение рано или поздно стремится восстановить свой первоначальный облик – вернуться к своей типичной для вида траектории. Эта траектория отражена в его архитектурной модели, наследственно закрепленной. А отклонения не закреплены наследственно. Отклонениям от главного вектора развития, поливариантности онтогенеза уделялось и продолжает уделяться большое значение. А обратный процесс – стремление к восстановлению остается пока не изученным. Отклонения показывают степень пластичности вида, в частности его жизненной формы.
Мы поставили перед собой задачу продемонстрировать стремление к возврату главной для вида, онтогенетической траектории при ее отклонениях. Объектом исследования нами избран дуб черешчатый (обыкновенный) (Quercus robur L.) произрастающий в Самарской области.Дуб обыкновенный распространен почти во всей Европе от Пиренейского полуострова до Урала. Северная граница его ареала находится в Скандинавии от 63° с. ш. на западе, до 60° с. ш. на востоке. На севере Русской равнины она постепенно смещается к югу, проходя через истоки реки Сухоны и верховья реки Уфы, вдоль Урала направляется к Аралу. Южная граница ареала дуба проходит с запада на восток через Кишинев, Кривой Рог, Днепропетровск, устье реки Северный Донец, южнее Волгограда, а затем идет к Саратову. Дизъюнкции имеются в Крыму и на Кавказе.
Дуб обыкновенный один из основных видов зоны широколиственных лесов на Восточно-Европейской равнине. Он широко распространен в подзоне хвойно-широколиственных лесов. Становится редким в подзоне южной тайги, где растет почти исключительно по долинам крупных рек. В степной зоне он также растет по долинам рек, днищам и склонам балок, образуя так называемые долинные и балочные или байрачные леса. П. Л. Горчаковский, изучая восточную границу распространения дуба обыкновенного, отмечает, что крайне северные местонахождения его связаны с относительно повышенными, хорошо прогреваемыми местами, где нет застоя холодных масс воздуха. Напротив, на юге дуб переходит в лучше увлажненные тенистые долины оврагов (Горчаковский, 1968).
Дуб черешчатый (Quercus robur)
Фото с сайта: http://www.dal.hubpages.comВ Московской области дуб черешчатый является деревом первой величины, достигающим 45 м высоты и до 2 м в диаметре (Рысин, Рысина, 1990). В условиях Самарской области, расположенной вблизи южной границы его ареала, дуб в большинстве случаев крупное одноствольное дерево до 17-18 м в высоту. Очевидно, приближение южной границы ареала сказывается на общей высоте растений. В. В. Благовещенский, проводя исследования на территории Приволжской возвышенности, отмечает высоту деревьев дуба 7-20 м (Благовещенский, 2005). Видно, что разбег по высоте наблюдаемых растений очень большой. Высотный минимум характерен для остепненных дубрав, которые содержат в травяном и кустарниковом ярусе степные элементы, соседствуют со степными участками. Более высокие деревья находятся в составе сложных дубовых лесов на глинистых субстратах в условиях сравнительной близости грунтовых вод (Благовещенский, 2005).
Дуб обыкновенный (черешчатый) наряду с другими древесными породами: липа (Tilia cordataMill.), клен платановидный (Acer platanoidesL.), осина (Populus tremulaL.), береза (Betula verrucosa Ehrh.) образует основу лесного фонда Самарской области. Между тем, площадь дубовых лесов на территории области сокращается, их все чаще сменяют вторичные сообщества – липняки и кленовники. Состояние коренных дубрав в основном признается как неудовлетворительное (Матвеев, Матвеева, Соловьева, 2009). Причинами такого состояния называются увеличение рекреационной нагрузки, сильные морозы, подтопление прибрежных лесов, вызванное изменением гидрологического режима при строительстве Саратовского водохранилища, ошибки ведения лесного хозяйства, провоцирующее размножение вредителей и болезней. Все эти факторы снижают возможность естественного возобновления дуба, и площади дубрав сокращаются. В связи с вышеназванными причинами появилось предложение о внесении дуба обыкновенного в Красную книгу Самарской области (Матвеев, Матвеева, Соловьева, 2009). Учитывая, что негативные процессы, происходящие в дубовых насаждениях, продолжают нарастать, а ученые лишь констатируют данные факты, это предложение не лишено смысла и вполне обосновано. Дуб обыкновенный является важнейшей лесообразующей и хозяйственнозначимой породой, заслуживающей пристального внимания и всестороннего изучения.
Наши исследования проводились в 2010-2011 годах на территории Самарской области в Жигулевском заповеднике (Предволжье), а также Красноярском и Сергиевском административных районах (Заволжье). При описании биоморф дуба мы пользуемся методикой разработанной нами для древесных растений: деревьев, кустарников (Мазуренко, Хохряков 1977, Мазуренко, 1980, 1986).
На территории Самарской области, в частности Жигулевских горах Самарской Луки, кроме основной биоморфы - высокого дерева, встречается многоствольно-стланцевая форма дуба. На других вершинах Жигулевских гор имеются аналогичные дубовые заросли. Впервые о таких необычных для дуба биоморфах упоминает А. М. Семенова-Тянь-Шанская, проводившая исследования растительного покрова на территории Жигулевского заповедника в 1945г. в составе экспедиции от Ботанического института им. Комарова. Среди широколиственных лесов Жигулей она выделила тип – остепненных дубняков (Семенова-Тян-Шанская, 1946). К этой категории она относит три типа ассоциаций дубняков, называя их при этом «дубовым криволесьем». Все три произрастают на южных склонах гор. Первый приурочен к верхней опушке леса, вторые окаймляют обычно сосновые леса снизу, третьи растут на маломощных щебнистых почвах южных пологих склонов древних оврагов (Семенова-Тян-Шанская, 1946). Очевидно, что здесь речь идет не об деревьях, а о стланцах или полустланцах.
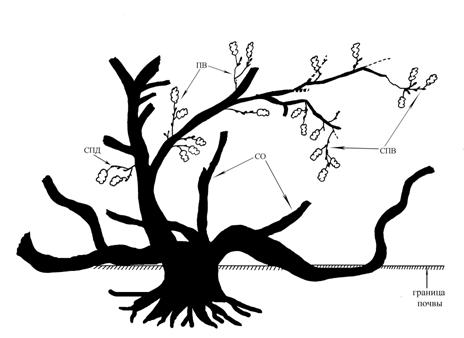
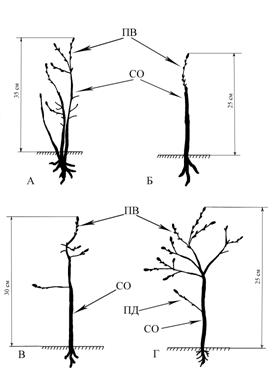
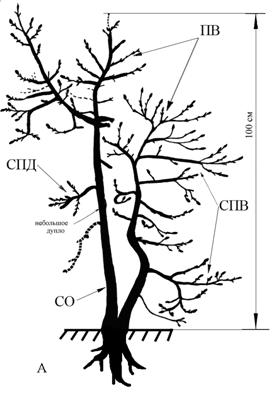
Рис 1. Полустланцевая биоморфа. Самарская область, Ставропольский район,
Жигулевский заповедник, вершина хребта Малой Бахиловой горы.
Слабая покатость к западу, граница степи и сосново-широколиственного
леса, на каменистом субстрате. Примечания: ПВ – побеги ветвления,
СО – скелетные оси, ПД – побеги дополнения, СПД – системы побегов дополнения,
СПВ – системы побегов ветвления. Отмершие части
растения выполнены на рисунке прерывающейся
линией. Тушью залиты живые частиС.В. Саксонов по результатам своих исследований в Жигулевском заповеднике описывает с участием «корявых» дубов сообщество – низкогорный корявый дубняк стоповидноосоковый. Эти сообщества находятся в верхних пологопокатых частях крутых склонов и узких гребнях восточной, южной и западной экспозиции и граничат с участками каменистых степей и остепненных сосняков. Сообщества описаны из трех точек на территории Жигулевского заповедника (Саксонов и др., 2006). Дуб здесь низкоствольный высотой 2,5-4 м,, нижние ветви стелются, простерты на земле. Кроме «корявых» дубов в этих сообществах встречаются реликтовые и эндемичные кальцефильные виды (Саксонов и др., 2006). Указывается, что сообщества такого рода есть лишь в Самарской области, на территории Жигулевского заповедника.
Биоморфы дуба черешчатого
Мы исследовали полустланцевую биоморфу дуба на склонах вершины хребта Малой Бахиловой горы. Склон к западу слабо покатый, растения растут на границе каменистой степи и сосново-широколиственного леса. Высота вершины над уровнем моря – 183 м. (рис. 1).
Возраст полустланцевой формы установить затруднительно. От основания ствола шириной 20-25 см отходят три ствола (скелетные оси) радиально расположенные по отношению к центру растения, стелющихся по склону. Стелющиеся стволы, отходя от основания, прижимаются к земле, какое-то время «ползут» по субстрату, затем выпрямляются, начинают нарастать ортотропно (по направлению «вверх»), как у дерева обычной биоморфы. Таким образом, сформированная крона оказывается гораздо ближе к поверхности земли, что возможно для растения в данных условиях более выгодно.
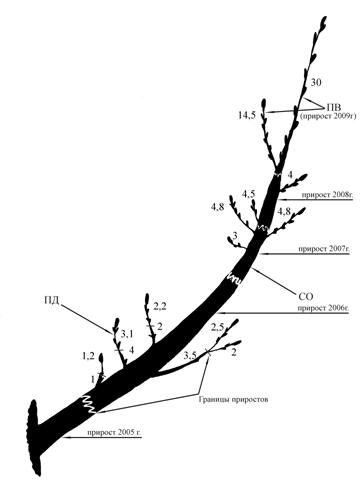
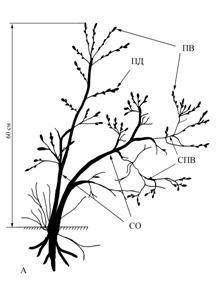
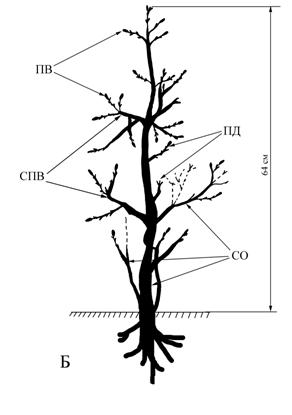
Рис. 2 Типичное ветвление дуба черешчатого
(прямоствольного) в условиях Самарской области
(Красноярский район, окр. с. Русская Селитьба).
Примечание: Цифрами указана величина прироста в смДва разновозрастных ствола нарастают ортотропно также от основания растения. Их диаметр в основании – 5-10 см. Высота более крупного ортотропного, ветвящегося ствола 2-2,5 м. Боковые ветви (СПВ) 20-25 см ветвятся до 3-4 порядка. Приросты 1-5 см, не типичные для основной формы роста дуба черешчатого. Вторичный, более молодой ствол слабо ветвится. Стоит обратить внимание на то, что отдельные побеги и небольшие системы побегов расположены на стволах вразброс, нерегулярно. Нет симметричного расположения боковых ветвей, свойственных типичной биоморфе дуба (рис.2). У полустланца изменяется не только биоморфа, но и тип ветвления. По нашему предположению главный экстремальный фактор – сильная обдуваемость ветром, и как следствие - сухость.
Такая форма роста не типична для дуба черешчатого и является явным отклонением от основной биоморфы. Вполне возможно, плагиотропное направление роста стволики приняли на второй стадии онтогенеза, когда под действием ветра молодые и гибкие стволики полегали, прижимались к почве, оказываясь в более благоприятных условиях. Именно они и создали энергетическую базу растения, позволившую в дальнейшем создать ортотропный ствол по типу основной жизненной формы дуба черешчатого, то есть стремиться к восстановлению типичной для дуба онтогенетической траектории.
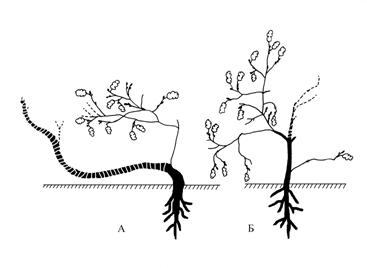
Процесс образования такого рода биоморфы возможно представить более полно при изучении молодых особей дуба, произрастающих вблизи описанного полустланца (рис. 1). На рисунке 3 представлены две особи, выросшие из семян, в возрасте 5-7 лет. Степень полегания у них различная. В случае 3А полегание более выражено, 3Б – менее. У растения 3А основной ствол, образовавшись, сразу начал полегать, хотя конец его отрастал ортотропно и поднимался над уровнем земли до 70 см. Затем по каким-либо причинам ствол усох до корневой шейки. Возникший побег дополнения сначала рос вертикально, затем приобрел плагиотропное направление роста. На момент исследования высота растения достигла около 40 см.
У растения (рис. 3Б) наблюдается лишь некоторое уклонение всей кроны в сторону, связанное с отмиранием части основного ствола. Приросты (ПВ) составляют 0,6-2 см. Общая высота растения – около 50 см.
Рис. 3. Молодые особи дуба черешчатого, склонные
к полустланниковости. Самарская область, Ставропольский район,
Жигулевский заповедник, вершина хребтаМалой Бахиловой горы.
Примечание: условные обозначения как на рис. 1Оба растения обнаружены не на самой границе леса и степи, а в разреженной части сосново-широколиственного леса на вершине Малой Бахиловой горы. Найденные молодые растения дуба, изображенные на рис. 3, встречаются единично, что соответствует слабому семенному возобновлению дуба. Стоит обратить внимание на то что имеются переходы от стланцев до стлаников.
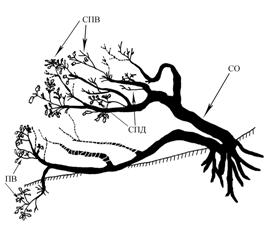
Сведения об аналогичных биоморфах дуба в Самарском Заволжье в литературе отсутствуют. Однако нами подобные формы наблюдались на севере Сергиевского района Самарской области, на склонах гор южной экспозиции у Кондурчинского водохранилища. Высота над уровнем моря – 206 м. Здесь на опушке лиственного леса обнаружены дубы стланцевой и полустланцевой формы (рис. 4).
У данных форм усиливается роль побегов дополнения, образующих на стволах небольшие системы СПД. Одновременно размеры побегов ветвления у этой биоморфы часто такие же как и у прямостоячей формы роста.
Рис 4. Стланиковая форма дуба. Самарская область,
Сергиевский район, окр. Кондурчинского водохранилища
в районе с. Крепость-Кондурча. Южная экспозиция
склона, граница лиственного леса и
разнотравно-кустарниковой степи. Примечание:
расшифровка сокращений и условные обозначения как на рис. 1В литературе имеются упоминания о стелющейся форме дуба на меловых холмах юго-востока Русской равнины (Мильков, 1951). На территории Казахстана, чуть ниже Оренбургской области. Стланцы там растут на меловых холмах левобережья Урала выше устья р. Утвы (Чингирлау). Высота растений 2 м. Прижимаясь к почве, стволы стелются на протяжении 6 м. Подобные экземпляры дуба находятся в составе низкорослых лесов, в составе которых береза, осина, вяз (Ulmus laevis), калина (Viburnum opulus), жимолость татарская (Lonicera tatarica), шиповник (Rosa cinnamomea).
Причинами, повлиявшими на формирование такой необычной биоморфы дуба, называются: неблагоприятные общеклиматические условия (юг степной зоны), иссушающие зимние и летние ветры, наиболее сильные на возвышенных частях рельефа, необычный для дуба меловой субстрат (Мильков, 1951).
Рис. 5. Одноствольные карликовые деревца.
Самарская область, Красноярский район,
окр. с. Русская Селитьба. Опушка дубравы.
Примечание: расшифровка сокращений
и условные обозначения как на рис. 1Вполне вероятно, что основной причиной, влияющей на образование стелющейся формы дуба и приближенной к ней (в случае Жигулевских гор) является более интенсивная обдуваемость ветрами. Возможно, это зависит от высоты и ориентации склона. В Казахстане же обдуваемость ветрами обеспечивается на относительно меньшей высоте (30-40 м над поймой) в связи с наличием безлесных пространств на большой площади.
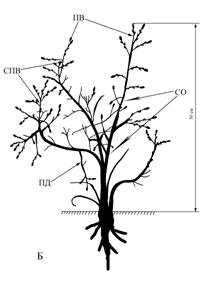
П. Л. Горчаковский, описывая южную границу распространения дуба, упоминает о низкоствольных дубах в районе Губерлинского мелкосопочника (река Губерля – правый приток Урала). Здесь в 30-летнем возрасте дуб достигает высоты 10 м. Ближе к опушке этих рощиц стволы дубов сильно изогнуты, многие ветви отмирают. Встречаются особи кустовидной или распластанной формы. Ближе же к центру колка деревья более прямоствольные и высокие (Горчаковский, 1968).Растения, имеющие отклонения от основной биоморфы, нами были обнаружены также за пределами дубравы. В Заволжье Самарской области в Красноярском районе, на юго-западных склонах Сокских яров у села Русская Селитьба есть дубки с биоморфой карликового одноствольного деревца (рис. 5).
Отклонение от основной траектории онтогенеза здесь заключается прежде всего в резком сокращении размеров всего растения и отдельных метамеров. У растения, изображенного на рисунке 5А, отсутствует типичный тип ветвления, свойственный дубу. Отрастают лишь единичные побеги ветвления (ПВ). Приросты сокращаются вдвое по сравнению с основной биоморфой. Усиливается роль побегов дополнения (ПД) возникающих из спящих почек. Причем они отрастают по всей протяженности ствола, т.е. из долго покоившихся почек. Эта биоморфа напоминает форму торчка вызванную сильным затенением в дубраве (Белостоков 1983). Известно, что в условиях стресса при сильном затенении в молодом возрасте у дуба черешчатого формируется форма торчка.
Одновременно от основания ствола, в зоне кущения, из спящих почек отрастают разновозрастные побеги формирования (ПФ). В начале роста усиленные, ростовые, а затем, также как и материнский образующие слабо ветвящуюся систему побега формирования (СПФ). Во втором и третьем вариантах (Б, В) дубки представлены только одним стволом высотой 25 и 30 см. Нарастание идет за счет единичных, очень слабых побегов ветвления.
В четвертом случае (вариант Г) картина иная. На верхушке невысокого 18 см стволика образуется небольшая крона из четырех, слабо и нерегулярно ветвящихся ПВ. Единичный побег дополнения отрос в середине стволика. В тех же местообитаниях встречается биоморфа дуба с более выраженной кустарниковостью (рис. 6). В этом случае насчитывается от 2-4 стволов различного размера и возраста.
Pис. 6а. Кустовидная биоморфа дуба черешчатого.
Самарская область,
Красноярский район, окр. с. Русская Селитьба.
Опушка дубравы Примечание: расшифровка сокращений
и условные обозначения как на рис. 1
Pис. 6б. Кустовидная биоморфа дуба черешчатого.
Самарская область, Красноярский район,
окр. с. Русская Селитьба. Опушка дубравы
Примечание: расшифровка сокращений
и условные обозначения как на рис. 1
Рис. 7. Кустовидная биоморфа дуба черешчатого
Самарская область, Сергиевский район,
окр. Кондурчинского водохранилища в районе
с. Крепость-Кондурча. Южная экспозиция склона,
граница лиственного леса и разнотравно-
кустарниковой степи. Примечание: расшифровка
сокращений и условные обозначения как на рис. 1Одни стволы лидируют, другие угнетены и отмирают. Побеги формирования имеют длину 7-14 см, отрастают от основания растения и служат резервом на случай гибели основного ствола. Они могут оставаться в угнетенном состоянии или отмирают. Побеги ветвления (ПВ) обращенные к свету с внешней адаксиальной стороны, более крупные, ортотропные. А те, что находятся с нижней стороны и более затенены, ветвятся долго и нерегулярно.
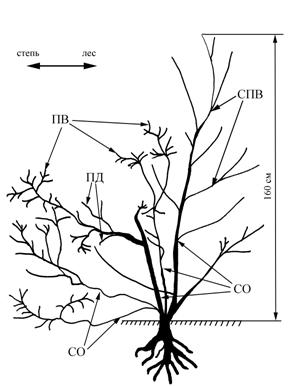
Кустовидные формы дуба есть и у Кондурчинского водохранилища. Здесь, в сочетании с кустовидностью, некоторые стволы приобретают стланцевую или стланиковую форму (рис. 7). Растение, изображенное на рисунке, находится на расстоянии 8 м от границы дубравы. Рост полегающих и наклонных стволов направлен в сторону степи. Два из них ползут по земле. Третий первоначально рос вверх, затем на высоте примерно 60 см сменил направление роста на плагиотропное. Четвертый ствол слабый, но растет исключительно вверх. Главный ствол высотой 160 см нарастает ортотропно, имеет приросты, характерные для прямоствольных дерев. Диаметры стволов данного растения – 1,5; 2; 4; 3 и 2 см.
Рис. 8а. Деревцевидно-кустовидная биоморфа дуба черешчатого.
Самарская область, Красноярский район,
окр. с. Русская Селитьба. Опушка дубравы.
Примечание: расшифровка сокращений
и условные обозначения как на рис. 1
Рис. 8б. Деревцевидно-кустовидная биоморфа дуба черешчатого.
Самарская область, Красноярский район,
окр. с. Русская Селитьба. Опушка дубравы.
Примечание: расшифровка сокращений
и условные обозначения как на рис. 1На южном и юго-западном склоне у Кондурчинского водохранилища, растут дубы склонные к кустовидности. На опушке лиственного леса растет дуб с одним стволом толщиной 23 см. Нижняя ветка длиной 2,5-3 м и толщиной 7 см, растет в сторону кустарниковой степи, наклонена. И находится почти на уровне земли.
У села Русская Селитьба Красноярского района на том же склоне юго-западной экспозиции встречаются также деревцевидно-кустовидные растения, высотой от половины до полутора метров, насчитывающие один или два неравных, но более сильных ствола (рис. 8).
В первом случае (рис. 8а) имеется два стволика толщиной в основании 1,7 и 1,2 см. Более крупный имеет форму дерева в своей верхней части с выраженной кроной. Второй стволик, высотой 75см, достигает лишь кроны первого, более высокого. В отличие от него, с внешней его стороны от основания до верхушки ветвятся в стороны, разновозрастные и разнообразные по своей структуре боковые ответвления – маленькие веточки заполняющие все воздушное пространство вокруг растения – от основания до его верхней части, и создающие эффект однобокого компактного куста.
Второй пример (рис. 8б) - одноствольное деревце с короткими боковыми ветвями, расположенными по всей протяженности стволика. Только верхний прирост сохраняет тип ветвления дуба черешчатого. Боковые ветви составлены системами мелких побегов, ветвящиеся нерегулярно. В основании ствола находятся три слабых, угнетенных побега формирования (ПФ). Главный ствол лидирует, четко проявляя тенденцию возврата к деревцевидной формы роста.
Заключение
Приведенные примеры демонстрируют разнообразие форм роста – от невысокого, карликового дерева с одним или двумя стволами, а также кустовидные формы с несколькими стволами, но меньших размеров, а также к склонности формировать стелющуюся форму роста. Ясно видны направления отклонений в сторону карликовости, кустовидности и стланиковости. И в то же время так же четко проявляются тенденции к возврату к основной форме дерева, к типичной для дуба черешчатого онтогенетической траектории.
Все описываемые растения расположены на опушке леса, на склоне холма высотой 180-200 м над ур. м. от слабой покатости до достаточно крутого уклона (более 90?). Условия произрастания отдельных экземпляров могут несколько отличаться. Расстояние между ними может быть различно, а граница леса расположена в пределах до 10 м. Отдельные экземпляры находятся прямо на опушке леса.
В составе лесного массива дубы представлены обычными деревьями. Очевидно, что формирование столь необычных форм дуба черешчатого связано с неблагоприятными условиями произрастания, в первую очередь, с сильными ветрами. Признаки кустовидности и стелющегося роста начинают проявляться в раннем возрасте. Но, в дальнейшем, когда растения могут обеспечить себя достаточной энергетической базой - пытаются возвратиться к форме дерева.
Отклонения, вызванные не типичными для вида условиями обитания, идут в сторону уменьшения метамеров, их нерегулярного отрастания, а также усиления возникновения побегов дополнения, компенсирующее их потери. Как следствие высота растений сокращается, они приобретают форму карликов. Уход от стресса выражается и в изменении направления роста - полегании стволов, образовании стелющихся форм роста и др.
Отклонения, связанные со стрессом, идут однонаправлено и, так же как и у других древесных растений, в условиях стресса (Мазуренко, 1986).
Кустарниковый дуб кермесовый на сухом каменистом склоне.
Фото с сайта: http://www.hedl.net/cestovani/…Все описываемые разнообразные отклонения, безусловно, являются факультативными, не закрепленными наследственно, что подтверждают быстрые реакции воызврата к основной форме роста.
Любое изменение в сторону более благоприятных условий вызывает тенденцию к возврату к основной биоморфе, к типичной для дуба черешчатого онтогенетической траектории. В данном случае стремление к формированию одного ствола, восстановлению типичного ветвления, увеличению размеров растений.
Среди представителей рода Quercus имеются кустовидные и стелющиеся биоморфы не факультативные, а наследственно закрепленные. Дуб кермесовый (Quercus coccifera L.) - типичный кустарник. В Средиземноморье он один из главных представителей формации маквис. Это его основная форма роста. Вместе с тем изредка встречаются довольно крупные деревья с плотной шаровидной кроной.
В горной Аджарии, на склонах Малого Кавказского хребта (Аджаро-Имеретинский, Шавшетский хребты и др.) растет реликтовый дуб понтийский (Qercus pontica K. Koch) - крупный стланец. Один из представителей колхидского подлеска «шкериани». Под лесной полог эта светолюбивая порода не заходит, а приурочена к субальпийскому поясу на высоте 2500-2700м над ур.м. Полегающие стволы от 6 до 12 м в длину, простерты вниз по склону. Краевые ветви саблевидно приподнимаются на высоту до 2 иногда и более м. Навалы снега прижимают стволы к земле. Они укореняются, постепенно нарастая и продвигаясь вниз по склону. С противоположного так же последовательно отмирая. (Долуханов, 1980, Гвианидзе, Гогитидзе, 1990). В условиях культуры в Батумском ботаническом саду на высоте 60 м над ур.м. в субтропическом климате, где снежные навалы возникают эпизодически и только на несколько дней – дуб понтийский растет как высокий кустарник с несколькими ортотропными скелетными осями. Такая же форма роста для дуба понтийского сохраняется и в других географических условиях – севернее субтропического пояса, при посадках в дендропарках Европы.
Дуб кермесовый: листья, плоды.
Фото с сайта: http://restauracionpaisajistica.comПример с дубом понтийским весьма показателен. Форма стланца – его основная. В процессе онтогенеза дуба понтийского в природных условиях - стланец формируется со второго периода онтогенеза (Гвианидзе, Гогитидзе, 1990) и существует в виде вегетативно-подвижного растения, задерживаясь на стадии взрослого растения, благодаря укоренению скелетных осей. Не проявляя признаков старения.
В условиях культуры дуб понтийский не становится вегетативно-подвижным, изменяя свою онтогенетическую траекторию. Главный фактор стресса, формирующий стланик – навалы снега. В условиях культуры дуб понтийский отклоняется от своей онтогенетической траектории. Вполне вероятно возникает биоморфа свойственная для этого вида до третичных похолоданий. В современное время она встречается только в культуре, и ее возникновение носит факультативный характер. А основная биоморфа, наследственно закрепленная в современное время, тысячелетиями подвергается влиянию навалов снега, укрывающих его стволы и ветви. Этот фактор способствует сохранению основной биоморфы вида в подобных условиях.
Приведенные примеры показывают тенденции формирования биоморф при влиянии экстремального фактора и возможность наследственного закрепления биоморфы при отклонении от основной (без определенного фактора стресса) онтогенетической траектории. Безусловно, влияние стресса должно быть весьма длительным, насчитывая несколько тысячелетий и более, как это демонстрирует пример с дубом понтийским – реликтом третичного периода, сохранившимся до наших дней в весьма специфических условиях.
Таким образом, в новых условиях, при отклонении от своей основной онтогенетической траектории, при наследственном закреплении вид приобретает в этих условиях свою, отличную от основной траекторию, которая закрепляется за ним как основная. Возможно это и является важным моментом при видообразовании. Этот тезис подтверждают исследования жизненных форм родов рододендрон (Rhododendron), багульник (Ledum), кассиопея (Cassiope) на севере Дальнего востока (Мазуренко 1980,1982,1984, Мазуренко, Хохряков, 1981,1987). Примеров множество.
Дуб понтийский. Фото с сайта:
http://www.monadire.com/2011/0…Дуб обыкновенный под влиянием стресса за пределами дубрав в Самарской области показывает те же направления адаптивной эволюции жизненных форм растений. Но, будучи лишь отклонением и не приобретя своей траектории, стремится к возврату на основную, наследственно закрепленную онтогенетическую траекторию. Процесс разбалансировки – процесс длительного влияния фактора стресса.
Литература
Белостоков Г.П. Системы стеблесложения подроста древесных растений // Бюл. МОИП, отд. биол., 1983, т.88, вып.1. - С. 87-104.
Белоусов Л.В Межуровневые отношения в онтогенезе // Журн. общ. биол. Т. 54 1983. - С. 23-31.
Благовещенский В.В. Растительность Приволжской возвышенности в связи с ее историей и рациональным использованием. Ульяновск, 2005. - 715 с.
Вахромеева М.Г. Морфологическая характеристика возрастных состояний клена остролистного // Вестн. МГУ. 1975. №6. -С. 116-119.
Гвианидзе Д.В., Гогитидзе Н.В. Биология, морфогенез и ритм развития дуба понтийского в природных условиях и в культуре // Известия Батумского ботанического сада. Батуми, 1990. т. 29. - С. 45-55.
Горчаковский П.Л. Растения европейских широколиственных лесов на восточном пределе ареала // Труды Института экологии растений и животных. Академия наук СССР, Уральский филиал. Свердловск, Вып. 59. 1968. - 207 с.
Долуханов А.Г. Колхидский подлесок. Тбилиси, Мецниереба, 1980. - 260 с.
Жукова Л. А. Популяционная жизнь растений. Йошкар-Ола, 1995. - 224 с.
Жукова Л.А. Многообразие путей онтогенеза в популяциях растений // Экология. 2001. № 3. - С. 169-176.
Жукова Л.А. Онтогенез и циклы воспроизведения растений // Журн. общ. биол. 1983. Т. 44. № 3. - С. 361-374.
Жукова Л.А. Онтогенез и циклы воспроизведения растений. Йошкар-Ола, 1995. - 224 с.
Заугольнова Л.Б. Возрастные этапы в онтогенезе ясеня обыкновенного (Fraxinusexcelsior L.) // Вопросы морфогенеза цветковых растений и строение их популяций. М.: Наука, 1968. - С. 81-102.
Мазуренко М.Т Биоморфологические адаптации растений к экстремальным условиям крайнего севера М.: Наука, 1986. - 209 с.
Мазуренко М.Т. Вересковые кустарнички Дальнего Востока. М.: Наука., 1982. -184 с.
Мазуренко М.Т. О гомеостазе онтогенеза древесных растений // V Всесоюзная школа теоретической морфологии растений. Львов, 1987. С. 65-69.
Мазуренко М.Т. О жизненной форме стелющихся лесных растений // Бот. журн. 1978. т. 63. № 4. - С. 593-603.
Мазуренко М.Т. Основные направления адаптации рода рододендрон к экстремальным условиям // Maт. V Моск. совещ. по филогении растений. М., 1976. - С. 104-107.
Мазуренко М.Т. Рододендроны Дальнего Востока. Структура и морфогенез М.: Наука., 1980. - 231 с.
Мазуренко М.Т. Структура побеговых систем, онтогенез и эволюция дальневосточных кассиопей // Журн. общ. биол. 1984. № 3. - С. 336-347.
Мазуренко М.Т. Хохряков А.П. Структура и морфогенез кустарников. М.: Наука, 1977. - 160 с.
Мазуренко М.Т., Хохряков А.П. Биоморфологическая изменчивость и ее связь с таксонообразованием урастений // Жизненные формы: структура, спектры и эволюция. М., 1981. - С. 12-30.
Мазуренко М.Т., Хохряков А.П. Биоморфологическая эволюция дальневосточных багульников // Журн. общ. биол. 1987. Т. ХLVIII. № 2. - С. 190-201.
Матвеев В.И., Матвеева Т.Б, Соловьева В.В. Quercus robur L. как вид, рекомендуемый для внесения в Красную книгу Самарской области // Раритеты флоры Волжского бассейна: доклады участников российской конференции. Тольятти, 2009. – С. 125-138.
Махатков И.Д. Поливариантность онтогенеза пихты сибирской // Бюлл. МОИП, отд.биол. 1991. Т. 96. вып. 6. - С. 79-88.
Мильков Ф.Н. Стелющаяся форма дуба на меловых холмах юго-востока Русской равнины // Природа. № 6, 1951. - С. 49-50.
Полтинкина И.В. Онтогенез, численность и возрастной состав ценопопуляции клена полевого в широколиственных лесах Европейской части СССР // Бюлл. МОИП, отд. биол. 1985. Т. 90. Вып. 2. - С. 79-88.
Рысин Л.П., Рысина Г.П. Дуб обыкновенный // Биологическая флора Московской области. М., 1990. ч. 8. - С. 102-130.
Савиных Н.П. Поливариантность особей и ее эволюционное значение // Методы популяционной биологии: Сб. материалов докл. VII Всерос. популяционного семинара. Сыктывкар, 2004. Ч. 1. - С. 186-187.
Саксонов С.В., Лысенко Т.М., Ильина В.Н., Конева Н.В., Лобанова А.В., Матвеев В.И., Митрошенкова А.Е., Симонова Н.И., Соловьева В.В., Ужамецкая Е.А., Юрицына Н.А. Зеленая книга Самарской области: редкие и охраняемые растительные сообщества / Под ред. Г.С. Розенберга и С.В. Саксонова. Самара, 2006. - 201 с.
Семенова-Тян-Шанская А.М. Растительный покров Куйбышевского государственного заповедника. Ботанический институт АН СССР им. Комарова. 1946. - 54 с.
Серебряков И.Г. Экологическая морфология растений. М., 1962. - 378 с.
Смирнова О.В., Заугольнова Л.Б., Торопова Н.А., Фаликов Л.Д. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений разных биоморф // Ценопопуляции растений. Основные понятия и структура. М., 1976. - 215 с.
Смирнова О.В., Чистякова А.А. Анализ поведения некоторых древесных растений широколиственных лесов европейской части СССР // Биология, экология и взаимоотношения ценопопуляций растений. М., 1982. - С. 52-56.
Смирнова О.В., Чистякова А.А. Анализ фитоценотических потенций некоторых древесных видов широколиственных лесов европейской части СССР // Журн. общ. биол. 1980. Т. 41. № 3. - С. 350-362.
Уранов А.А. Онтогенез и возрастной состав популяций // Онтогенез и возрастной состав популяций цветковых растений. М., 1967. - С. 3-8.





|
|